











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
En el océano Pacífico Tropical Oriental (PTO) existe una franja con diferentes niveles de riqueza de especies de colonias de corales formadores de arrecife en México, El Salvador, Nicaragua, Costa Rica, Panamá, Colombia, Ecuador, así como las islas del Coco, Galápagos, Malpelo, Revillagigedo y otras (Cortés, 2003). Diferentes procesos geológicos, geográficos y oceanográficos permitieron la existencia de especies de corales hermatípicos en esta región (Laborda, 2018).
En El Salvador se documentó la presencia del coral duro Porites lobata Dana, 1843 en la franja intermareal baja, hasta los 2.5 m de profundidad aproximadamente, en el área natural protegida (ANP) y sitio Ramsar ''Complejo Los Cóbanos” (13º 31'22.0” N, -89º 48'27.2” W) (Barraza, 2017; Molina, 2004; Segovia-Prado & Navarrete-Calero, 2007). También, mediante observaciones de restos de coral en playa, se confirmó la ocurrencia de especies hermatípicas desaparecidas de esta ANP, como Pocillopora capitata Verrill, 1864, P. damicornis (Linnaeus, 1758) y P. meandrina Dana, 1846 (Reyes-Bonilla & Barraza, 2003). Otras especies de coral duro observados en el área son Psammocora obtusangula (Esper, 1749) y P. stellata (Verrill, 1866) (López & Jiménez, 2008). Esta área presenta una importante riqueza de especies de invertebrados (Barraza, 2014) y peces (González-Murcia et al., 2016), así como de cetáceos (Ibarra-Portillo et al., 2022). Existen evidencias físicas, aunque no documentadas en literatura, como la existencia una línea de rieles para carritos de acarreo de material en la zona, que refleja que al menos una especie de corales de las mencionadas anteriormente fue explotada para fabricar cemento en el área portuaria de Acajutla entre 1951 a 1953, aproximadamente.
Recientemente, varias especies de coral a nivel global experimentaron importantes daños por blanqueamiento durante períodos extremos de El Niño, incluyendo el bienio 2015-2016 debido a la elevación de la temperatura del agua de mar y factores ambientales adversos asociados a actividades humanas (Alvarado et al., 2020). Blanqueamiento es la ruptura de la simbiosis entre las microalgas simbiontes que habitan en los tejidos de corales hermatípicos provocando que éstos presenten una coloración blanquecina, se vuelven más vulnerables a enfermedades y muerte (Boilard et al., 2020; Douglas, 2003). Eakin et al. (2019) ampliaron las causas de este evento de blanqueamiento de coral debido al estrés por calor entre 2014 a 2017. En la región centroamericana también han ocurrido este tipo de episodios, como blanqueamientos por pérdida de zooxantelas y muerte de corales escleractinios hermatípicos que sucedió en las costas Pacífica y Caribeña de Costa Rica, ocurrido entre 1982 y 1983 (Cortés et al., 1984). Similar situación se observó en el Pacífico de Panamá en 1983 (Glynn, 1983). En el golfo de California, México, se registró blanqueamiento importante de corales duros en 1987 (Reyes-Bonilla, 1993).
En El Salvador se ha documentado poca información sobre eventos de blanqueamiento, por ejemplo, Barraza (2020) mencionó que estos episodios experimentados por colonias intermareales y submareales de P. lobata en El Salvador posiblemente estuvieron asociados a temperaturas elevadas del agua, así como floraciones algales causadas por Margalefidinium polykrikoides (Margalef) F. Gómez, Richlen & D.M. Anderson, 2017 (anteriormente Cochlodinium polykrikoides).
Ante los eventos de pérdida de zooxantelas y muerte de corales a nivel global, comenzaron a aparecer iniciativas de restauración directa, es decir ubicar o reubicar fragmentos o colonias de estos cnidarios en zonas donde las poblaciones se redujeron. En el Pacífico de Costa Rica comenzaron las técnicas de trasplante exitoso de fragmentos de Pocillopora spp. en isla del Caño (Guzmán, 1991). Posteriormente se ha considerado que componentes indirectos como la participación ciudadana, entrenamiento y manejo de áreas naturales protegidas son elementos clave para la complementar iniciativas exitosas de trasplante de estos organismos (Alvarado et al., 2017).
Debido a las diversas amenazas como desarrollo de la infraestructura turística, extracción de vida silvestre no autorizada, debilidad institucional en la zona, entre otras, se han realizado diversos esfuerzos por restaurar la biodiversidad local (Alvarado et al., 2017). En respuesta se colocaron arrecifes artificiales con el objeto de promover la colonización de peces y otra biota, incluyendo corales; también se declaró área natural protegida mediante Decreto Ministerial No. 22 publicado en el Diario Oficial del 12 de febrero de 2008, se creó la unidad de guarda recursos en esa área, entre otras actividades. Además, se declaró sitio Ramsar el 02 de febrero de 2019 (Mariona, 2020). A pesar de estos esfuerzos nacionales mencionados, eventos globales como el blanqueamiento afectaron las poblaciones locales de P. lobata (Barraza, 2020). Hasta el momento las actividades de restauración de la biodiversidad acuática en esta área natural protegida han enfatizado en esfuerzos que Edwards & Gómez (2007) clasificó como indirectos y en menor grado directos.
Este documento presenta un análisis de más de 200 fotografías que reflejan el estado de los corales incluyendo los principales eventos de blanqueamiento de P. lobata en el ANP Los Cóbanos entre 2006 a 2022, además se detalla una recopilación cronológica de los esfuerzos realizados para la conservación de los arrecifes y corales hermatípicos en El Salvador. Además, presenta el uso de una herramienta importante para medir el estado de la simbiosis entre las zooxantelas y los corales huéspedes es la fluorescencia de clorofila mediante la emisión de pulso de amplitud modulada (PAM en inglés), debido a que refleja cambios de la eficiencia fotosintética de las algas, indicando en forma indirecta el estado de salud del coral (Warner et al., 2010).
Materiales y métodos
Área de estudio: La recopilación de datos sobre blanqueamiento se basa en aproximadamente 2 100 registros fotográficos subacuáticos y de campo del autor, realizados mediante buceo autónomo, así como snorkel en una zona intermareal y una submareal, a partir de los años 2006-2007 hasta agosto de 2022 sin una frecuencia establecida (Tabla 1, Fig. 1), en el área natural protegida y sitio Ramsar Complejo Los Cóbanos, departamento de Sonsonate, El Salvador.
Tabla 1
Estado del blanqueamiento de las colonias de P. lobata en el área El Faro, área natural protegida Complejo Los Cóbanos (2007-2022). / Table 1. Bleaching condition of P. lobata colonies in El Faro area, natural protected area Complejo Los Cóbanos (2007-2022). Categorías: sin evidencia (SE), tenue (TE), parches (PA), severo (SV). Categories: without evidence (SE), light (TE), patches (PA), severe (SV).
AÑO
SE
TE
PA
SV
AÑO
SE
TE
PA
SV
2007
EN 07
100
2017
EN 17
60
40
MA 07
100
MA 17
85
15
AG 07
60
40
NO 17
100
SE 07
75
25
2018
EN 18
100
2008
EN 08
90
10
MA 18
40
FE 08
100
JU 18
50
50
2009
MA 09
100
SE 18
10
90
OC 09
40
60
OC 18
78
22
2010
EN 10
100
DI 18
95
OC 10
100
2019
EN 19
95
2011
FE 11
80
20
MA 19
70
28
OC 11
80
20
JL 19
35
65
2012
FE 12
85
15
AG 19
66
32
MA 12
60
40
SE 19
46
52
AB 12
50
50
OC 19
41
59
MY 12
20
80
NO 19
69
31
NO 12
50
30
20
2020
FE 20
75
25
2013
EN 13
80
20
AG 20
23
76
FE 13
100
OC 20
95
MA 13
85
15
NO 20
100
OC 13
80
20
DI 20
100
DI 13
70
30
2021
FE 21
40
60
2014
FE 14
90
10
MA 21
100
SE 14
100
AB 21
20
80
OC 14
100
MY 21
97
DI 14
50
40
10
JU 21
95
2015
EN 15
100
JL 21
95
FE 15
100
OC 21
10
87
MA 15
50
50
NO 21
78
10
AB 15
100
DI 21
50
50
SE 15
100
2022
EN 22
70
30
OC 15
100
MA 22
60
40
DI 15
95
AB 22
20
80
2016
EN 16
80
20
MY 22
91.4
8.6
FE 16
85
15
MA16
90
10
AB 16
90
10
JU 16
20
80
OC 16
30
70
DI 16
100
Categorías: sin evidencia (SE), tenue (TE), parches (PA), severo (SV). Categories: without evidence (SE), light (TE), patches (PA), severe (SV).

Fig. 1 Categorías de blanqueamiento de coral (P. lobata) utilizadas en este estudio: A. sin evidencia de blanqueamiento, las marcas blancas corresponden a mordeduras de peces, B. blanqueamiento leve, cuando se conservan parte de la coloración natural, C. parche, pérdida de tejido a causa del blanqueamiento, se puede observar espacios colonizados por algas, D. blanqueamiento severo, pérdida total de las zooxantelas. / Fig. 1. Bleaching categories of coral (P. lobata) used in this study: A. without bleaching evidence, white marks belong to fish bites. B. light bleaching, when part of natural coloration is retained, C. patch, loss of tissue due to bleaching, areas colonized by algae can be observed, D. severe bleaching, total loss of zooxanthellae.
Método de muestreo: El primer tipo de reconocimiento se realizó en el área somera denominada ''El Faro” cercano a Punta Remedios, para ello se realizó un recorrido de 287 m entre un punto de origen zona litoral de playa (Fig. 2B) (13º 31'16.3” N - 89º 48'24.4” W), y otra zona ubicada al suroeste en el área submareal (Fig. 2A) (13º 31'25.6” N - 89º 48'22.5” W), en este transecto se tomaron fotografías de colonias de coral y observaciones en un área visual de 2 metros a cada lado en época seca (noviembre a abril) y lluviosa (mayo-octubre). Las fechas de los muestreos se presentan en la Tabla 1.
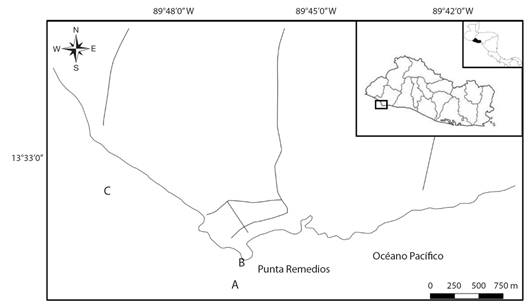
Fig. 2 Mapa de Centroamérica, El Salvador y zona de estudio dentro del sitio Ramsar Complejo Los Cóbanos. A. área de ubicación de colonias de P. lobata someras, B. zona intermareal de referencia para el transecto de monitoreo A-B, C. colonias submareales de P. lobata. / Fig. 2. Map of Central America, El Salvador and study zone within Ramsar site Complejo Los Cóbanos. A. Location area of shallow P. lobata colonies, B. reference intertidal zone for monitoring transect A-B, C. subtidal colonies of P. lobata.
La segunda forma de monitoreo incluyó observaciones totalmente al azar en un transecto desde la embarcación hacia el noroeste, en un área de colonias submareales de P. lobata (0.1 km2, 2 - 3 m de profundidad, en bajamar (Fig. 2C) (13º 31'47.7” N - 89º 49'8.1” W) mediante buceo autónomo por un período de 30 a 45 minutos durante la época seca. La ubicación de las áreas de monitoreo se detalla en la Fig. 2. Las fechas de estos registros se incluyen en la Fig. 3.
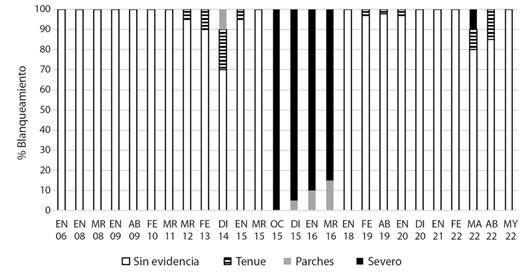
Fig. 3 Porcentajes de los diferentes tipos de estados de blanqueamiento en las colonias submareales de P. lobata, entre 2006 y 2022. / Fig. 3. Percentages of the different bleaching condition types in P. lobata subtidal colonies between 2006 and 2022.
Además, entre los años 2006 y 2007 en época seca, se realizaron al menos cuatro diferentes observaciones mediante buceo autónomo a 6-7 m de profundidad, sin frecuencia definida, a unos arrecifes tipo domo (''reefball”) en el ANP Complejo Los Cóbanos para determinar la posible colonización y duración de este tipo de estructura. Los registros se basaron en la presencia de taxones. El monitoreo duró menos de seis meses debido a los impedimentos de visibilidad causados por la época lluviosa, en el siguiente período seco, las estructuras ya no se encontraron. Estos arrecifes artificiales fueron colocados por una organización no gubernamental denominada FUNDARRECIFE (Fundación para la Protección del Arrecife de Los Cóbanos).
Durante este período se fueron incorporando registros puntuales de temperatura del agua superficial con equipo portátil SUUNTO, HACH y posteriormente YSI (Professional Plus) a partir de 2011 y con sensor HOBO ONSET (usualmente 6 registros por día) a, 16 m de profundidad, entre 2019 a 2021. Estas mediciones continúan realizándose hasta el momento con equipo portátil. La eficiencia fotosintética de colonias de coral someras y submareales se midió utilizando un fluorómetro subacuático de pulso de amplitud modulada (DIVE-PAM, WALZ) entre septiembre de 2014 a mayo de 2016 (Fig. 4). Para ello, se colocó un cilindro corto estándar al extremo libre, que permitía mantener una distancia fija de 1 cm a 90° de la superficie del coral a aplicarle el pulso de luz. Los parámetros de fluorescencia incluidos en este estudio abarcan el rendimiento efectivo cuántico del fotosistema II de la fotosíntesis (Φ) y el máximo rendimiento cuántico del mismo fotosistema definido por la relación de la fluorescencia variable y la fluorescencia máxima (FV/FMAX), en ambos casos en condiciones de luz solar sin adaptación a oscuridad (diurno). El total de registros para este parámetro fue 358 lecturas (Fig. 4, Fig. 5). Para determinar el grado de blanqueamiento de P. lobata se establecieron cuatro patrones principales que se detallan en la Fig. 1. La cobertura superficial de las diferentes categorías de blanqueamiento propuestas se calculó al observar cada imagen de colonia de coral estableciéndose porcentajes para cada muestreo fotográfico en el que se utilizaron cámaras subacuáticas (Canon G12, Olympus C-760 y TG-6). La cantidad de fotografías durante cada muestreo osciló entre 15 a 50 en la franja de El Faro y 20 a 40 en la zona submareal, según las condiciones de visibilidad en el agua, altura y corrientes de marea.
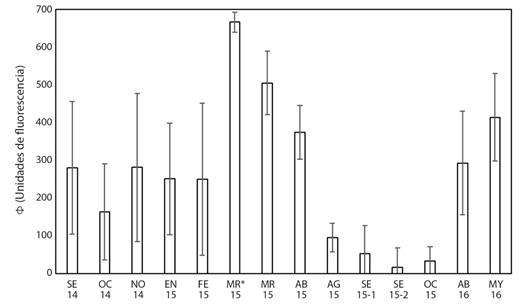
Fig. 4 Promedios del rendimiento efectivo cuántico del fotosistema II (Φ) diurno de las colonias de P. lobata en el área El Faro y submareales (*) entre septiembre de 2014 a mayo de 2016. Líneas verticales representan la desviación estándar. Se presentan dos registros de datos de septiembre 2015. / Fig. 4. Averages of effective diurnal quantum yield of photosystem II (Φ) of P. lobata colonies in El Faro and subtidal (*) areas between September 2014 to May 2016. Vertical lines represent standard deviation. Two sets of data of September 2015 are presented.
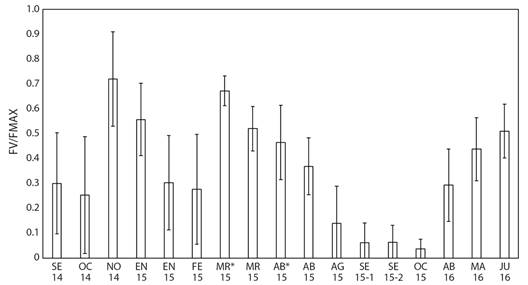
Fig. 5 Promedio del máximo rendimiento cuántico del fotosistema II (FV/FMAX) de las colonias de P. lobata en el área El Faro y submareales (*) entre septiembre de 2014 a mayo de 2016. Líneas verticales representan la desviación estándar. / Fig. 5. Average of maximum quantum yield of photosystem II (FV/FMAX) of P. lobata colonies in El Faro and subtidal (*) areas between September 2014 to May 2016. Vertical lines represent standard deviation.
Análisis de datos: se calculó el promedio y desviación estándar de los datos de temperatura, así como del rendimiento efectivo cuántico del fotosistema II (Φ) diurno, también del máximo rendimiento cuántico del fotosistema II diurno (FV/FMAX) de las colonias someras y submareales (Hoogenboom et al., 2012; Magnusson, 1997; Wangpraseurt et al., 2019).
Resultados
Blanqueamiento: El estado de las colonias someras de coral (El Faro), según la categorización incluida en este estudio, se revela la ausencia de la pérdida de pigmento y también blanqueamiento tenue entre 2007 hasta el primer semestre de 2015. Posteriormente, el blanqueamiento severo se volvió recurrente, lo que condujo a una pérdida del 40-50 % de la cobertura original, con base a estimaciones del autor basadas en la categorización propuesta aplicada a los registros fotográficos (Tabla 1). Los blanqueamientos severos de cobertura importante sucedieron entre septiembre y diciembre de 2015, junio de 2016, septiembre de 2018, julio-septiembre de 2019 y agosto de 2020. Además, eventos de floraciones algales nocivas, particularmente las causadas por el dinoflagelado Margalefidinium polykrikoides afectaron a las colonias del coral duro de esta zona en varias ocasiones, incluyendo mayo de 2012 (Espinoza et al., 2012) y marzo de 2022 (Genovés, 2022).
Las colonias submareales de mayor tamaño (2-4 m de diámetro máximo) presentaron un episodio de blanqueamiento severo en más del 80 % de la cobertura de P. lobata entre octubre 2015 y marzo 2016. Pero se recuperaron y la mortalidad fue reducida después de este episodio (5 %, 2 colonias muertas/40, enero de 2018) (Fig. 3). Otro blanqueamiento severo y tenue (10 % cada uno) ocurrió en marzo de 2022. Esta especie de coral masivo reflejó una recuperación importante en abril de 2022.
La temperatura superficial del agua en El Faro varió entre 25.0 a 36.0 oC (prom = 30.3 ± 2.1 °C) (Fig. 6A). Los picos más altos de este parámetro ocurrieron en septiembre de 2015 y 2016, coincidiendo con el blanqueamiento severo del primer año. En la zona de las colonias submareales la temperatura osciló entre 27.0-31.7 oC (prom = 28.3 ± 0.9 °C), que corresponden a datos registrados solamente en época seca (octubre-mayo) (Fig. 6B). Además, a partir de noviembre de 2019 hasta septiembre de 2021 se registraron 4 033 datos por el sensor de profundidad (16 m), reflejando valores oscilantes entre 20.7-31.3 oC (prom = 28.6 ± 1.5 °C), entre noviembre de 2019 y agosto de 2020, respectivamente.
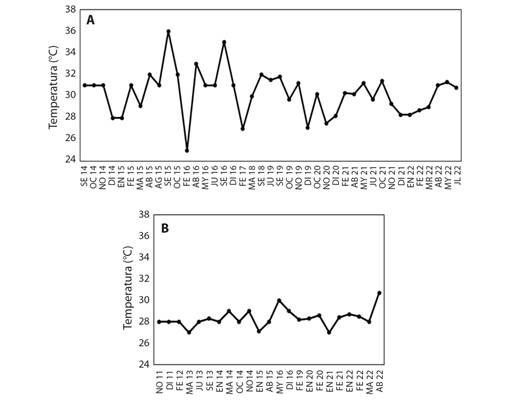
Fig. 6 Temperaturas del agua superficial en zonas de monitoreo de blanqueamiento de P. lobata en el área natural protegida Complejo Los Cóbanos: A. El Faro, B. colonias submareales. / Fig. 6. Surface water temperature in bleaching monitoring zones at natural protected area Los Cóbanos: A. El Faro, B. subtidal colonies.
El promedio del rendimiento efectivo cuántico diurno (Φ) alcanzó picos elevados entre marzo y abril de 2015 (504.4 ± 89.2, 373.8 ± 70.9 unidades de fluorescencia (uf), respectivamente para las colonias de El Faro, y 666.1 ± 26.8 uf en las colonias submareales). Posteriormente presentó importantes descensos entre agosto y octubre de 2015 (16.5 ± 50.7 a 94.7 ± 37.6 uf en colonias de El Faro), coincidiendo parcialmente con las fechas de los blanqueamientos observados en las colonias someras y submareales (Fig. 3, Fig. 4, Tabla 1). El promedio del índice diurno FV/FMAX varió de forma similar: niveles altos en las colonias intermareales entre marzo y abril de 2015 (0.52 ± 0.09 y 0.37 ± 0.11, respectivamente), las submareales presentaron índices más altos en los mismos meses: 0.67 ± 0.06 y 0.46 ± 0.15, respectivamente, también hubo descensos importantes en las colonias intermareales entre agosto y octubre 2015: 0.04 ± 0.04 a 0.14 ± 0.15 (Fig. 5).
Restauración indirecta y directa: La revisión de literatura demuestra algunos esfuerzos nacionales indirectos con incidencia en la restauración de las colonias de coral en el país como: a) aprobación de la Ley del Medio Ambiente cuyo artículo 74 declara inalterables a los manglares y arrecifes, en esa misma normativa se requiere a los nuevos proyectos elaborar un estudio de impacto ambiental para analizar, aprobar o rechazar su ejecución (Ley de Medio Ambiente, 1998); b) a través de la aprobación de la Ley de Áreas Naturales Protegidas en 2005, se declara bien nacional a los arrecifes rocosos y coralinos, lo que permitió ubicar a Los Cóbanos en ese estatus en 2008; c) desarrollo de proyectos de investigación básica en el área y organización social con financiamiento nacional e internacional desde hace más de 15 años; d) inclusión de corales hermatípicos y ahermatípicos en el listado oficial de especies de vida silvestre amenazadas y en peligro de extinción a partir de 2009 (Acuerdo Ministerial No 36); e) establecimiento del sitio Ramsar Complejo Los Cóbanos en febrero de 2019 (Mariona, 2020).
Las actividades de restauración directas incluyeron la colocación de 50 arrecifes tipo reefball (domo) de aproximadamente 1.30 m de diámetro y 0.80 m de altura que se ubicaron a profundidades entre 5-7 m sobre fondo arenoso en el ANP Los Cóbanos (Fig. 7) (2006-2007) con diversas fuentes de financiamiento. Estas estructuras se ubicaron para permitir la recolonización de organismos marinos abarcando corales (Instituto de Ciencias del Mar y Limnología de El Salvador (ICMARES), 2008). Los grupos que colonizaron estos domos fueron: macroalgas (particularmente Rhodophyta), otras algas no determinadas, poliquetos serpúlidos (Annelida), esponjas (Porifera) y usualmente un espécimen solitario de pez asociado (Arothron hystrix o Diodon holocanthus). Estos arrecifes artificiales colapsaron eventualmente y desaparecieron.

Fig. 7 Arrecife tipo reefball (domo) ubicado sobre fondo arenoso a 6 m de profundidad en ANP Complejo Los Cóbanos. Aproximadamente a 3-4 meses de su colocación. / Fig. 7. Reefball (dome) located on sandy floor at 6 m deep at Los Cóbanos NPA. 3-4 months after placement approximately.
Todos los proyectos y actividades para el manejo sostenible de recursos del ANP Complejo Los Cóbanos y para la restauración directa e indirecta de la biodiversidad marina (realizadas por entidades estatales, organizaciones no gubernamentales, academia, empresa privada y cooperación internacional), han facilitado la protección de estas especies y mejorado las capacidades institucionales pertinentes, sin embargo, los eventos severos de blanqueamiento recientes han causado mortalidad de P. lobata. Además, los arrecifes tipo domo fueron colonizados con una reducida variedad de especies, excluyendo corales de cualquier tipo.
Discusión
Blanqueamiento: Los episodios de blanqueamiento severo de amplia cobertura que experimentaron las colonias someras aledañas al área denominada El Faro, ocurrieron entre 2015 a 2020, particularmente en el segundo semestre de cada año. Las colonias submareales también presentaron un evento severo de pérdida de zooxantelas entre octubre 2015 a marzo de 2016. Estos sucesos coincidieron con el blanqueamiento global de corales registrado entre 2015 a 2016 con variaciones temporales locales (Alvarado et al., 2020; Hughes et al., 2018; Monroe et al., 2018; Sully et al., 2019). En ambos casos, el área de las colonias de coral con pérdida de tejido rápidamente fue colonizada por macroalgas no determinadas (Fig. 1D). Se ha señalado como causa principal del blanqueamiento reciente en corales duros a cambios en el ambiente físico, particularmente por el calentamiento de los océanos tropicales, exposición solar y también a agentes patógenos (Baker, 2004; Stoskopf, 2011; Van Oppen & Lough, 2018). También se incluyen los sedimentos suspendidos que pueden cubrir corales (Sáez-Moreno, 2020). El género Porites también ha experimentado pérdida del alga simbionte (blanqueamiento) y mortalidad en diferentes partes del mundo (Cantin & Lough, 2014; Furby et al., 2017). La especie Porites lobata ha presentado estos episodios de pérdida de coloración en el océano Pacífico Oriental incluyendo Costa Rica (Boulay et al., 2013), Hawái (Kramer et al., 2016), golfo de Arabia y Mar Rojo (Hadaidi et al., 2017).
También las floraciones de algas nocivas afectan a los corales formadores de arrecife en varias regiones del planeta por ejemplo golfo de Omán (Bauman et al., 2010), Tailandia (Thawonsode et al., 2015), costa Pacífica de Costa Rica (Méndez-Vanegas et al., 2021). Floraciones causadas principalmente por el dinoflagelado M. polykrikoides provocaron pérdida de tejido y blanqueamiento de P. lobata en ambas áreas en diferentes períodos, incluyendo mayo de 2012 (Espinoza et al., 2012; Espinoza et al., 2013) y marzo de 2022 (Genovés, 2022; MARN, 2022).
Los registros de temperatura del agua superficial presentaron coincidencias cronológicas de los picos más altos ocurridos en septiembre de 2015 (Fig. 6A) con los episodios de blanqueamiento severo de las colonias aledañas a playa El Faro (septiembre-diciembre 2015, Tabla 1), coincidiendo con hallazgos de varias investigaciones que concluyen que el estrés por calor en el agua ha causado y causará eventos de blanqueamiento en Porites spp. en diferentes regiones (Carilli et al., 2012; Donner & Carilli, 2019; Mumby et al., 2001). En el caso de las colonias submareales no pudo establecerse coincidencias de temperatura local con los blanqueamientos severos debido a falta de información del parámetro (Fig. 3, Fig. 6B). Sin embargo, el evento severo de pérdida de zooxantelas que estas colonias experimentaron entre octubre 2015 y marzo 2016 ocurrió simultáneamente, aunque tardaron más en recuperarse, además, la pérdida de cobertura de coral vivo fue mínima (menos de 5 % aproximadamente). Esta situación refleja que las colonias someras están adaptadas a mayor estrés por agua con temperatura elevada e irradiación solar durante las mareas negativas que ocurren cinco a seis veces por año en la época seca principalmente, sin embargo, experimentan mortalidades importantes y recuperación relativamente rápida (menos de cuatro meses), en cambio las submareales que responden con menor intensidad a los factores estresores mencionados anteriormente, presentan un mecanismo de recuperación lento. Esto podría estar vinculado también a la resiliencia térmica de las algas simbiontes (Barshis et al., 2018), posiblemente son diferentes especies o genotipos asociados a las colonias someras y submareales.
Los dos parámetros de fluorescencia de clorofila utilizados en este estudio (Φ y FV/FMAX) reflejaron una baja eficiencia de la fotosíntesis de las zooxantelas simbiontes de P. lobata entre agosto y octubre de 2015 (Fig. 4, Fig. 5). Estos registros ocurrieron simultáneamente con los períodos de blanqueamiento (Tabla 1, Fig. 3). Estos bajos registros de fluorescencia de clorofila confirmaron que los procesos fotosintéticos de las algas simbiontes de P. lobata experimentaron perturbación causada posiblemente por temperatura elevada del agua de mar principalmente, así como irradiación solar u otra condición ambiental estresante (Hoogenboom et al., 2012; Wangpraseurt et al., 2019; Warner et al., 2010).
Restauración indirecta y directa: En la sección de resultados de este documento se presentaron los esfuerzos institucionales, legales, fortalecimiento de gobernanza y otros afines, como actividades indirectas para permitir la restauración indirecta y directa del arrecife de Los Cóbanos, que incluyó corales.
La Ley de Medio Ambiente aprobada en 1998 declaró en el Artículo 74 a los arrecifes como ecosistemas frágiles y reserva ecológica por lo que no se permite alteración en ellos. Este artículo ha permitido a las autoridades nacionales proteger con respaldo legal a los arrecifes. En la misma ley existen Artículos (16-29) que requieren la obtención de un permiso ambiental previo a la ejecución de obra física. Este procedimiento incluye la elaboración de un estudio de impacto ambiental en el caso de proyectos ambientalmente sensibles con incidencia en los arrecifes (Turner et al., 2006). Esta legislación ha permitido al MARN (Ministerio de Medio Ambiente y Recursos Naturales) regular actividades en playas, incluyendo las cercanas a arrecifes. Sin embargo, los monitoreos de aguas residuales por parte de las autoridades son escasos o no se realizan, por lo que se desconoce si las colonias de P. lobata en Los Cóbanos son afectadas por estos desechos líquidos. En otras partes del mundo, como Guam (Redding et al., 2013) y Hawai, (Yoshioka et al., 2016), se han encontrado alteraciones de salud y morfología de esta especie y otras del mismo género.
La aprobación e implementación de la Ley de Áreas Naturales Protegidas en 2005 facultó al MARN declarar área natural protegida a Los Cóbanos mediante decreto ejecutivo No. 22 publicado en el Diario Oficial 29 del martes 12 de febrero de 2008. Con este respaldo legal, ese ministerio pudo administrar esa área de 21 312 ha de superficie marina, fondos arenosos y parches rocosos, que también abarca pequeñas áreas terrestres de bosque tropical seco y manglar (576 ha). Ello permitió la coadministración del área en conjunto con organizaciones no gubernamentales con capacidad técnica.
También, fondos internacionales se han destinado para fortalecer las capacidades de gobernanza y desarrollo sostenible en el área, posiblemente desde antes de la aprobación de la Ley de áreas naturales protegidas. Estas fuentes facilitaron el comienzo de la gestión ambiental del área que incluyó contratación de personal local para participar como guarda recursos por una ONG local. Esto comenzó a reducir las extracciones desordenadas y no autorizadas de organismos marinos, particularmente corales negros (Antipathidae) y posiblemente colonias de Psammocora spp. Algunos de ellos aún laboran en esa unidad actualmente administrada por el MARN que cuenta con 13 personas aproximadamente, aunque se desconoce si será en forma temporal o permanente. También se apoyó el financiamiento de investigaciones básicas que permitieron comprobar la ocurrencia del coral masivo P. lobata en esa ANP (Molina, 2004), tallas del caracol Titanostrombus gigas (Hernández-Rodríguez, 2006), calidad del agua de ríos confluyentes al ANP en el que se confirmó la elevada contaminación orgánica, particularmente por material fecal (Melara, 2008), entre otros.
Considerando la extracción ilegal de vida silvestre marina y que el MARN comenzó a aplicar la Ley de Conservación de Vida Silvestre mediante decreto Legislativo 441 del año 2001 (Constitución Política del Salvador, Ley de Conservación de Vida Silvestre, 2001) y con ello, la responsabilidad de proteger especies de vida silvestre de la extinción mediante la elaboración y divulgación de listados oficiales de especies amenazadas y en peligro, a partir de 2009 se incluyeron especies marinas como todos los holoturoideos, corales hermatípicos y ahermatípicos, abarcando a Porites lobata, Psammcora spp. y otros. Esto permitió limitar estas extracciones, aunque las poblaciones de corales de la familia Antipathidae y del género Psammocora ya habían sido fuertemente diezmadas al menos 10 años antes. Ambos taxones aún se encuentran presentes en el ANP (Barraza, 2008; López & Jiménez, 2008, respectivamente).
Al declarar esta ANP también como humedal de importancia internacional en 2019, las autoridades ambientales consideraron comprometerse ante esa convención internacional para procurar el desarrollo sostenible de la zona. Parte de estos esfuerzos incluyen el fortalecimiento de personal antes mencionado, así como ordenamiento del turismo en la zona, por ejemplo, el avistamiento responsable de la ballena jorobada (Megaptera novaengliae) (Fajardo, 2017) y otras acciones de manejo territorial y ordenamiento turístico.
Los esfuerzos mencionados anteriormente comprenden actividades que facilitarían la restauración indirecta de las colonias de coral, sin embargo, los eventos repetitivos de blanqueamiento en las colonias someras y las floraciones algales nocivas no han permitido procesos de restauración natural desde el año 2015 posiblemente. Aparentemente las colonias submareales no han experimentado mayor pérdida (Barraza, 2020).
Además, la colocación de 50 arrecifes tipo domo (reefball) a 5-7 m de profundidad fue parte de un esfuerzo de restauración directo en el ANP entre 2006 a 2007. Se considera que condiciones como el oleaje intenso en la zona durante mar de fondo y que en la época lluviosa el sedimento arenoso se suspende en la columna de agua, causando un efecto abrasivo a 1 m del fondo aproximadamente, durante el cual ocurrió una colonización con pocas especies de algas e invertebrados en los arrecifes tipo domo al término aproximado de 60 días de colocación (Fig. 7); asimismo se observó que el interior del domo era oclonizado por un espécimen de pez de las especies A. histrix o D. holocantus. A los seis a 12 meses después de su colocación se encontraron cubiertos por arena o colapsados. Este tipo de arrecife también fue cubierto por sedimento y mostró éxito reducido para atraer peces en comparación a piedras conglomeradas en islas del Mar Caribe oriental (Hylkema et al., 2020). Esta experiencia reflejó que este tipo de arrecife no es el más apropiado para utilizarse en actividades de restauración en esta ANP. Una experiencia incipiente para realizar restauración directa de colonias de P. lobata se ha desarrollado en Costa Rica, ya que se ha podido cultivar fragmentos de ese coral en condiciones artificiales para luego reintroducirlos al océano, sin embargo, no existen datos sobre las condiciones de oleaje y corrientes del posible lugar de restauración (Moesbergen, 2020). Esto podría constituir una posibilidad futura de restauración directa de P. lobata, Psammocora spp., Pocillopora spp. en El Salvador, con la consideración del oleaje fuerte y aparente elevada cantidad de sedimentos suspendidos en el agua durante la estación lluviosa (Reyes-Bonilla & Barraza, 2003), así como el acompañamiento de las autoridades ambientales.
Un dato final derivado de observaciones realizadas entre julio y agosto de 2022 reflejaron poca cobertura de macroalgas frondosas, así como una importante cobertura nunca observada de algas coraliáceas costrosas y ausencia de blanqueamiento en P. lobata del área somera, contrario a lo observado en ocasiones anteriores en esos meses (Tabla 1).
Las actividades de restauración directa e indirecta (Edwards & Gómez, 2007) de P. lobata, otros corales hermatípicos y ahermatípicos, así como otra biota de arrecife han permitido la protección de especies y empoderamiento de la población a nivel local y nacional, pero el oleaje del área, eventos de blanqueamientos masivos y otros estresores ambientales recurrentes no han permitido procesos de restauración completos.
Las principales causas del blanqueamiento mencionado posiblemente fueron estrés térmico, exposición solar, floraciones algales nocivas y otros factores ambientales. Desde la aprobación de la Ley de Medio Ambiente se han implementado medidas directas e indirectas relativas a la restauración de los arrecifes del país, que previnieron la extracción ilegal de vida silvestre del ANP Los Cóbanos, pero ha incidido poco en la restauración de corales hermatípicos debido a los episodios de blanqueamiento mencionados. Se requiere enfocar esfuerzos integrales en el manejo de las cuencas hidrográficas asociadas a esta área, prevención de la contaminación, así como experimentar con restauración directa e indirecta con base a experiencias exitosas desarrolladas en países cercanos.
Declaración de ética: el autor declara que este documento no presenta conflicto de interés alguno. Y se aclara que los datos se recolectaron cuando el autor laboró en el MARN y posteriormente en Universidad Francisco Gavidia.

