











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
1. Introducción
Los bosques representan el 30 % de la superficie emergida de la tierra y las plantaciones el 3.8 % del área de bosques (140 millones de ha) (FAO, 2018), la región tropical presentó una pérdida neta de bosques en el periodo 2000-2010 de 7 millones ha, mientras las plantaciones para el mismo periodo sumaron 3.1 millones de ha (FAO, 2016). El bosque plantado, desde tiempos ancestrales provee a la sociedad múltiples bienes y servicios, tales como empleo rural, conservación de la biodiversidad, protección del suelo, regulación del ciclo hidrológico, etc. (Cabrera, 2003), y más recientemente, se les reconoce su capacidad para el secuestro de carbono (Salinas y Hernández, 2008).
Gmelina arborea es una especie tropical, que ocupa actualmente, el segundo lugar en área reforestada en Costa Rica con 13 314 ha (INEC, 2018), se caracteriza por su rápido crecimiento, con un incremento medio en volumen por el orden de 20 hasta 35 m3*ha-1 (Jiménez, 2016); su madera es usada en diversidad de usos: estructural en construcciones livianas, carpintería general, empaque, tallados, muebles comunes y chapas decorativas, instrumentos musicales, palitos de fósforo, tableros de partículas, puntales de minas, carrocerías, botes y para la fabricación de productos para embalaje (Rojas et al., 2004; Jiménez, 2016).
En Costa Rica, el desarrollo de estudios tendientes a consolidar paquetes tecnológicos de silvicultura clonal para el manejo forestal de esta especie data desde los años noventa, es por ello, que la mayor parte del área hoy plantada proviene de reproducción vegetativa. Distintos autores concuerdan que, la producción clonal, a partir de un enfoque genético, potencia la productividad y la calidad de la materia prima producida por unidad de área (Kumar, 2007; Murillo et al., 2013), lo que conlleva a un incremento en la capacidad de la especie para mitigar el cambio climático y aportar sustancialmente, en la reducción de la brecha entre oferta y demanda.
La producción de biomasa para melina ha sido poco investigada, y menos para plantaciones con silvicultura intensiva; dada la importancia de la especie en Costa Rica y en concordancia con el plan de descarbonización de la economía (MINAE, 2018), se hace necesario proveer herramientas que faciliten el cálculo del stock de carbono.
Además, la cuantificación de las fracciones de biomasa es importante para el manejo sostenible y para la producción de bioenergía. Con este fin, se desarrolló un estudio cuyo objetivo fue desarrollar modelos matemáticos para estimar-predecir la biomasa y el carbono por árbol y sus componentes (hojas, ramas, tronco, raíz), y aportar información adicional como factores de expansión de biomasa, porcentaje de materia seca y fracción de carbono en la biomasa.
2. Metodología
2.1 Área de estudio
El estudio comprendió 3 zonas en Costa Rica: norte, pacífico norte y sur; en aquellos sitios en donde se ubicaron plantaciones establecidas con material clonal, con edad entre 1 y 8 años. En el norte se trabajó en los cantones Guatuso, Los Chiles, San Carlos y Upala que poseen un clima lluvioso todo el año, la precipitación media anual es de 3 056 mm y la temperatura media de 24 °C. En el pacífico norte (distrito Cóbano) la precipitación promedio es de 1 888 mm al año y la temperatura media de 27 °C. En el sur, el distrito Puerto Jiménez registra una precipitación promedio de 3 650 mm al año y la temperatura media 26 °C (Costa Rica, 2014).
2.2 Evaluación de la biomasa
Los árboles elegidos para cuantificar la biomasa se seleccionaron aleatoriamente, de forma proporcional a la frecuencia por rango diamétrico (rango de 5 cm), según el estado silvicultural de las plantaciones evaluadas. La madera de melina, aún con un diámetro pequeño que logra a temprana edad, es muy usada para fabricar productos para embalaje (tarimas), por lo cual, en la cuantificación de la biomasa del fuste se emplearon cuatro técnicas:
2.2.1 El método destructivo
Consiste en derribar el árbol y pesar en el campo (peso húmedo o verde en kg) cada componente por separado (fuste, ramas, hojas y raíz), práctica usada para árboles de tamaño pequeño sin valor comercial o en árboles de porte mayor, pero que el propietario permitió subdividirlo en secciones pequeñas para facilitar su pesado. De cada componente se tomó una muestra de aproximadamente 1 kg para determinar la materia seca (a 60 °C durante 72 horas) y la fracción de carbono (Fonseca, Alice y Rey-Benayas, 2009). La cantidad de árboles cortados fue de 85.
2.2.2 El método del análisis fustal
Se utiliza para obtener información sobre el crecimiento en volumen del árbol a través de los años. En este caso el mismo fue derribado y se pesó en campo sus componentes, luego se extrajeron discos a cada metro sobre su eje hasta el ápice. Se cubicó el árbol en secciones de un metro con la fórmula de Smalian y la biomasa se calculó como el producto del volumen por la densidad específica de la madera (Chave et al., 2014).
La densidad específica usada para la especie fue de 0.4 gr/cm3, cifra mayormente citada en la literatura (Rojas et al., 2004; Muñoz y Berrocal, 2005). La biomasa total del árbol (incluida ramas, hojas y raíz), a diferente edad, se calculó haciendo uso de factores expansión (Fonseca et al., 2009) calculados para los árboles derribados, según lo indicado en el apartado 2.2.1.
A cada árbol, independientemente de la técnica usada para determinar la biomasa, se le midió el diámetro a la altura del pecho a 1.3 m sobre el nivel del suelo (Dap) y en cada sitio o plantación se trató de elegir individuos abarcando la amplitud diamétrica observada, según la edad. La raíz se extrajo con una retroexcavadora o pulif (tipo tecle manual que funciona de forma horizontal), posteriormente, se lavó para eliminar componentes como piedra y tierra. Se continuó con el proceso de pesado aproximadamente dos horas después de secadas al aire.
2.3 Determinación de la fracción de carbono en biomasa
La muestra seca de cada componente de biomasa se trituró a 10 micras para determinar la materia orgánica (MO). Este análisis se realizó haciendo uso de la metodología propuesta por Walkley y Black (1934).
2.4 Ajuste y selección de modelos
Los modelos se ajustaron mediante el método de mínimos cuadrados ordinarios con el programa estadístico Statgrapihcs Centurion XVI. Se ensayaron aproximadamente, 15 modelos para predecir la biomasa y el carbono de cada componente del árbol (hojas, ramas, fuste, raíz), para la biomasa leñosa del árbol (fuste, ramas y raíz) y para el individuo total, usando como variable independiente el diámetro normal (Dap).
Cuando algún modelo presentó heterogeneidad de varianza se ajustó con la función ln_robust del paquete estadístico Estmatr. En la selección de la ecuación de mejor ajuste se tomó como referencia la metodología expuesta por Salas (2002) y Segura y Andrade (2008), tomando en consideración los siguientes estadísticos: R2 (coeficiente de determinación), EEE (error estándar de los estimados), EMA (error medio absoluto), DW (estadístico Durbin Watson), IF (índice de Furnival), E % (error del modelo o sesgo), AIC (criterio de información de Akaike). Adicionalmente, se calculó el ECM (error cuadrático medio), la DA (diferencia agregada) y el PRESS (la suma de cuadrados de los residuos de los predichos), estos últimos no se muestran en los resultados para simplificar el cuadro dónde aparecen los otros estadígrafos.
3. Resultados
3.1 Biomasa del árbol y de sus componentes (factor de expansión de biomasa)
En el estudio se utilizaron 125 observaciones, provenientes de mediciones de biomasa directas e indirectas, abarcando la amplitud diamétrica observada en las plantaciones evaluadas que osciló entre 3.9 y 39.8 cm. La representación de la muestra por clase de tamaño varió entre 10 % y 32 % para las clases diamétricas de entre 5 y 30 cm y menor a 5 % para los rangos mayor a 30 y menor de 5 cm.
El fuste tiene el mayor aporte (65.6 %) a la biomasa total del árbol, seguido por la raíz y las ramas, la contribución de las hojas fue muy baja (3.15 %). La variabilidad observada entre el aporte porcentual de cada componente a la biomasa total fue alta, más baja en hojas y ramas, con cifras muy similares; en el fuste y la raíz se presenta mayor variabilidad, ambos con valores prácticamente iguales (Cuadro 1).
3.2 Materia seca y fracción de carbono por componente
La fracción de materia seca varió de 0.32 (SD=0.02) en hojas a 0.44 (SD=0,06) en ramas, mientras el carbono de 0.39 (SD=2.7) en hojas a 0.48 (SD=3.3) en fuste; en ambos casos existe muy poca variabilidad en la muestra lo que resulta en errores de muestreo inferiores al 4 % para la materia seca y máximo 8 % para la fracción de carbono (Cuadro 2), en este caso, el reducido tamaño de la muestra (9) puedo haber incrementado este valor.
Cuadro 1 Componentes utilizados para desarrollar las ecuaciones de biomasa. Entre paréntesis y en negrita, el aporte porcentual de cada componente a la biomasa total.
| Valor | Variable | ||||||
|---|---|---|---|---|---|---|---|
| Dap (cm) | Biomasa por componente (kg) | ||||||
| Hojas | Ramas | Fuste | Raíz | Componente leñoso | Biomasa total | ||
| N | 125 | 125 | 125 | 125 | 125 | 125 | 125 |
| Mínimo | 3.9 | 0.37 | 1.07 | 4.66 | 1.51 | 7.62 | 7.25 |
| X | 18.69 | 9.03 (3.15) | 29.63 (10.33) | 188.00 (65.60) | 59.92 (20.91) | 277.55 (96.85) | 286.58 |
| Máximo | 38.9 | 51.43 | 169.22 | 1598.12 | 518.22 | 2285.73 | 2337.16 |
| SD | 7.69 | 9.56 | 35.23 | 286.45 | 92.98 | 412.42 | 420.13 |
| E % | 7.36 | 18.93 | 21.27 | 27.26 | 27.76 | 26.58 | 26.22 |
n: tamaño de la muestra, X: promedio, SD: desviación estándar, E %: error relativo de muestreo.
Cuadro 2 Materia seca y fracción de carbono (entre paréntesis) por componente.
| Estadístico | Materia seca | |||
|---|---|---|---|---|
| hojas | ramas | fuste | raíz | |
| n | 46 (20) | 46(20) | 46 (21) | 34(9) |
| X | 0.32(0.39) | 0.44(0.46) | 0.37(0.48) | 0.37(0.45) |
| SD | 0.02(2.7) | 0.06(3.2) | 0.03(3.3) | 0.05(4.6) |
| E % | 2.13(3.2) | 3.99(3.2) | 2.80(3.0) | 4.60(7.9) |
X: promedio, SD: desviación estándar, E %: error relativo de muestreo.
3.3 Modelos predictivos de biomasa y carbono
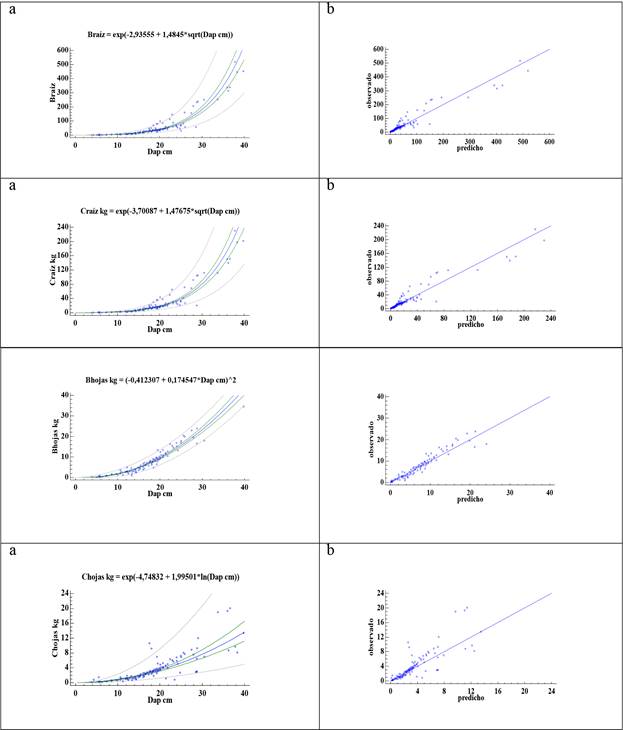
Los modelos elegidos con sus respectivos estadísticos se presentan en el Cuadro 3 . El diámetro, como variable independiente mostró una correlación muy alta (R>0.96) con la biomasa y el carbono, excepto con el carbono en hojas que fue de 0.88. Para todos los modelos los parámetros de la regresión fueron significativos (α = 0.05), lo cual indica que el Dap como variable independiente explica significativamente, la predicación de variable dependiente (biomasa o carbono).
El EEE y EMA poseen valores muy similares y muy bajos, que significa un error máximo en la estimación de 0.49 kg por árbol, según sea el componente evaluado. Los resultados del coeficiente de determinación (R2) indicaron que el diámetro explicó entre 91 % y 93 % de la varianza observada en la biomasa (el valor más para el componente ramas) y entre 78.2 % y 93.7 % de la varianza observada en el carbono (el valor más para el componente hojas).
La prueba de Durbin Watson y el método gráfico (no existe evidencia de un comportamiento sistemático en la distribución de los residuos) indican que no existe autocorrelación entre los residuos, es decir, no se viola el principio de normalidad. El Índice de Fournival, calculado para comparar modelos lineales con otros con variable dependiente transformada, muestra valores bajos, así como el AIC con un valor máximo de 4.93.
La bondad de predicción de los modelos evaluada en función DA (considerado como sesgo) y el error (la raíz cuadrada de ECM), como medidas de variabilidad presentan valores bajos que se ven reflejados en errores también bajos, el más alto de 10.9 % para carbono en hojas, esto se comprueba con la Figura 1b, al observarse, que en todos los casos, la nube de puntos se ubica muy cerca de la línea central indicando que los modelos generan estimaciones con poco sesgo.
Cuadro 3 Modelos para estimar biomasa (kg) y carbono (kg) para distintos componentes del árbol, para el árbol completo y volumen (m3) del fuste. El tamaño de la muestra fue de 125 y todos los modelos poseen una P<0,0001.
| Modelo | R2 | EEE | EMA | DW | ECM | DA | IF | E% | AIC |
|---|---|---|---|---|---|---|---|---|---|
| Btotal = exp(0.641384 + 2.75382*Log10(Dap )^2) | 93.0 | 0.3642 | 0.2667 | 1.0432 | 127.4 | 15.4 | 48.3 | 6.5 | 102.9 |
| Ctotal = exp(-1.78291 + 1.41272*√Dap) | 92.4 | 0.3637 | 0.2679 | 1.1872 | 63.6 | 2.6 | 24.4 | 5.7 | 100.1 |
| Bleñosa = exp(-1.25398 + 1.44722*√Dap) | 93.0 | 0.3761 | 0.2776 | 1.2104 | 137.6 | 0.5 | 46.6 | 8.5 | 111.6 |
| Cleñoso = exp(-1.87626 + 1.42962*√Dap )) | 93.4 | 0.3383 | 0.2557 | 1.1994 | 62.0 | 1.3 | 20.7 | 5.5 | 84.2 |
| Bfuste = exp(-1.65997 + 1.44652*√Dap) | 93.0 | 0.3634 | 0.2715 | 1.0241 | 88.9 | 6.0 | 31.2 | 5.2 | 100.8 |
| Cfuste = exp(-2.44019 + 1.469*√Dap) | 93.7 | 0.3464 | 0.2633 | 1.1543 | 45.8 | 0.4 | 14.5 | 5.7 | 97.9 |
| Bramas = exp(-4.09297 + 2.46917*ln(Dap)) | 91.0 | 0.3761 | 0.2766 | 0.9682 | 13.6 | 2.4 | 5.0 | 7.2 | |
| Cramas = exp(-4.87134 + 2.4727*ln(Dap )) | 91.6 | 0.3723 | 0.2756 | 0.9814 | 6.3 | 1.1 | 2.9 | 6.9 | |
| Braíz = exp(-2.93555 + 1.4845*√Dap) | 93.0 | 0.3681 | 0.2779 | 1.3056 | 31.7 | 64.9 | 4.4 | 5.8 | 103.0 |
| Craíz = exp(-3.70087 + 1.47675*√Dap) | 93.4 | 0.3563 | 0.2669 | 1.2242 | 13.4 | 0.3 | 4.4 | 3.5 | 93.8 |
| Bhojas = (-0.412307 + 0.174547*Dap)^2 | 93.0 | 0.2975 | 0.2273 | 1.7127 | 3.3 | 0.6 | 0.3 | 2.4 | |
| Chojas = exp(-4.74832 + 1.99501*ln(Dap)) | 78.2 | 0.4948 | 0.3357 | 1.0234 | 2.2 | 0.4 | 1.2 | 10.9 |
Btotal-C total: biomasa-carbono del árbol completo, Bleñoso-C leñoso: biomasa-carbono de componente leñoso, Bfuste-C fuste: biomasa-carbono del fuste, Bramas-C ramas: biomasa-carbono de ramas, Braíz-C raíz: biomasa-carbono de raíz, Bhojas-C hojas: biomasa-carbono de hojas, exp: exponente, Dap: diámetro normal a 1,30 m sobre el nivel del suelo (cm), ln: logaritmo natural, R, R2, EEE, EMA, DW (=0.0000), n, IF, E % y AIC definidos en la metodología.
4. Discusión de resultados
4.1 Biomasa del árbol y de sus componentes (factor de expansión de biomasa)
La proporción de los componentes en la biomasa aérea varía según las especies (Blujdea et al., 2012; Mensah et al., 2016). En este estudio, el fuste con corteza acumuló el 65.6 % (83 % de la biomasa aérea) de la biomasa total, las ramas el 15.3 %, las raíces un 20.9 %, cifra muy similar a la reportada por Jiménez et al., (2018) para la biomasa en el fuste (89.6 %) y 10.4 % el follaje y ramillas en melina de 3 años, valores muy similares a los citados por Onyekwelu (2004) para árboles entre 5 y 21 años (tallo 84 %; rama 13 % y follaje 3 %).
Mientras Swamy, Kushwaha & Puri (2004), encontraron que entre 1 y 6 años, el tallo aportó entre 55.3 % y 56.3 % (sitio 3 y 1), las ramas de 18.3 % al 19.8 % (sitio 2 y 3), las raíces del 17.9 % al 18.5 % (sitio 3 y 2) y el follaje de 6.6 % a 7.0 % (sitio 2 y 3), de la biomasa total. Según Mensah et al. (2016) el mayor aporte del fuste a la biomasa total del árbol se debe a que la biomasa leñosa generalmente, se acumula a expensas de la biomasa de la hoja, asignando más recursos para madera y ramas a medida que aumenta el tamaño, con el objetivo promover la altura y el crecimiento de la copa para aumentar la competencia por la luz con los árboles vecinos.
Información sobre la cantidad de biomasa en las raíces, en G. arborea y para otras especies, es nula o poco frecuente en la literatura, a pesar de ser un componente que acumula un porcentaje alto de la biomasa total del árbol, 20.9 % en este estudio y 14.3-15.3 % según Lin et al. (2017), para bosque natural, y de mantener el carbono almacenado por más tiempo en comparación con las ramas y el follaje. Posiblemente, la dificultad y el alto costo que conlleva la extracción del sistema radical (Schlegel, 2001, Sierra et al., 2001), desmotiva investigar este componente con más frecuencia.
La contribución de cada componente del árbol a la biomasa total en G. arborea sigue el mismo patrón reportado para especies como Pinus oocarpa, Pinus lumholtzii, Querqus rugosa y Querqus durifolia (Vargas et al., 2017).
4.2 Materia seca y fracción de carbono por componente
La materia seca correspondió a 37 % en el fuste y raíz, 44 % y 32 % en ramas y hojas, respectivamente, similar a lo reportado por Jiménez et al., 2018 (42.1 % en el fuste, 50.1 % en el follaje y 37.6 % en la corteza (siendo esta parte la que mayor humedad acumuló). Para Pinus caribaea y Cupressus lisitanica, la materia seca es más alta, corresponde a 43.7 % y 41.6 % en hojas, respectivamente, 52.3 % y 47.8 % en ramas, 50.7 % y 42.3 % en el fuste y de 44.4 % y 44.4 % en la raíz (Rojas, 2014).
La concentración promedio de carbono en los diferentes componentes varió de 39 % en hojas a 48 % en el fuste, similar a lo reportado para Tectona grandis (Fonseca et al., 2020), con 40.7 % y 45.9 % en hojas y fuste, respectivamente. Cifras que también, están dentro de lo reportado por Rojas (2014) para P. caribaea (de 47.5 % en hojas a 49.4 % en ramas) y de 32.7 % en hojas a 52.7 % en fuste para C. lusitanica.
4.3 Modelos predictivos de biomasa y carbono
Los modelos elegidos tienen solo como variable independiente el diámetro, que en diversidad de estudios ha mostrado alta correlación con la biomasa, el carbono o el volumen, mostrando ajustes (R2) muy satisfactorios (Quirós, 2015; Jiménez et al., 2018; Jiménez et al., 2019), presenta la ventaja de que su medición es muy precisa, simple de realizar, no requiere mucho conocimiento y el equipo usado para realizar la medición es simple y económico, si se compara con la medición de la altura.
Todos los modelos presentan muy buen ajuste (R2 > 91 %), excepto el carbono en hojas con un 78.2 %. Para la misma especie con 3 años de edad y proveniente de rebrotes, Jiménez et al. (2018) desarrollaron un modelo para determinar la biomasa aérea total en estado verde en función del diámetro normal y altura total y lograron un ajuste R2 mayor a 0.96. Rodríguez et al. (2018), con información de plantaciones de G. arborea con edades de 4 y 9 años, desarrollaron modelos con un R2 de 0.65 a 0.82 con valores de CME (cuadrado medio del error) inferior a 124.0.
Mientras Wayne et al. (2017) eligió el mejor con un R2 de 0.68 para árboles de 7 años de edad, e indica que la predicción de la biomasa en G. arborea, se ve afectada por factores como la densidad y el uso de material genético natural (semilla) o mejorado (clones). También, Melo (2015) desarrolló modelos de regresión múltiple, por componente y para el árbol completo, con R2 entre 0.9 y 0.99, los más bajos para hojas y ramas.
El ajuste en la biomasa de hojas y ramas en este trabajo fue muy similar al de los otros componentes (fuste, raíz), contrario de lo ocurrido en otros estudios, en donde para las ramas y hojas no lograron obtener ajustes satisfactorios (Vargas et al., 2017; Fonseca et al., 2009).
En relación con la bondad de predicción, los modelos elegidos mostraron ser muy confiables con sesgos y errores inferiores a 15.4 % y 10.9 %, respectivamente. Al respecto, Henry et al. (2010) citado por Jiménez et al. (2018), indica, para estimar biomasa en plantaciones forestales, los modelos con errores mayores a 20 % no son funcionales, porque sobre o subestiman la biomasa. Husch (1963), cita que un modelo es aceptable cuando presenta una diferencia agregada (DA) no superior o inferior a ± 5 %, y un error cuadrático medio (ECM) que no supere el 10 %.
5. Conclusiones
El aporte de cada componente del árbol, a la biomasa total, es un valor muy importante para considerar en la planificación de inventarios de carbono; queda demostrado que el follaje debería ser el componente al cual se le asigne menor esfuerzo para inventariarlo, además de que representa un carbono de muy corta permanencia. A pesar de que las ramas y hojas presentan pequeñas aportaciones al stock de carbono, son importantes componentes para el manejo sostenible, por ejemplo, la evaluación de la sostenibilidad nutricional de las plantaciones, manejo de restos y acumulación de cargas de combustible para incendios, extracción residuos para bioenergía, entre otros.
Los modelos elegidos, además de su sencillez y practicidad por usar solo el diámetro con variable independiente, presentan muy buena bondad de ajuste y de precisión, lo cual los convierte en una herramienta muy confiable para medir la capacidad de almacenamiento de una plantación forestal, siempre que se utilicen bajo condiciones similares a las de este estudio.
6. Ética y conflicto de intereses
Las personas autoras declaran que han cumplido totalmente todos los requisitos éticos y legales pertinentes, tanto durante el estudio como en la producción del manuscrito; que no hay conflicto de interés de ningún tipo; que todas las fuentes financieras se mencionan completa y claramente en la sección de agradecimientos, y que están totalmente de acuerdo con la versión final editada del artículo.

