











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkLa familia Lutjanidae en el Pacífico de Centroamérica Meridional está representada por diez especies (Bussing et al., 1994; Márcano et al., 2002; Robertson y Allen, 2002), de ellas la más importante y que se distribuye en el golfo de Nicoya es Lutjanus guttatus (Soto et al., 2009). Según Fischer et al., (1995), esta especie vive en pequeños grupos o solitaria y muy pocas veces forma cardúmenes, se alimenta principalmente de peces e invertebrados y se localiza en zonas costeras hasta los 30 m de profundidad.
Rojas (1997a) menciona que esta especie en el golfo de Nicoya se reproduce durante todo el año con dos picos máximos reproductivos en los meses de abril y octubre, mientras que Soto et al. (2009) reportan dos máximos reproductivos: uno en marzo y el otro en setiembre.
Vargas (1998) indica que L. guttatus es capturado por la flota artesanal en la parte externa del golfo de Nicoya. Araya y Vásquez (2009) afirman que las capturas del pargo mancha (Lutjanus guttatus) en las zonas 3 y 4 de este golfo variaron entre 1994 y el 2005 de 118 a 229 toneladas anuales. Soto et al. (2009) determinaron la edad de esta especie mediante la lectura de anillos de crecimiento en otolitos e indican que la clave talla-edad permite observar un traslape de las cohortes de la población a lo largo del tiempo, por lo que se debe establecer como grupo de edad máxima seis años, permitiendo esto una adecuada interpretación de las lecturas de otolitos.
Debido a que este recurso sufre altos niveles de explotación pesquera es que esta investigación busca evaluar algunos parámetros poblacionales, de reproducción y hábitos alimenticios de la población del pargo mancha que se localiza en el caladero de pesca del Área Marina de Pesca Responsable Paquera-Tambor, en el golfo de Nicoya.
Metodología
La investigación se realizó en el área Marina de Pesca Responsable Paquera-Tambor (AMPR Paquera-Tambor) de abril del 2015 a abril del 2016, se muestreó mensualmente en los caladeros de pesca del pargo mancha en esta área (Figura 1) y en el Centro de Acopio de la Asociación de Pescadores Artesanales del área; en total se analizaron 1109 especímenes.
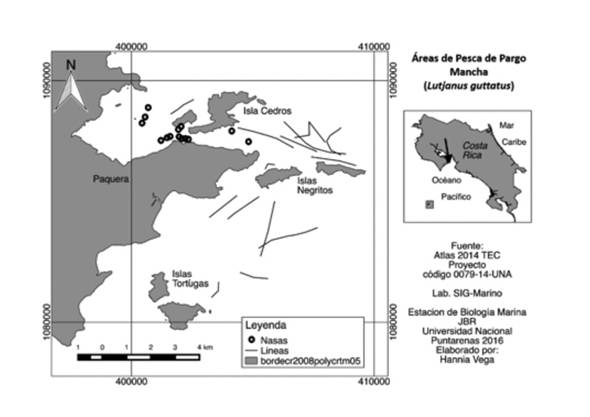
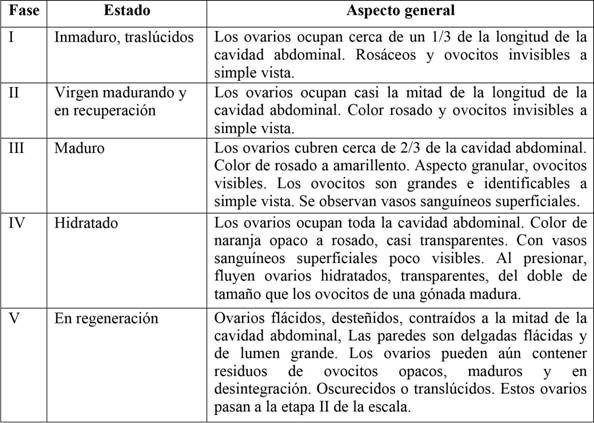
Las muestras se obtuvieron utilizando dos líneas o palangre de fondo de monofilamento de 1 mm de grosor y anzuelos tipo mustad número cinco, seis y siete; también se analizaron organismos capturados por los pescadores artesanales en el sitio de pesca; a cada espécimen se le midió la longitud total (Lt) y el peso total (Pt), se le identificó el sexo, y el estado de madurez se determinó por observación macroscópica, utilizando la escala de madurez gonadal propuesta por Holden y Raitt (1975) (Tabla 1).
Para el análisis morfométrico cada grupo de datos fue examinado mediante valores externos antes del análisis; con este fin, se utilizó el valor de entrada apropiado para el complemento “solver” en Excel (Andrade, 2003), las estimaciones no lineales fueron conducidas para obtener la ecuación potencial de las relaciones corporales P = a*(Lt) b (Sparre y Venema, 1995).
El crecimiento fue analizado considerando la metodología propuesta por Canales y Arana (2009) relativa al ajuste simultáneo de múltiples componentes modales de la composición de tallas. La longitud promedio de los individuos se estimó según la ecuación de von Bertalanffy.
L ( t ) = L∞ * 1 - exp (- K * (t - to)
K es el coeficiente de crecimiento, L∞ talla asintótica y L(t) talla a la edad t (en años); t es el tiempo y to edad teórica del organismo a la talla cero.
El valor de to se estimó por medio de la ecuación propuesta por Pauly (1984a).
Log (-to) = -0,3922 -0,2752 log L∞ (cm) -1,038 log K (año-1)
La mortalidad natural (M) se calculó mediante la ecuación de Pauly (1984b).
Log (M) = -0,0066 -0,279Log (L∞)+0,6543Log(K)+0,4634LogT°C
T es la temperatura promedio del agua, siendo 28 °C para el golfo de Nicoya (Calvo et al., 2014).
La mortalidad total “Z”, se estimó usando curvas de capturas por talla en la subrutina de FiSAT II (Gayanilo et al., 2005); la estimación de la mortalidad por pesca (F) se obtuvo de la diferencia entre Z y M (F = Z - M) (Sparre y Venema, 1995). La tasa de explotación se estimó mediante la generalización de Gulland (1983), E = F/M(Cadima, 2003). El grado de explotación se definió de acuerdo con el criterio de Gulland y Carroz (1968); si E = 0,5, la población está en el punto óptimo de explotación; si E < 0,5, la población está subexplotada, y si E > 0,5, la población está sobreexplotada.
La talla media de madurez sexual (TMMS) se determinó agrupando los individuos por sexo y, posteriormente, por clase de longitud total, con la finalidad de obtener la frecuencia acumulada estimada en porcentaje de individuos maduros (estadio tres o mayor) para cada intervalo de longitud; luego se graficó la frecuencia acumulada porcentual contra los intervalos de longitud. De la curva resultante, se consideró como talla media de madurez sexual la correspondiente a la intersección de la curva en la frecuencia del 50 % (Montreuil et al., 1998).
Para conocer los hábitos alimenticios de la especie en la zona, se analizaron 203 estómagos, a cada uno se le determinó el grado de digestibilidad del contenido estomacal; además, se identificaron los ítems al taxón más bajo posible; en los casos en los cuales se denota “restos”, se debe a que el grado de digestión del ítem solo permitió su identificación a un taxón muy general. Mientras, el material totalmente digerido se clasificó como “material digerido”.
A los ítems identificados se les aplicó los siguientes métodos: A. Método gravimétrico (% W): consiste en pesar cada taxón encontrado en cada estómago, para luego hacer una sumatoria por clase de ítem de todas las muestras; los resultados se expresan como porcentajes del peso total de alimento para todas las muestras analizadas (Hyslop, 1980). B. Frecuencia de ocurrencia (% FO): en este método se establece el número de muestras en las que se encuentran cada una de las diferentes clases del taxón alimenticio (Starck y Schroeder, 1970). C. Numérico (% N): se realiza un conteo del número de individuos encontrados en cada clase del taxón alimenticio presente en cada ejemplar. Posteriormente, se obtiene el número de individuos de cada clase de alimento .
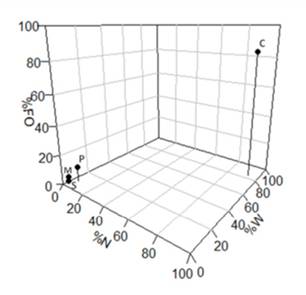
Los parámetros obtenidos (% W, % FO y % N) fueron utilizados para calcular la importancia y composición de la dieta de los peces, con base en el índice de importancia relativa (IRI) propuesto por Pinkas et al., (1971): IRI = (% N + % W) * % FO y el cálculo del índice de coeficiente de alimentación Q = % N * % W. Además, se agruparon los ítems individuales en grupos más generales y se graficaron mediante el método propuesto por Costello (1990), aplicando la modificación propuesta por Cortés (1997), esto con el fin de resaltar la importancia de distintos grupos en la dieta del pargo mancha. Con el afán de estimar la cantidad de estómagos necesarios para reflejar adecuadamente los hábitos alimenticios de pargo mancha, se construyó una curva de acumulación de presas, seleccionando aleatoriamente los 203 estómagos analizados.
Resultados
La relación longitud-peso se estimó en Pt = 0,0237X2.80 (Figura 2), indicando que esta especie presenta un crecimiento alométrico negativo; además, se determinó que tal población está compuesta por cuatro cohortes, lo que hace prever que en cualquier mes del año se pueden capturar individuos de cuatro grupos de tallas diferentes (Figura 3).
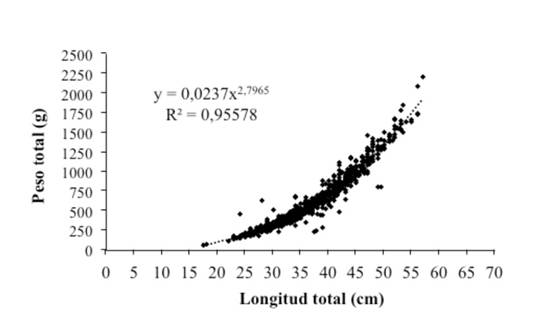
Figura 2 Relación longitud-peso del pargo mancha en el Área Marina de Pesca Responsable de Paquera-Tambor, golfo de Nicoya
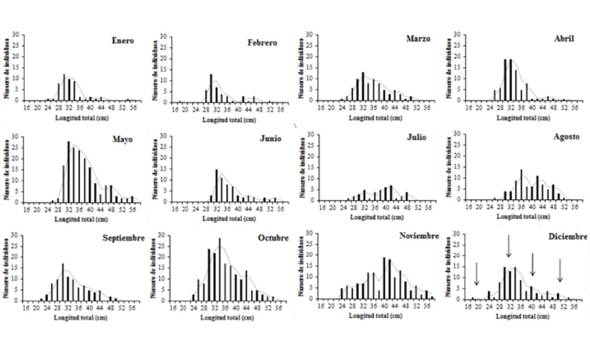
Figura 3 Distribución de frecuencia de tallas del pargo mancha en el Área Marina de Pesca Responsable de Paquera-Tambor, golfo de Nicoya. Las flechas indican las cuatro cohortes presentes en esta población.
La población en estudio presenta una proporción sexual de un macho por cada hembra (1:1), además muestra desoves continuos durante todo el año, con dos máximos reproductivos: el primero de mayo a agosto, coincidiendo con el inicio de la época lluviosa, y el segundo de octubre a noviembre (inicio de la época seca) (Figura 4). La talla media de primera madurez sexual para esta población fue estimada en 30 cm de longitud total, talla en la cual al menos el 50 % de los individuos maduran por primera vez (Figura 5).
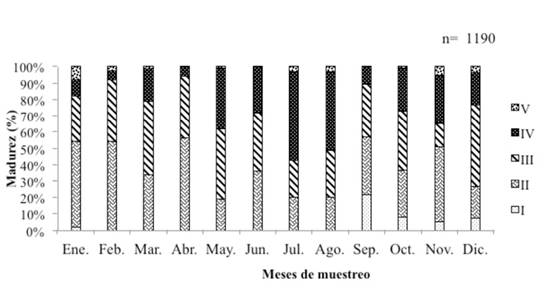
Figura 4 Periodos de desove del pargo mancha Lutjanus guttatus en el Área Marina de Pesca Responsable de Paquera-Tambor, golfo de Nicoya
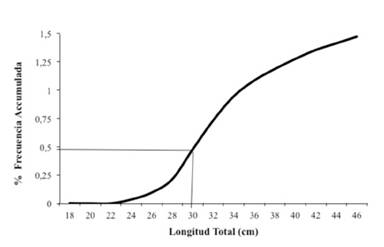
Figura 5 Talla media de madurez sexual del pargo mancha Lutjanus guttatus en el Área Marina de Pesca Responsable de Paquera-Tambor, golfo de Nicoya
Relacionando el dato estimado para la TMMS con la distribución de frecuencia por talla, se demuestra que aproximadamente el 60 % de los especímenes analizados presenta longitudes superiores a esta, mientras que un 40 % de los organismos capturados presenta talla menores a los 30 cm de longitud total (Figura 6).
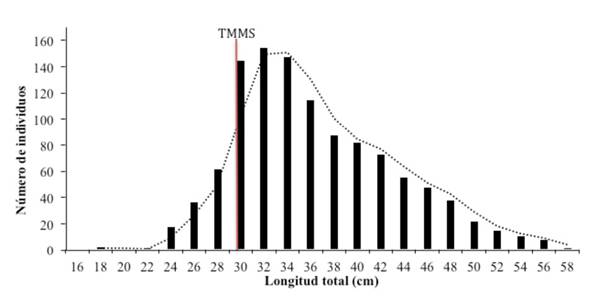
Figura 6 Distribución de frecuencia de longitud y talla media de madurez sexual del pargo mancha Lutjanus guttatus en el Área Marina de Pesca Responsable de Paquera-Tambor, golfo de Nicoya
Los parámetros de crecimiento se estimaron en 65 cm de longitud asintótica (L∞); el parámetro de curvatura (K), en 0,32 anual, y el t0 igual a -0,42. Al incluir estos valores en el modelo de crecimiento von Bertalanffy, se generó la curva de crecimiento en longitud (Figura 7), en esta se puede observar que los individuos de la población alcanzan su talla máxima a los ocho años de edad aproximadamente.
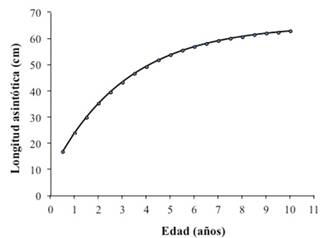
Figura 7 Curva de crecimiento del pargo mancha Lutjanus guttatus en el Área Marina de Pesca Responsable de Paquera-Tambor, golfo de Nicoya
La tasa instantánea de mortalidad natural (M) se estimó en 0,68, la de mortalidad por pesca (F) en 2,38 y la de mortalidad total (Z) en 3,06, dando como resultado un índice de explotación (E) igual a 0,77; esto último indica que la población se encuentra en estado de sobreexplotación.
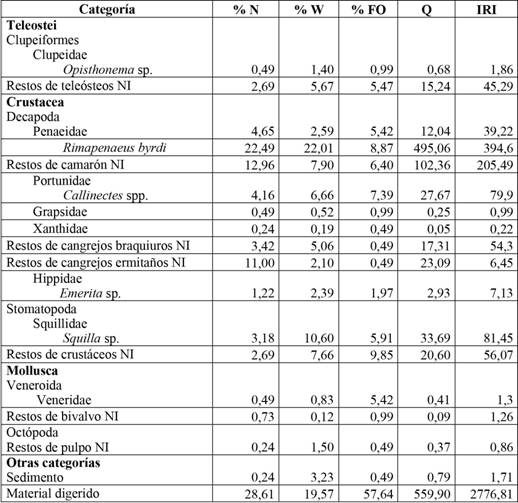
Se analizó el contenido de 203 estómagos de pargo mancha, la curva de acumulación de presas comienza a estabilizarse aproximadamente en 70 estómagos, lo cual refleja que el número de estómagos analizados brinda una estimación representativa de la dieta del pargo mancha en la zona (Figura 8). La dieta de L. guttatus en esta región se compone principalmente de crustáceos; sin embargo, no hay una clara evidencia de preferencia por uno u otro ítem en específico. Se puede observar que el camarón carabalí (Rimapenaeus byrdi) es el ítem alimentario que presenta el valor más alto del índice de importancia relativa (IRI), igual a 394,60, seguido por restos de camarones no identificados con un IRI de 205,49. En tercer lugar estuvo el género Squilla spp., cuyo IRI es igual a 81,45, seguido de Callinectes spp. el cual presenta un IRI igual a 79,9; los ítems restos de crustáceos no identificados y de cangrejo braquiuro no identificado presentan valores de IRI iguales a 56,07 y 54,30, respectivamente; los demás ítems se pueden ubicar en un segundo grupo de importancia (Cuadro 2). Es importante aclarar que los ítems categorizados como “restos no identificados” (peces, pulpo, camarón, cangrejos, crustáceos y bivalvo) se deben a que no se logró identificar a un taxón más específico, por el grado de digestibilidad que presentaron, aunque sí se alcanzó identificar algunas estructuras propias de estos organismos.
Al comparar los taxones crustáceos, peces y moluscos junto con la categoría sedimento, se identifica una fuerte dominancia del grupo de los crustáceos en la dieta del pargo mancha, ya que representa más del 80 % en los tres parámetros (% FO, % N y % W). En segundo lugar, se encuentran los peces; sin embargo, no superan el 10 % de ninguna de las categorías. Los moluscos y el sedimento se pueden clasificar como grupos raros, pues muestran un bajo porcentaje en frecuencia y en abundancia (Figura 9).
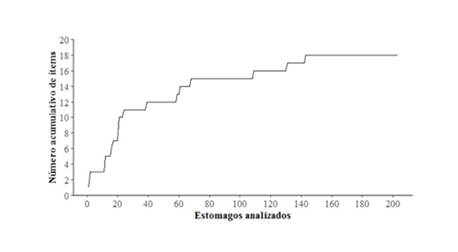
Tabla 2 Porcentaje en número (% N), porcentaje en peso (% W) coeficiente de alimentación (Q), porcentaje de frecuencia de ocurrencia (% FO) e índice de importancia relativa (IRI) para Lutjanus guttatus en el Área Marina de Pesca Responsable Paquera-Tambor, golfo de Nicoya
Discusión
El crecimiento alométrico negativo en la relación longitud-peso obtenida en esta investigación coincide con resultados reportados por Vargas (1998) y Soto et. al., (2009), los cuales estimaron un valor de “b” igual a 2,87, ambos estudios realizados en el golfo de Nicoya. Nieto-Navarro et al. (2010), para la población del golfo de California, estimaron este valor en 2,89 y Rojo-Vásquez et al., (2009) igual a 2,82 para la población del Pacífico Central Mexicano; estos resultados evidencian que el crecimiento alométrico negativo es característico del pargo mancha. Según King (1995) y Froese y Pauly (2007), esta alometría indica que la especie presenta un crecimiento volumétrico no proporcional, es decir, sin una fuerte tendencia cúbica.
Los máximos reproductivos estimados en esta investigación, el primero de mayo a agosto (inicio de la época lluviosa) y el segundo de octubre a noviembre (inicio de la época seca), coinciden parcialmente con los resultados obtenidos por Soto et al., (2009), ya que ellos reportan los meses de marzo y setiembre como periodos de máxima reproducción para esta misma especie en el golfo de Nicoya; tal diferencia puede ser producto de la metodología utilizada, debido a que ellos realizaron la estimación mediante análisis del índice gonadosomático. Mientras, Rojas (1997a) estimó abril y noviembre como meses de máxima reproducción del pargo mancha en el golfo de Nicoya.
La TMMS coincide parcialmente con lo reportado por Vargas (1998) para esta especie, quien estimó la talla media de madurez sexual en 33 ± 3 cm de Lt, mientras que Rojas (1997a) reporta que los individuos de esta especie en el golfo de Nicoya alcanzan la madurez sexual entre 31,7 y 34,3 cm de longitud total. De acuerdo a Smith (1996); la disminución en la talla media de madurez sexual, puede ser producto de la variación genética que sufre la población en respuesta a la presión pesquera. Los especímenes analizados con tallas iguales o menores a los 30 cm de longitud total evidencian que estos organismos se reclutan en los caladeros de pesca, sin haber alcanzado su talla media de madurez sexual, aspecto importante por considerar en futuras medidas de manejo.
Los parámetros de crecimiento estimados en esta investigación coinciden con los reportados por Vargas (1998), ambos estudios obtuvieron un valor de L∞ igual a 65 cm y una constante de crecimiento (K) de 0,30 anual. Sin embargo, difieren un poco de las estimaciones de Soto et al., (2009). Esta última indagación estimó la L∞ en 65,9 cm y K igual a 0,13 anual, diferencias que pueden ser producto de las metodologías utilizadas y de la presión pesquera ejercida sobre la población en estudio. El índice de explotación obtenido de 0,77 indica que esta población en el golfo de Nicoya se encuentra en estado de sobreexplotación, lo que hace urgente la implementación de medidas de manejo que minimicen ese impacto y permitan la recuperación de la población.
Los resultados obtenidos en este estudio y en anteriores investigaciones (Rojas, 1997b; Rojas-Herrera et al., 2004; Rojas et al., 2004) sobre hábitos alimenticios, sugieren que el pargo mancha es una especie generalista con preferencias por crustáceos y, en menor medida, por peces. Además, al tener capacidad de ingerir un amplio rango de presas, sus preferencias alimenticias podrían estar definidas en mayor medida por la disponibilidad de presas en una determinada región. Sin embargo, no se puede descartar que esta especie muestre alguna preferencia por un tamaño o tipo de presa en específico (Rojas-Herrera et al., 2004)

