











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkDe acuerdo con MARENA (1999), en Costa Rica y otros países de América Central, la actividad agropecuaria es la principal causa de deforestación. Para el año 2014, el área total con uso agropecuario fue de 2 406 418,4 hectáreas, lo que representa el 47,1% del territorio nacional (INEC, 2015).
Conjuntamente, la conversión de los bosques a la agricultura en Mesoamérica ha transformado los paisajes boscosos en paisajes fragmentados y dominados por pasturas y tierras agrícolas (Harvey, Alpízar, Chacón, & Madrigal, 2005). Asimismo, dichos cambios pueden tener un impacto negativo en las comunidades de aves, al reducir hábitats naturales, aislar poblaciones de aves, disminuir el tamaño de las poblaciones y aumentar la depredación en los fragmentos de bosque remanentes (Stiles, 1985). Además, estos cambios pueden provocar la extinción local o regional de especies que dependen de bosques continuos y resultar en cambios en la composición de las especies, a través de la colonización de especies oportunistas (Kattan, 2002).
No obstante, estudios recientes han demostrado que ciertos usos de la tierra tienen el potencial para proporcionar hábitat y otros recursos para una variedad de plantas y animales, lo que podría contribuir significativamente a los esfuerzos de conservación de las mismas en paisajes fragmentados (Estrada, Coates, & Meritt, 1997). Conjuntamente, Haber (1999) señala que los árboles en los potreros cumplen un papel importante en la conservación de especies de aves silvestres en paisajes fragmentados, al proporcionar refugio, sitios de descanso, anidación y alimento.
Además, las cercas vivas, cortinas rompe vientos y fragmentos de bosque también pueden servir como corredores biológicos para algunas especies en paisajes agropecuarios, mejorando la conectividad del paisaje y ayudando al desplazamiento de las aves silvestres entre parches de bosques naturales remanentes (Areskoug, 2001). Sin embargo cada localidad es muy independiente y puede tener diferentes comportamientos por lo cual se debe estudiar independientemente.
La presente investigación tuvo como objetivo determinar la composición y riqueza de avifauna en cuatro tipos de cobertura vegetal (bosque ribereño, pastura natural con alta densidad de árboles, pastura natural con baja densidad de árboles y cercas vivas,) en la comunidad del Invu, distrito de Peñas Blancas, San Ramón, Costa Rica. Y como objetivos específicos: estimar la diversidad alfa de especies para cada tipo de cobertura vegetal, analizar la tasa de recambio de especies en los cuatro tipos de cobertura y describir el grado de dependencia de bosque, especies indicadoras y los gremios tróficos de las especies observadas.
MATERIALES Y MÉTODOS
El estudio se realizó en la comunidad del Invu, distrito Peñas Blancas, San Ramón, Alajuela, Costa Rica. Se encuentra a 116km de San José y a altitud promedio de 160msnm. La zona se caracteriza por tener una temperatura promedio anual de 26°C y una precipitación media anual de 3500mm, con una estación lluviosa de mayo a diciembre, y una estación seca de enero a abril. Los fragmentos de bosques naturales remanentes y la vegetación nativa predominante pertenecen a la zona de vida del Bosque Húmedo Tropical (Holdridge, 1987).
Actualmente, el paisaje está caracterizado por la actividad agropecuaria, donde se desarrollan principalmente el cultivo de tubérculos, plátano y ganadería.
En el muestreo de avifauna se estudiaron cuatro tipos de hábitats dominantes en el paisaje: bosque ribereño (BR), pastura natural de alta cobertura de árboles (16-25% de cobertura arbórea; PAC) y pastura natural de baja cobertura de árboles (1-15% de cobertura arbórea; PBC) y cercas vivas (CV). Estos hábitats fueron seleccionados intencionalmente mediante visitas de campo a fincas que contaran con la presencia de los mismos.
En cada finca se eligió un tipo de cobertura vegetal, realizando transeptos de 300m con puntos de conteo cada 75m. Para el caso de Pasto de alta cobertura, debido a al tamaño de las fincas se usó un diseño en distinto (L y parcela cuadrada, Fig. 1). Las observaciones fueron realizadas en puntos de conteo de 25m de radio durante un tiempo de 10 minutos por punto. Cada vez que se llegó a un punto de conteo se dejaron pasar lapsos de 5 min, con el fin que las aves se acostumbrasen a nuestra presencia. Los muestreos se efectuaron entre 6:00-9:00a.m. en el periodo de mayo a agosto del 2016.
A cada individuo o grupo presente se le registró el número de punto de muestreo, la especie y el número de individuos por especie según Ralph, Geupel et al. (1996). Se calculó la riqueza, abundancia de especies. Además, se realizó un análisis de los hábitats con base en la composición de especies (presencia o ausencia), empleando el índice de similitud de Jaccard; estos índices serán calculados utilizando el software BioDiversity Pro.
Finalmente se clasificaron las aves según el gremio trófico al que pertenecen, distinguiendo entre carnívoras (incluyen carroñeras), frugívoras, granívoras, insectívoras, nectarívoras, omnívoras y piscívoras, de acuerdo con la guía de las aves de Costa Rica de Stiles y Skutch (1998). Se determinó el grado de dependencia de bosque de las especies presentes según Stiles (1985) y las especies indicadoras según Dufrene y Legendre (1997).
Ética, conflicto de intereses y declaración de financiamiento: los autores declaran haber cumplido con todos los requisitos éticos y legales pertinentes, tanto durante el estudio como en el manuscrito; que no hay conflictos de interés de ningún tipo, y que todas las fuentes financieras se detallan plena y claramente en la sección de agradecimientos. Asimismo, están de acuerdo con la versión editada final del documento. El respectivo documento legal firmado se encuentra en los archivos de la revista.
RESULTADOS
Se registraron 548 individuos pertenecientes a 27 familias y 75 especies. Las familias con mayor número de especies fueron Tyrannidae con 12 y Thraupidae con 11 respectivamente. Las cuatro especies más abundantes fueron: Ramphocelus passerinii, Turdus grayi, Crotophaga sulcirostris y Troglodytes aedon que se caracterizan por ser especies generalistas.
Riqueza y abundancia de especies: la riqueza y abundancia puntual fue mayor en pasto natural de alta cobertura vegetal y en bosque ribereño que en cercas vivas y pastos de baja cobertura vegetal.
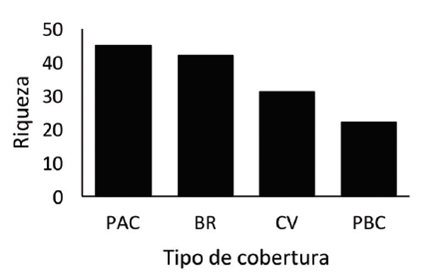
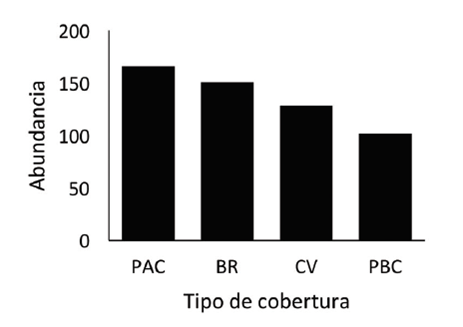
Fig. 3 Abundancia de especies según el tipo de cobertura vegetal para un paisaje de Peñas Blancas, Alajuela, 2016.
Beta diversidad: del Cuadro 1 se puede resaltar que los hábitats que tuvieron mayor similitud fueron BR-CV y PBC-CV, mientras los que tuvieron menos están representados por BR-PBC y PAC-PBC. En relación al primero se puede destacar que dichas especies compartidas en su mayoría son especies generalistas y no así las dependientes de bosque o pequeños fragmentos de los mismos. Asimismo en el caso de pastos de baja cobertura, donde se registraron especies de áreas abiertas, las cuales no frecuentan bosques como es el caso de Molothrus aeneus y Quiscalus mexicanus, lo cual hace que la similitud con otros tipos de hábitats sea menor.
CUADRO 1: Índice de similitud de Jaccard, según el número de especies compartidas con el número total de especies exclusivas para un paisaje de Peñas Blancas, Alajuela, 2016
| Hábitats | Jaccard |
| BR-PAC | 0,43 |
| BR-PBC | 0,25 |
| BR-CV | 0,61 |
| PAC-PBC | 0,29 |
| PAC-CV | 0,43 |
| PBC-CV | 0,51 |
Especies indicadoras y dependencia de bosque: se identificaron seis especies de aves indicadoras del total de 75 registradas. En el bosque ribereño dos especies, una especie para pasto de alta cobertura, dos especies para pasto de baja cobertura y una especie para cercas vivas. Estas especies presentaron valores de p<0,05, por lo que son consideradas como especies indicadoras para este tipo de cobertura vegetal, según lo plantea el índice de indicador de especies (Cuadro 2).
CUADRO 2: Especies de aves indicadoras y dependientes de bosque en un paisaje de Peñas Blancas, Alajuela, 2016
| Especie | a | b |
| Ardea alba | 3 | BR |
| Manacus candei | 2 | BR |
| Capsiempis flaveola | 3 | PAC |
| Crotophaga sulcirostris | 3 | PBC |
| Quiscalus mexicanus | 3 | PBC |
| Todirostrum cinereum | 2 y 3 | CV |
a Especies dependientes de bosque según el criterio de Stiles (1985). Clase 1 requieren de un 50% de cobertura de bosque con gran interconexión entre fragmentos de bosque continuo, especies de la Clase 2 pueden persistir en superficies con menos de un 50% de cobertura de bosque, Clase 3 no requieren de bosque y pueden persistir en paisajes agrícolas. b Especies indicadoras según el criterio de Dufrene y Legendre (1997). Con valores de p<0,05. Abreviaturas: BR, Bosque ribereño; PAC, Pasto alta cobertura; PBC, Pasto baja cobertura; CV, Cercas vivas.
Según el criterio de dependencia de bosque, se encontraron cinco especies del total de 75 especies registradas, como dependientes de bosque. De estas, la única especie con un valor de uno fue Chondrohierax uncinatus, que necesita de bosque casi continuo para subsistir y que este interconectado en un 50% con otros fragmentos de bosque. En cuanto a las otras 4 especies, están presentan un valor igual a uno y dos, por lo que necesitan de bosque casi sólido y de pequeños fragmentos de bosque para su supervivencia en el paisaje (Apéndice 1).
De estas, cuatro fueron registradas únicamente en bosque ribereño (C. uncinatus, Celeus loricatus, Piculus simplex y Henicorhina leucosticta), mientras una fue observada en pasto natural de alta cobertura arborea (Pheugopedius atrogularis). Cabe señalar la importancia que juegan este tipo de hábitats en el paisaje, a pesar de que la cobertura arborea es mayor en bosque ribereño, el grado de similitud que tiene pasto natural de alta cobertura, le permite a algunas especies desplazarse ocasionalmente a dichos sitios en busca de alimento y refugio.
Gremios tróficos: el 39% de las especies estuvo representada por el gremio insectívoro, seguido por frugívoro con 23%, omnívoro con 17%, carnívoro con 16%, granívoro con 4% y nectarívoro con 1%.
DISCUSIÓN
La composición y riqueza de aves tuvo diferencias significativas según el tipo de cobertura vegetal. Sin embargo los pastos de alta cobertura vegetal y bosques ribereños presentaron una mayor riqueza de aves que pasto natural de alta cobertura y cercas vivas. Esto podría deberse a que los primeros presentan mayor cobertura arborea, mayor disponibilidad de alimento y zonas de refugio.
Con respecto a la composición de especies, estudios similares revelan patrones semejantes a los obtenidos en este trabajo, donde las áreas con alta cobertura vegetal contienen mayor riqueza en cuanto a especies de aves, siendo de vital importancia para la conservación de las mismas (Carvajal, 2008; Estrada et al., 1997; Schulze et al., 2004; Waltert, Mardiastuti, & Muhlenberg, 2004; Cárdenas, Harvey, Ibrahim, & Finegan, 2003; Vilchez, Harvey, Sánchez, Medina, & Hernández, 2004). En cuanto a familias más abundantes (Carvajal, 2008; Pérez et al., 2006; Styles & Skutch, 1998) coinciden con Tyrannidae como la más representativa en el hemisferio occidental.
Por otra parte, el alto porcentaje de similitud entre bosque ribereño y cercas vivas, podría estar ligado con el rol que estas juegan en la conectividad de diferentes hábitats en el paisaje. Además, de las 75 especies de aves, solo 10 (Leptotila verreauxi, Tangara larvata, Thraupis episcopus, Troglodytes aedon, Turdus grayi, entre otras) estuvieron presentes en todos los hábitats; estas especies son de áreas abiertas y generalistas excepto Pteroglossus torquatus que requiere de pequeños fragmentos de bosque. Además, 15 especies de aves fueron registradas una vez en todo el muestreo, sobre todo especies de aves típicas del bosque como (Celeus loricatus, Chondrohierax uncinatus), las cuales fueron encontradas solamente en bosque ribereño.
Conjuntamente se registró Gampsonyx swainsonii, especie que según del Hoyo, Elliott y Sargatal, (1994) y Elizondo (2013) solo se encuentra en Nicaragua y desde Colombia hasta el Río Amazonas, además la misma podría encontrarse en el noroeste de Costa Rica, sin embargo para este caso se encuentra en un área distinta.
En cuanto a los gremios tróficos, en un estudio realizado en la región Huetar Norte de nuestro país, Sánchez (2009) coincide en este estudio y cita al grupo de insectívoros como el más representativo.
Finalmente cabe destacar que debido a la poca cantidad de réplicas y al corto periodo del estudio provocó que se registraran solo la mitad de las especies que han sido observadas en la zona, en días y sitios distintos al periodo de este estudio.

