











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkEl bosque húmedo premontano transición seca, se caracteriza por presentar suelos volcánicos fértiles (Fournier, Flores, & Rivera, 1985; 1991). Este tipo de suelo presentan acumulación de carbono, edafotaxon rico en bases, prevaleciendo calcio y magnesio (Peña, 2016, comm. pers.). Autores comoBergoeign y Protti (1997) indica que de los materiales parentales de estos suelos sobresalen las andesitas y rocas básicas ignimbríticas que proporcionan la característica de basicidad en los sustratos, asociados a la actividad volcánica del Poás.
Este ecosistema constituye, después del bosque tropical seco, el tipo de bosque más alterado y reducido de Costa Rica (Cascante et al.,2001; Amador, 2003; Rodríguez & Brenes, 2009). Se estima que tan solo un 21% del área original del bosque premontano transición seca se mantiene actualmente en el país. Este presenta un alto grado de fragmentación conformado por pequeños remanentes o islas de bosques (Sánchez, 2001; Cascante et al., 2012). Probablemente, dicha situación es la consecuencia de que las condiciones climáticas de este tipo de bosque son propicias para la agricultura; ganadería y el asentamiento de poblaciones humanas, por lo que se ha convertido en el área más desarrollada en términos sociales y económicos (Meave, Romero-Romero, Salas-Morales, Pérez-García, & Gallardo-Cruz, 2012; Meneses & Armbrecht, 2018).
En cuanto a las comunidades vegetales en regeneración conduce al conocimiento de la composición, estructura y dinámica de las mismas, es vital para entender cómo deben ser llevados a cabo los planes de restauración y reforestación para su regeneración (Londoño-Lemos & Torres González, 2015).
Asimismo, la estabilidad de los ecosistemas depende de la estructura, su composición y su funcionamiento (Trombulak, Omland, Robinson, Fleischner, Bromws, & Domroese, 2004; Gann & Lamb, 2006; Vargas, 2011; Vargas, Diaz, Reyes, & Gómez, 2012). La primera consiste en propiciar la presencia de ciertas especies (cuantificar riqueza) y la composición es la evaluación de la diversidad (Garibello, 2003; Rodríguez & Brenes, 2009; Cascante & Estrada, 2012; Londoño-Lemos & Torres González, 2015).
La comparación de la composición y la estructura entre dos sitios puede efectuarse mediante la aplicación de ciertas técnicas estadísticas (Dueñas, Betancur, & Galindo, 2007,Dzib, Chanatásig, & González, 2014;Muñoz, Erazo, & Armijos, 2017). Esta caracterización de la vegetación representa el primer paso hacia el entendimiento de la estructura y dinámica de un ecosistema, fundamental para comprender los diferentes aspectos ecológicos, incluyendo el manejo exitoso de los bosques tropicales (Bawa & McDade 1994; SER, 2004).
Es necesario desarrollar más investigaciones en el bosque húmedo premontano transición seca, conocer su dinámica y funcionamiento, proponer un buen manejo, así como estrategias de conectividad y restauración de esos bosques. Esto es fundamental para comprender diferentes aspectos ecológicos, incluyendo el manejo exitoso de los bosques tropicales (Bawa & McDade, 1994; SER, 2004). Por lo anterior, se evidencia la importancia de realizar estudios de composición y estructura vegetal, con el fin de caracterizarla y entender la dinámica de estos ecosistemas (Ramírez et al., 2013; Cárdenas-Torres, 2014; Caranqui, Lozano, & Reyes, 2016).
Con el fin de solventar la deficiencia de conocimiento que existe sobre el bosque húmedo premontano transición seca, se analizó la variación de la estructura y la composición del bosque debido a la actividad agrícola y ganadera. En este estudio se analiza la composición y estructura de la vegetación en un bosque húmedo premontano transición seca en dos fragmentos de bosque.
MATERIALES Y MÉTODOS
Área de estudio:la investigación se desarrolló en dos sitios, uno con un grado de alteración bajo (Andrómeda) debido a que no se realizan actividades ganaderas o comerciales destructiva y otro muy alterado (Los Llanos) por la actividad ganadera extensiva, ambos sitios se encuentran a una distancia lineal de aproximadamente 9,5km (Fig. 1).
La cobertura vegetal actual de la finca Los Llanos (Alajuela, La Garita, 9°58’44,94”N & 84°18´7,056”W, 500m.s.n.m) está compuesta por pastizales y alguna vegetación arbórea nativa muy dispersa, por lo que se puede considerar un sitio fragmentado. La degradación del suelo por el uso extensivo de la ganadería y los monocultivos ocasionó la pérdida de gran parte de la cobertura boscosa (Fig. 2).
El sitio menos alterado Andrómeda, (Alajuela, Atenas, 9°56’52,4’’ N & 84°23’5,9’’W, 700m.s.n.m.) es administrado por el Sistema Nacional de Áreas de Conservación (SINAC). Este se encuentra rodeado por fincas agrícolas con pasto y es un área en proceso de restauración pasiva (Fig. 3).
Fig. 1 Ubicación de las zonas de investigación; finca Los Llanos y bosque Andrómeda Alajuela, Costa Rica, 2016.
Análisis y recolección de datos:en cada sitio se establecieron al azar 20 parcelas circulares de muestreo de 500m2(radio 12,56m), se contabilizaron y se identificaron todas las especies de plantas con un diámetro (DAP) mayor o igual a 10cm. Para la definición de estas parcelas, se utilizó la herramienta Hawths Tools v3.27 (2006), con la cual, acoplada al programa de sistema de información geográfica, ArcGis v10.3, por medio de los módulos Create Vector Grid y el módulo Create Random Selection, se crearon cuadrículas correspondientes a las parcelas y con el segundo se hizo la selección al azar.
Una vez seleccionadas las celdas al azar, se utilizó la herramienta ETGeowizards v11.0 con el módulo Polygon to Point, con el fin de obtener la coordenada central de cada celda. Estas herramientas se utilizaron en el programa ArcMap 10.3 de Esri. Por otra parte, en ambos sitios se llevaron a cabo varios tipos de estimaciones de diversidad: a) número de especies, género y familia por parcela; b) número total de individuos por parcela y por especie; c) distribución por categoría de diámetros; d) área basal (m2) total, por parcela y por especie; e) mediciones de abundancia, frecuencia, dominancia por especie y f) el índice de valor de importancia (conocido como IVI) para cada especie, además, g) índice de importancia familiar o FIV, propuesto por Mori, Boom, de Carvalho, y Dos Santos (1983).
También se utilizaron los estimadores beta para medir el grado de cambio o reemplazo en la composición de especies entre las comunidades que se encuentran en un área mayor (Villarreal et al., 2006). Para este fin, se aplicaron: a) índice de diversidad Shannon y Wiener; b) índice de dominancia de Simpson; d) índice de equidad de Pielou (J); e) índice de similitud de Jaccard y Bray-Curtis.
Por otra parte, para los análisis de biodiversidad, se aplicaron dos procedimientos, el primero consistió en el uso de siete estimadores no paramétricos (ACE, ICE, Chao 1, Chao 2, Jackknife 1, Jackknife 2 y Bootstrap), mediante el programa EstimateS versión 7 (Colwell, 2004), obtenidos a partir la herramienta EstimateS (Versión 9.1.0), Copyright R. K. Colwell: http://purl.oclc.org/estimates. En segunda instancia, se aplicaron las curvas de acumulación de especies (Gotelli & Colwell, 2001). Ambos procedimientos estiman el número de especies que podrían existir en un sitio a partir de datos generados, los cuales sirven para tener una idea de las especies encontradas como parte de los resultados obtenidos de riqueza registrada (Villarreal et al., 2006).
Ética, conflicto de intereses y declaración de financiamiento: la autora declara haber cumplido con todos los requisitos éticos y legales pertinentes, tanto durante el estudio como en el manuscrito; que no hay conflictos de interés de ningún tipo, y que todas las fuentes financieras se detallan plena y claramente en la sección de agradecimientos. Asimismo, están de acuerdo con la versión editada final del documento. El respectivo documento legal firmado se encuentra en los archivos de la revista.
RESULTADOS
La riqueza de especies en Los Llanos fue de 16 familias, 18 especies y 76 individuos y para Andrómeda de 29 familias, 51 especies y 305 individuos. Con respecto a la distribución diamétrica de los individuos, en Los Llanos se identificaron 34 especies con DAP≥80cm y en Andrómeda, el rango con mayor cantidad de individuos fue 10-14,99cm con 91 especies (Cuadro 1).
CUADRO 1: Rangos de distribución diamétrica con mayor porcentaje de individuos identificados en Los Llanos y en Andrómeda, 2015-2017
| Los Llanos | Andrómeda |
| 15-99,99cm con 9% | 10-14,99cm con 29% |
| 30-34,99cm con 10% | 15-19,99cm con 22% |
| DAP≥80cm con 45% | 20-24,99cm con 16% |
En cuanto a la composición florística en el área basal (AB) promedio por hectárea fue de 76,60m2/ha (min.=0,007; máx.=14,52; D.E.=2,22), mientras que el promedio de Andrómeda fue de 24,69m2/ha (min.= 0,007; máx.=2,207; D.E.=0,20). Los Llanos, se ve fuertemente influenciado por la presencia de individuos de gran tamaño (DAP), como el caso de la especieFicus(dos individuos, uno con 430cm), Enterolobium cyclocarpum(presentes 16 individuos, de los cuales tres tienen diámetros de 100 a 330cm),Guazuma ulmifolia(ocho individuos, de los cuales dos de ellos supera los diámetros de 100cm),Tabebuia rosea(de los siete individuos sus diámetros oscilan entre 64 a 200cm), entre otros en diámetros que van de los 100cm hasta 430cm.
Dos especies (Enterolobium cyclocarpumyFicussp.) para Los Llanos presentaron un (AB) superior a los 12m2/ha. Asimismo, en Andrómeda fue laCedrela odoratacon 4,08m2/ha. Las familias con mayor área registrada en Los LLanos fueronFabaceae, seguido de Moraceae y en Andrómeda fueron las familias Malvaceae y Meliaceae (cuadro 2).
El área basal es el indicador más heterogéneo del índice de ocupación de espacio. En Los Llanos se encuentra concentrada en solo tres especies (Ficus sp.,Guazuma ulmifoliayEnterolobium cyclocarpum). Por este motivo, no se identifican estratos definidos, siendo elEnterolobium cyclocarpumla especie dominante por sus características fisiológicas (i.e. especies de fácil adaptación, persistencia en áreas altamente degradadas, especies de rápido crecimiento y fijadoras de nitrógeno radical). Mientras que en Andrómeda, hay mayor cantidad de estratos de crecimiento basal, lo que se refleja en una mayor homogeneidad (cuadro 2).
La estructura (ordenamiento horizontal) según los parámetros de abundancia, dominancia y frecuencia, de las especies florísticas identificadas para Los Llanos sobresalen en cuanto a dominancia dos especies:Enterolobium cyclorarpumyFicussp. En Andrómeda se destaca la presencia de más individuos y sobresaleCedrela odorata(Cuadro 3).
En relación con el IVI para Los Llanos elEnterelobium cyclocarpumes la especie de mayor importancia seguida de laBursera simarubay para Andrómeda es laGuazuma ulmifoliayAnacardium excelsum. Dentro de las 10 principales especies según su IVI, cinco de ellas están presentes en ambos sitios con diferente valor de importancia (Cuadro 3).
En cuanto a las familias mejor representadas, la Fabaceae (Caesalpiniaceae, Mimosaceae y Papilionaceae) está presente en mayor cantidad en Los Llanos, en contraposición, la familia Melastomataceae se encontró solamente en el sitio más conservado. La Fabaceae es la familia con el IVF mayor representado en Los Llanos y la Malvaceae está mayormente en Andrómeda. En cuanto a cantidad de familias e individuos, es importante visualizar que, de las 10 principales familias, siete comparten presencia en ambos sitios, lo que muestra una homogeneidad en ambos lugares (Cuadro 4).
CUADRO 2: Área basal (m2/ha) de las especies y familias en parcelas de 500m2, establecidas en Los Llanos y Andrómeda, 2015-2016
| Los Llanos | Andrómeda | Los Llanos | Andrómeda | |||||
| Especies | Área basal | Especies | Área basal | Especies | Área basal | Especies | Área basal | |
| Enterolobium cyclocarpum | 18,62 | Cedrela odorata | 4,08 | Fabaceae Mimosoidea | 18,7 | Malvaceae | 6,5 | |
| Ficus | 14,71 | Pseudobombax septenatum | 3,49 | Moraceae | 15,5 | Meliaceae | 5,1 | |
| Guazuma ulmifolia | 10,41 | Guazuma ulmifolia | 2,96 | Malvaceae | 10,4 | Fabaceae | 2,9 | |
| Tabebuia rosea | 9,87 | Enterolobium cyclocarpum | 2,63 | Bignoniaceae | 9,9 | Anacardeaceae | 2,7 | |
| Bursera simaruba | 6,25 | Anacardium excelsum | 2,63 | Burseraceae | 6,2 | Rubiaceae | 1,8 | |
| Cedrela odorata | 4,95 | Calycophyllum candidissimum | 1,71 | Meliaeae | 4,9 | Euphorbiaceae | 0,9 | |
| Andira inermis | 4,01 | Sapium glandulosum | 0,94 | Fabaceae Papilionoidea | 4 | Sapindaceae | 0,9 | |
| Erythrina berteroana | 3,42 | Trichilia americana | 0,88 | Fabaceae | 3,4 | Moraceae | 0,8 | |
| Anacardium excelsum | 1,79 | Thouinidium decandrum | 0,55 | Anacardeaceae | 1,8 | Polygonaceae | 0,3 | |
| Maclura tinctoria | 0,81 | Chlorophoratinctoria | 0,52 | Urticaceae | 0,8 | Burseraceae | 0,3 | |
CUADRO 3: Índices de valor de importancia de las especies (DAP≥10cm) principales en Andrómeda y Los Llanos, 2015-2016
| Los Llanos | Andrómeda | ||||||||||
| Especies | Número de ind. | Den. Rel. % | Frec. Rel. % | Dom. Rel % | I V I | Especies | Número de ind. | Den. Rel. % | Frec. Rel. % | Dom. Rel % | I V I |
| Enterolobium cyclocarpum | 16 | 21,05 | 15,69 | 24,31 | 61,05 | Guazuma ulmifolia | 36 | 11,84 | 7,65 | 12,00 | 31,49 |
| Bursera simaruba | 12 | 15,79 | 13,73 | 8,16 | 37,67 | Anacardium excelsum | 37 | 12,17 | 8,24 | 10,64 | 31,04 |
| Guazuma ulmifolia | 8 | 10,53 | 9,80 | 13,60 | 33,93 | Cedrela odorata | 16 | 5,26 | 5,29 | 16,53 | 27,09 |
| Tabebuia rosea | 7 | 9,21 | 11,76 | 12,88 | 33,86 | Calycophyllum candidissimum | 21 | 6,91 | 4,12 | 6,92 | 17,95 |
| Andira inermis | 7 | 9,21 | 5,88 | 5,23 | 20,33 | Pseudobombax septenatum | 3 | 0,99 | 1,76 | 14,14 | 16,89 |
| Cedrela odorata | 4 | 5,26 | 5,88 | 6,46 | 17,60 | Enterolobium cyclocarpum | 5 | 1,64 | 1,18 | 10,65 | 13,47 |
| Erythrina berteroana | 3 | 3,95 | 3,92 | 4,46 | 12,33 | Sapium glandulosum | 14 | 4,61 | 4,71 | 3,82 | 13,14 |
| Sapium glandulosum | 3 | 3,95 | 5,88 | 0,23 | 10,06 | Trichilia americana | 16 | 5,26 | 2,94 | 3,55 | 11,75 |
| Maclura tinctoria | 3 | 3,95 | 3,92 | 1,06 | 8,93 | Chlorophora tinctoria | 12 | 3,95 | 4,12 | 2,11 | 10,17 |
| Ficus sp | 2 | 2,63 | 3,92 | 19,21 | 25,76 | Trichilia martiana | 11 | 3,62 | 4,71 | 0,77 | 9,09 |
CUADRO 4: Índices de valor de importancia familiar de las especies leñosas principales (DAP≥10cm) Los Llanos y Andrómeda, 2015-2016
| Los Llanos | Andrómeda | ||||||||
| Familias | Número ind. | Den Rel.% | Dom. Rel % | I V F | Familias | Número ind. | Den Rel.% | Dom. Rel % | I V F |
| Fabaceae Mimosoidea | 17 | 22,37 | 24,35 | 57,83 | Malvaceae | 41 | 13,49 | 26,44 | 47,77 |
| Moraceae | 5 | 6,58 | 20,27 | 37,96 | Meliaceae | 43 | 14,14 | 20,97 | 40,99 |
| Malvaceae | 8 | 10,53 | 13,60 | 29,68 | Anacardiaceae | 46 | 15,13 | 11,14 | 30,20 |
| Burseraceae | 12 | 15,79 | 8,16 | 29,50 | Rubiaceae | 30 | 9,87 | 7,49 | 29,12 |
| Bignoniaceae | 7 | 9,21 | 12,88 | 27,65 | Fabaceae | 14 | 4,61 | 11,73 | 26,14 |
| Fabaceae Papilionoidea | 7 | 9,21 | 5,23 | 20,00 | Sapindaceae | 23 | 7,57 | 3,69 | 21,05 |
| Meliaeae | 4 | 5,26 | 6,46 | 17,28 | Moraceae | 28 | 9,21 | 3,30 | 20,35 |
| Fabaceae | 3 | 3,95 | 4,46 | 13,97 | Euphorbiaceae | 14 | 4,61 | 3,85 | 10,41 |
| Anacardeaceae | 2 | 2,63 | 2,34 | 10,53 | Burseraceae | 11 | 3,62 | 1,11 | 6,68 |
| Euphorbiaceae | 3 | 3,95 | 0,23 | 9,73 | Polygonaceae | 3 | 0,99 | 1,23 | 6,14 |
Con la aplicación de los índices de biodiversidad, se generan datos donde muestra una diversidad similar en los dos sitios de estudio. De acuerdo con el índice de Shannon (H’) y Simpson (Dsi), existe una diversidad media entre los dos; en cuanto a la equitatividad (Pielou), presenta una abundancia homogénea y los índices de similitud Bray-Curtis y Jaccard determinaron una composición similar en cuanto a especies. Entre ambos sitios se comparten 15 familias y 9 especies (Cuadro 5).
Las estimaciones no paramétricas en ambos sitios de estudio muestran que las especies únicas (presentes en una o dos parcelas) tuvieron un porcentaje de 39% y de lassingletons(especies con un solo individuo) el porcentaje fue de 27% (Andrómeda) y 28% (Los Llanos) (Cuadro 6).
La curva de acumulación de especies mostró que mantienen un patrón ascendente y no presenta una asíntota. Se observa una sobre posición de las curvas en Los Llanos, lo que significa una distribución más aleatoria de las especies. En Andrómeda, se muestra un distanciamiento muy leve al inicio, lo que podría evidenciar una agrupación de especies raras agrupadas en pocas muestras, finalizando con una sobre posición de la misma. La curva de Coleman se encontró por arriba de la curva de acumulación de especies, indicando que las especies relativamente raras aparecieron en pocas muestras y que están agregadas (Fig. 4a & 4b).
De acuerdo con los estimadores no paramétricos, la curva de acumulación de especies en ambos sitios es muy disímil, en el sitio más alterado la curva que tuvo un crecimiento inicial alto y una asíntota definida fue ICE (Fig. 5 & 6). La curva singletons tendió a declinar en Los Llanos, mientras tanto, en Andrómeda alcanza su asíntota; la doubletons no se sobrepone en ninguno de los dos sitios (Fig. 6 & 7).
En Los Llanos, el intervalo de riqueza total estimada fue de 20-28 especies; mientras que, en Andrómeda, dicho intervalo fue de 59 a 111 (Fig. 5 & 6). Cuatro estimadores (Chao2 y Bootstrap; Jack1 y ACE) en Los Llanos coinciden en un valor de riqueza de 21 y 23 especies para todas las especies agrupadas (Cuadro 5).
CUADRO 5: Comparación de estimadores de índices de diversidad donde se muestra como resultado la composición de especies entre diferentes comunidades, 2015-2017
| Índice | Estimaciones | |||
| - | Los Llanos | Andrómeda | ||
| Diversidad Shannon-Wiener (H´) | 2,501 | 3,322 | ||
| Dominancia de Simpson (DSi) | 0,891 | 0,947 | ||
| Equitatividad Pielou (J´) | 0,865 | 0,853 | ||
| Similitud/ disimilitud de Bray-Curtis | 0,81 | |||
| Similitud/disimilitud Jaccard | 0,89 | - | ||
CUADRO 6: Riqueza observada y esperada en cada uno de los sitios de estudios según estimadores no paramétricos, 2017
| Sitio más alterado Los Llanos | Sitio más conservado Andrómeda | |
|---|---|---|
| CantCantidad de parcelas | 20 | 20 |
| Especies observadas (Sobservadas) | 18 | 51 |
| EspeEspecies únicas | 4 | 9 |
| Singletons | 5 | 14 |
| ACE | 23±6,71 | 72±12,21 |
| ICE | 28±5,22 | 111±17,49 |
| Chao 1 | 20±5,31 | 60±12,26 |
| Chao 2 | 21±5,03 | 71±15,85 |
| Jackknife 1 | 23±5,95 | 70±17,06 |
| Jackknife 2 | 25±6,85 | 81±21,41 |
| Bootstrap | 21±5,19 | 59±14,39 |
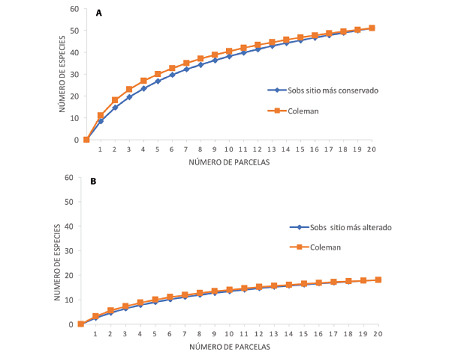
Fig. 4 Curvas de Coleman para explorar el nivel de agregación de las especies de los sitios de estudio, (a) sitio más conservado (Andrómeda) y (b) sitio más alterado alterado (Los Llanos), Alajuela, 2017.
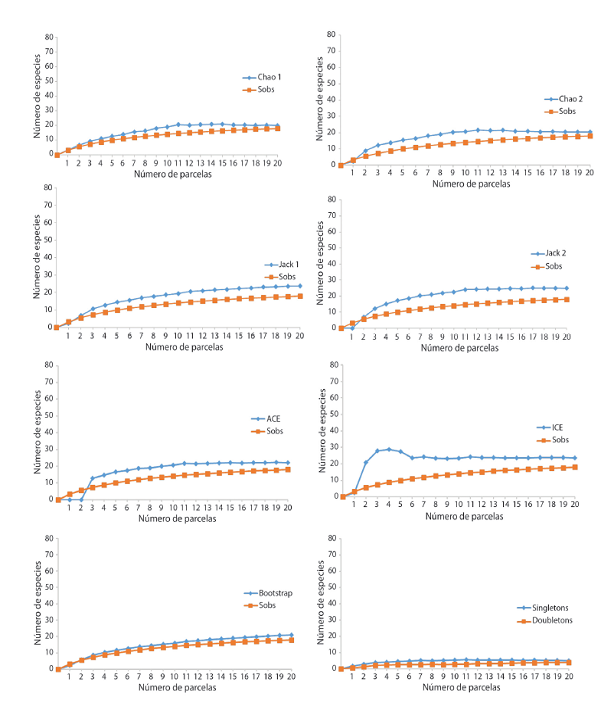
Fig. 5 Comparación de curvas de acumulación de especies de la riqueza observada y la riqueza estimada por siete estimadores no paramétricos, así comosingletonydoubletonspara plantas leñosas con un DAP≥10cm, para Los Llanos, Alajuela, 2017. Nota: Sobs = Observaciones.
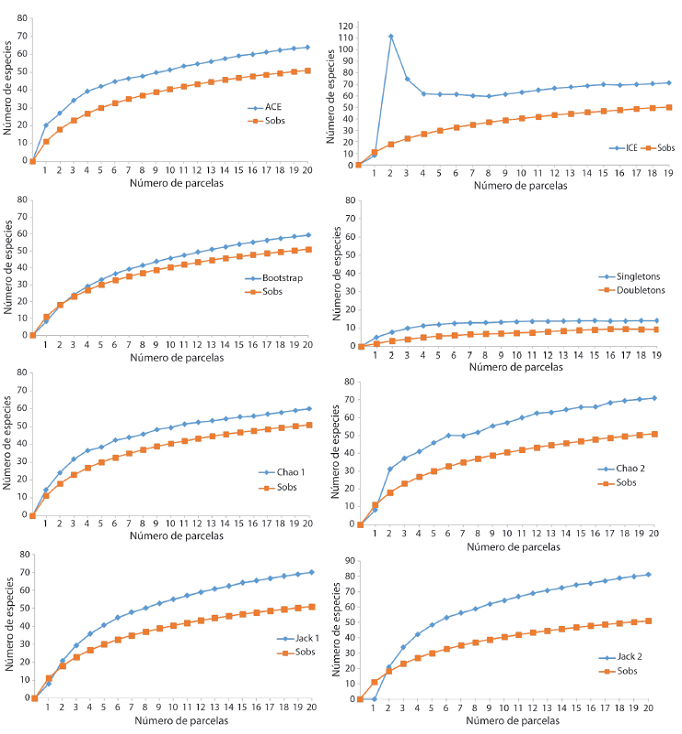
DISCUSIÓN
En diversos estudios sobre la estructura y composición de la vegetación entre dos sitios, se han reportado similitudes en cuanto a su diversidad florística, a pesar de ser sitios muy diferentes (Quisber & Macía, 2005). Igualmente, en la presente investigación, los índices de biodiversidad aplicados dieron como resultado una similitud entre ambos, a pesar de la condición de conservación de una zona y la degradación de la otra (Olivares & Ramos, 2013).
La riqueza de Andrómeda fue de 51 especies de árboles en un área de muestreo de una hectárea, lo cual está dentro del ámbito de 44 a 78 especies por hectárea que estableció Fournier Flores y Rivera (1985) para el Valle Central de Costa Rica (Hernández-Ramírez & García-Méndez, 2015). Al mismo tiempo Hernández y García (2015); mencionan que estos bosques presentan riquezas que varían entre 22 y 105 especies. Los datos obtenidos en el sitio más conservado indican que hay un buen número de especies representadas, autores comoWattenberg, Breckle y Ortizet (1996), Di Stefano, Brenes y Mora (1995) y Cascante y Estrada (2012) refuerzan este dato en sus investigaciones. Asimismo, Vargas (2012); menciona que los bosques humeros premontanos tiene una alta capacidad de resiliencia y pueden recuperarse rápidamente después de un disturbio.
Por otro lado, la composición florística de Los Llanos, presenta una baja riqueza de especies (18 spp), lo que se explica por el uso de suelo durante 32 años (desde 1981), (deforestación, agricultura y ganadería), a partir del 2013 se da una regeneración pasiva.
La composición florística del bosque húmedo premontano transición seca estudiada se asemeja en cuanto a riqueza de especies del bosque seco como por ejemplo en los bosques tropicales secos de la región Pacífico Ecuatorial (Espinosa, Cruz, Luzuriaga, & Escudero, 2012) y en el Parque Nacional Santa Rosa (Guanacaste) se determinaron 56 especies aproximadamente según (Burnham, 1997).
Cascante y Estrada (2012) reportan para el Valle Central 69 especies en en bosque húmedo premontano (bh-P) y para el bosque premontano muy húmedo una riqueza 92 spp para la reserva Madre Verde, Palmares, Costa Rica (Rodríguez & Brenes, 2009). También se reportan 94 spp en la reserva biológica Alberto Manuel Brenes, Wattengerg et al. (1996) y en la zona protectora el Rodeo, se determinó una riqueza promedio de 70 especies por hectárea (Cascante & Estrada, 2012).
A nivel general con respecto a la riqueza de especies reportadas en diferentes investigaciones podemos analizar que la cantidad de especies encontrada es intermedia comparada con bosques más secos y los más húmedos en Costa Rica.
Por otra parte, las especiesEnterolobium cyclocarpum, Guazuma ulmifolia, Cedrela odorataySapium glandusolumse encuentran en ambos sitios como especies con mayor IVI, sin embargo, no necesariamente presentan el mismo orden de importancia (Aguirre, 2013). Menciona Stokes (2001) que estas especies se encuentran, por lo general, en fincas agroforestales, debido a que proporcionan follaje y frutos para alimentar el ganado y proporcionar medicinas y alimentos a los propios ganaderos y a la fauna silvestre.
Asimismo, las familias Fabaceae, Moraceae, Malvaceae, Burseraceae, Meliaeae, Anacardiaceae y Euphorbiaceae presentan mayor IVF en ambos sitios, a pesar de no encontrarse en el mismo orden de importancia, todas son representativas de la zona de vida bh-P transición seca (Fournier et al., 1985). La abundancia de las familias Fabaceae y Malvaceae sugiere que Los Llanos presenta condiciones para que especies como Enterolobium cyclocarpum y Guazuma ulmifoliasean consideradas especies dominantes, por su adaptabilidad a condiciones de suelo perturbados, en presencia de contenidos de arcillas y corresponden a especies de rápido crecimiento (Sánchez et al., 2005; Aguirre, 2013).
Las especies con mayor abundancia en Los Llanos (i.e.Enterolobium cyclocarpum, Guazuma ulmifolia y Bursera simaruba) son típicas del Valle Central (Fournier et al., 1985). Estas se caracterizan por ser de fácil dispersión (e.g. las dos primeras especies son dispersadas por zoocoría o hidrocoría, mientras que la Bursera simaruba por ornitoquiropterocoría), lo cual permite una pronta y fácil colonización de sitios que hayan sido abandonados (CONABIO s.f.; UICN s.f.; CATIE s.f.). Con respecto a estas especies, estudios demuestran que, en general, las fincas ganaderas de América Latina presentan una alta abundancia de estas especies arbóreas, aunque muchas se encuentran en bajas densidades (Guevara, Laborde, & Sánchez; 1998; Harvey & Haber, 1999; Otero-Arnaiz, Castillo, Meave, & Ibarra-Manríiquez, 1999; Cajas, & Sinclair, 2001).
En este mismo sitio de estudio, se identificaron especies con DAP≥80cm, lo que puede estar asociado a menos presión en cuanto a competencia por nutrientes, agua y espacio (Aguirre, 2013). Otros estudios han encontrado en sus investigaciones árboles grandes con un DAP≥100cm (Esquivel, Ibrahim, Harvey, Villanueva, Benjamín, & Sinclair, 2003). Por otro lado, Donoso (1981) describe que el espacio de crecimiento de las especies depende de las características bióticas y abióticas, además, que existe un conjunto de factores que inciden en el crecimiento y se refleja en la densidad.
Ahora bien, otro aspecto importante en cuanto a la composición y estructura es la dominancia de las especies, se identificó que, de las tres especies representativas de Andrómeda solamenteGuazuma ulmifoliapresenta dominancia en Los Llanos. Esta especie se caracteriza por ser de fácil adaptación a suelos vertisoles, crecen en suelos perturbados, son de rápido crecimiento, son recuperadores de terrenos degradados, comunes en áreas húmedas y secas, proporcionan sombra y refugio a fauna (CONABIO, s.f.), son especies importantes para procesos de restauración ecológica.
Los resultados de los índices de biodiversidad aplicados en los dos sitios de estudio nos indican en términos generales que la composición de las especies es similar, o sea podemos encontrar las mismas especies ambos lugares (se comparten 15 familias y nueve especies). Con la generación de estos datos podemos tomar como base las especies identificadas para realizar procesos de restauración ecológica en el sitio más degradado (Cabrera-Condarco, 2005; Quisbert & Macía, 2005; Dzib, Chanatásig, & González, 2014).
La aplicación de los índices de diversidad se utiliza para hacer comparaciones en donde no se conoce el número total de especies y para áreas más grandes (Ponce, Mehltreter, & Elías, 2002). Además, la obtención de estos índices da pie para conocer el estado actual de ambos lugares y sus datos sirven para monitorear y darle seguimiento a los procesos de restauración (González, Avella, & Díaz, 2015)
En cuanto a la distribución espacial, la curva de acumulación de especies presenta una diferencia entre la curva Coleman y la de todos los individuos observados. Muestra relativamente una mayor sobreposición, lo que señala una distribución más aleatoria de las especies para ambos sitios (López & Williams, 2006).
En Andrómeda, se mostró que en la curva de acumulación de especies se da un distanciamiento al inicio y finaliza con una sobreposición, lo que significa que al inicio hay una agregación de especies y termina con una distribución más azarosa. A diferencia de Los Llanos, donde ambas curvas están sobrepuestas, significando más aleatoriedad de las especies. Esta diferencia se marca por la abundancia de especies en cada sitio de estudio.
Por otro lado, las curvas de acumulación de ambos sitios no muestran su asíntota definida, lo que podría significar que se requiere aumentar el esfuerzo de muestreo, sin embargo, Romero-Saltos, Valencia y Macía (2001) yDuivenvoorden et al. (2001) mencionan que una mayor intensidad de muestreo no asegura la estabilidad de la curva especies-área, sugiere que la diversidad local del área es mayor a la encontrada. En la evaluación de los estimadores no paramétricos, se compara el comportamiento de la curva de acumulación de especies de Chao 1 y 2, ACE, ICE, Jackknife 1 y 2 (Chazdon, Colwell, Denslow, & Guariguata, 1998), con el fin de estimar el número de especies esperadas para ambos sitios de estudio (Palacios, Mendoza, Lozano, & Yaguana, 2017).
Con respecto al comportamiento general de los estimadores no paramétricos, se puede observar que, para Los Llanos, se muestra un distanciamiento menor entre sus curvas, lo que evidencia más aleatoriedad de las especies. En Andrómeda, se observa un distanciamiento mayor, lo que significa una agrupación de las especies. Se considera que, si aumenta la agregación, también aumenta la presencia de las especies en una sola muestra. Esto se podría relacionar también con el hecho de que en Andrómeda hubo mayor porcentaje de especies únicas y una densidad de especies menor que en el sitio más perturbado (López & Williams, 2006).
En Los Llanos, se encontró que el estimador ICE tiene una tasa inicial alta y con asíntota definida. Para Andrómeda ICE y Chao2 muestran ambas características. Los mejores estimadores de riqueza son los que presentan curvas de acumulación con un crecimiento inicial rápido y alcanzan una asíntota definida (Gimaret-Carpentier, Pélissier, Pascal, & Houllier, 1998;Skov & Lawesson, 2000; Longino, Coddington, & Colwell, 2002). De ahí que se da una estimación para el sitio más conservado de 71-111 especies y en el más alterado una estimación de 28 especies.
Debido a la historia de degradación que ha presentado Los Llanos, no es posible conocer las características vegetativas iniciales. Cabe mencionar que la distancia a la cual se encuentra el sitio más conservado es considerable y por ello la accesibilidad a fuentes semilleras es muy limitada.
Para concluir, es importante tener presente que la generación de estos datos contribuyen al conocimiento sobre el ecosistema y proporcionan información base para la implementación de procesos de restauración ecológica (Rodríguez & Brenes, 2009; Cascante & Estrada, 2001, 2012; Di Stefano et al., 1995;Founier, 1998). El bh-P transición seca es el más alterado y reducido en Costa Rica (Fournier, 1985;Hartshorn, 1991), en los últimos años la pérdida de este ha sido más evidente. Este estudio generó información sobre especies presentes y ausentes en la transición de esta zona de vida, proporcionando una base importante para próximas investigaciones.
Por tanto, a raíz de los resultados de la investigación, se da respuesta a la hipótesis planteada: la estructura y composición de la vegetación arborescente difiere entre sitios alterados por deforestación y ganadería y sitios conservados, los diversos índices de biodiversidad aplicados indican que hay una similitud entre ambos sitios en cuanto a composición, pero no son homogéneos y en su estructura es muy diferente, por lo tanto, se acepta la hipótesis ya que ambos sitios difiere uno del otro.
Al mismo tiempo, la estructura florística muestra una diferencia un poco más notoria asociada a la perturbación y conservación respectiva de cada sitio, lo cual ha permitido que para el sitio de control exista una mayor diversidad de especies, presentando una mayor abundancia de individuos, especies, géneros y familias, esto responde al objetivo de la entidad propietaria, a saber, el SINAC. Los Llanos presenta características de uso de suelo histórico producto de la actividad agropecuaria, propiciando la degradación de este ecosistema, presenta en menor cantidad individuos, especies, géneros y familias.

