











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkEn Costa Rica existe una gran cantidad de lagos asociados al vulcanismo (Gocke, 1997; Alvarado & Salani, 2004). La laguna Hule se encuentra localizada en una depresión volcánica ocupada por tres lagos, de los cuales Hule es el más grande ocupando 54,7Ha y una profundidad de 26,5m. La zona de vida presente en la región es el bosque muy húmedo premontano, con una precipitación media anual del área de 4 500-5 500mm (Holdridge, 1987).
En las comunidades de organismos acuáticos, uno de los grupos de mayor importancia son los peces, cuyos mecanismos de supervivencia comprenden adaptaciones morfológicas, fisiológicas, reproductivas y alimentarias, que repercuten directa o indirectamente en los hábitats que frecuentan (Jackson, Peres-Neto & Olden, 2001). La ictiofauna continental neotropical se caracteriza por poseer el 24% de la totalidad de los peces a nivel mundial (Albert & Reis, 2011). Esta riqueza se ve representada en los diferentes ambientes (loticos, lenticos o estuarinos) de la región y en Costa Rica convergen tres de las cuatro provincias ícticas, factor que explica su considerable diversidad (Bussing, 1998; Angulo et al., 2013).
Los peces juegan un papel importante en las dinámicas tróficas de los cuerpos de agua, donde a través de la depredación del zooplancton promueven efectos de cascada trófica (Carpenter & Kitchell, 1996; Frank et al., 2005; Kratina et al., 2012). Además, conectan diferentes zonas mediante la translocación de nutrientes entre hábitats (Vander Zanden & Vadeboncoeur, 2002). Permiten el desarrollo del perifiton y el fitoplancton (Jones & Sayer, 2003, p. 2156) al proveerlos con nutrientes excretados (Scheffer et al., 1993; Vanni, 2002; Tessier & Woodruff, 2002; Ellis et al., 2011).
Una de las formas de describir una comunidad ecológica es determinando su composición de especies y la abundancia relativa de cada una. Los índices de diversidad ayudan a resumir las asociaciones de organismos (Wilhm, 1968; Mason et al., 2005). Los cambios en la composición de especies son indicadores de presiones en el ecosistema y de los impactos ambientales a través del tiempo (Livingston, 1976; Moore, 1978; Smith, 1991; Weibull, Östman & Granqvist, 2003; Gorelick, 2006). Se registran pocos trabajos relacionados con peces en este lago (Tabash, Farid & Guadamuz 2000), de modo que este es el primer estudio descriptivo de la comunidad íctica en la Laguna del Hule.
Materiales y Métodos
La laguna Hule se ubica en una depresión volcánica a 800msnm, en la Vertiente Caribe de la Cordillera Volcánica Central, en el cantón de Río Cuarto, Alajuela, Costa Rica; forma parte del Refugio Nacional de Vida Silvestre Bosque Alegre. La investigación se realizó con el permiso de investigación del SINAC No. 03116. El muestreo se hizo dos veces al mes, en el día y la noche, durante la temporada lluviosa de agosto a diciembre del 2015. El muestreo también incluyó el efluente quebrada Pata de Gallo, que representa el cuerpo de agua más cercano a la laguna.
La abundante cantidad de vegetación acuática condicionó el muestreo a los siguientes artes de pesca: líneas de pesca, redes y chinchorro de diferente luz de malla. Se muestreó en la orilla expuesta y entre la vegetación un tramo total de aproximadamente 800m, mismos que se dividieron en ocho secciones de diez metros cada una, en promedio se muestreaba durante 20min en cada sección. Los peces capturados fueron identificados in situ utilizando la clave taxonómica elaborada por Bussing, (1998), se midió su longitud estándar (LT) usando un ictiómetro (±1mm) o una regla plástica (±0,5mm) cuando su tamaño lo condicionaba y se liberaron en el mismo sitio donde se capturaron. Además, se verificó el sexo de los poecílidos.
El aporte de las especies dominantes se determinó mediante el criterio de Ayala-Pérez et al. (1998), quienes lo establecen como el aporte de más del 80% de la captura total. Para el procesamiento estadístico y la creación de gráficos se utilizó el software libre R (R Development Core Team, 2008) y Excel. Se evaluaron algunas relaciones ecológicas utilizando índices de diversidad y el paquete estadístico “Vegan” del software R. Los índices elegidos para la abundancia relativa fueron los de Pielou, Fischer, Soresen y especies no detectadas; además se generó una curva de acumulación de especies.
Para determinar si las diferencias entre el tamaño de machos y hembras de las especies de poecílidos eran significativas se aplicó la prueba de Welch Two Sample t-test. En caso que los datos no presentaran homogeneidad de varianza, se aplicó un test de Wilcoxon; misma prueba que se utilizó para buscar preferencias en el uso del hábitat de las especies capturadas. Además, se evaluó visualmente el estado ribereño de la laguna en busca acciones antropogénicas que pudieran amenazar la integridad y biodiversidad del sitio, cuantificando las principales problemáticas presentes para su posterior discusión.
Resultados
Seis especies se registraron en las orillas de la Laguna Hule y dos en la quebrada Pata de Gallo, para un total de 223 y 117 individuos, respectivamente (Cuadro 1). Las especies más abundantes en la laguna correspondieron a A.aeneus, P.amates y P.gillii, mientras que en la quebrada fue B. olomina. La frecuencia relativa con la que se encontraron las especies fue de 0,013 para A. alfari, 0,25 para A. aeneus, 0,184 para P. dovii, 0,197 para P. amates y 0,348 para P. gillii.
Cuadro 1 Especies, cantidad de ejemplares capturados, y tallas de peces en la Laguna de Hule y en el efluente Pata de Gallo.
| Especies | Hule | Pata de Gallo | Largo total (cm) | |
|---|---|---|---|---|
| min | max | |||
| Amatitlania siquia* | 2 | 0 | 6 | 9.5 |
| Amphilophus alfari | 3 | 0 | 2.3 | 20 |
| Astyanax aeneus | 40 | 0 | 1.9 | 14 |
| Parachromis dovii | 59 | 0 | 4 | 45.5 |
| Phallichthys amates | 37 | 0 | 2.7 | 6.1 |
| Poecilia gillii | 79 | 0 | 2.5 | 8.3 |
| Brachyrhaphis olomina | 0 | 110 | 5.3 | 8.6 |
| Rhamdia laticauda | 0 | 7 | 8.9 | 14.5 |
| Cynodonichthys sp** | 3 | 0 | - | - |
| Total | 223 | 117 | - | |
*Capturado en una visita a la Laguna Congo.
** Capturado en charcos cerca de la laguna.
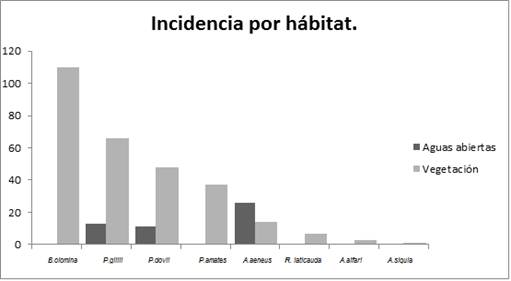
Un evento de relevancia es el primer reporte de la presencia del género Cynodonichtys en el sitio, específicamente en charcas temporales cerca de la laguna, siendo la zona más cercana donde se ha reportada el Río Sarapiquí, a una distancia de 4Km de la laguna. Además, se informa de la presencia de A. siquia en la laguna Congo, que a pesar de la cercanía con la laguna Hule, no fue encontrada en la misma durante todos los meses de muestreo. Relacionado con la elección de hábitat (Fig.1), se determinó que las especies A. alfari, P. amates, P. gillii (t-test (p<0.05)) y P. dovii (Wilcoxon (p>0.05)) muestran una tendencia marcada a vivir resguardadas entre la vegetación acuática o de la orilla, mientras que A. aeneus posee una preferencia hacia aguas abiertas (t-student (p<0.05)).
En las tres especies de poecílidos las hembras son más abundantes que los machos, con frecuencias que varían entre 0.7 y 0.83, siendo la más alta para B. olomina, que se encontró en la quebrada, donde la frecuencia de machos (0.17) es la más baja. De todos los individuos muestreados, se encontró que los de esta quebrada mostraban parásitos dermales en forma de manchas negras, en una frecuencia de 0.16. Mientras que en la laguna Hule se observó que la mayor parte de los juveniles de P. dovii presentaron gran cantidad de quistes dermales.
No hubo diferencia entre el tamaño promedio de los sexos para P. gillii y P. amates (Welch Two Sample, t-test (p>0.05)), caso contrario B. olomina si presentó diferencias (Welch Two Sample t-test (p<0.05)). En general, las hembras de B. olomina y P. amates presentaron mayores tallas, al contrario que con P. gillii (machos de mayor tamaño) (Cuadro 2).
Cuadro 2 Evaluación del promedio de la longitud total (LT) de las especies de poecílidos, según su sexo.
| Especie | LT hembras (cm) | LT machos (cm) |
|---|---|---|
| B. olomina | 6.528 a | 5.278 b |
| P. gilli | 4.302 a | 4.823 a |
| P. amates | 4.208 a | 4.109 a |
Promedios con diferente letra, en la misma fila, presentaron diferencia significativa (p<0.05).
Los índices de diversidad en los dos sitios muestran que esta es baja, siendo la Laguna de Hule la más diversa de las dos; el índice de equidad de Pielou refleja una dominancia bastante repartida en la laguna; mientras que en la quebrada la alta dominancia por parte de una especie es muy notable. Ninguna de las especies encontradas en la quebrada estaba presente en la Laguna de Hule y viceversa, aspecto que se confirma con el índice de disimilitud de Soresen. La curva de acumulación de especies señala que la mayor parte de estas fue hallada en la laguna. Según los índices de especies no detectadas, faltaron de tres a cinco de encontrar en los sistemas de agua dulce muestreados (Cuadro 3). Los problemas de origen antropogénico encontrados se resumen en el Cuadro 4, así como sus riesgos y posibles soluciones.
Cuadro 3 Valores de los índices ecológicos aplicados a las poblaciones de peces capturados en los diferentes sitios muestreados durante un periodo de 5 meses en la laguna Hule
| Índices | Hule | Pata de Gallo |
|---|---|---|
| Shannon | 1,4839 | 0,2265 |
| Pielou | 0,7626 | 0,3268 |
| Fisher | 1,3737 | 0,3427 |
| Soresen | 1 | |
| Acumulación Especies | 6,5636 | 2 |
| Índices de especies no detectadas | ||
| Jack knife | 4,92 | |
| Bootstrap | 2,86 | |
Cuadro 4 Lista de problemáticas encontradas en la laguna de Hule durante los muestreos, riesgos que conllevan y posibles soluciones.
| Problemáticas | Riesgos | Posibles soluciones |
| Contaminación por parte de los visitantes (equipo de pesca abandonado, latas, basura en general) | Los animales corren riesgos de ingestión, quedar atrapados o ser heridos, pérdida de la belleza escénica. | Acondicionar basureros en el sitio, rótulos recordando al visitante sus responsabilidades, crear un grupo de limpieza que visite regularmente el sitio |
| Desconocimiento del estatus de protección de la zona | Se realizan actividades no permitidas en lugares protegidos (pesca, caza, contaminación) | Creación y colocación de rótulos aclarando el estatus de protección de la zona. |
| Introducción de especies invasivas | Desbalance en la cadena trófica que puede poner en riesgo la supervivencia de varias especies y comprometer la estabilidad del ecosistema | Impartir talleres en la zona advirtiendo los riesgos que implica introducir especies en el lago |
| Pesca no controlada y utilización de artes de pesca destructivos | Disminución de las poblaciones de las especies afectadas | Colocar rótulos donde explícitamente se indique la ilegalidad de la actividad y las medidas legales (multas u otras); crear un grupo comunal que vigile la zona varias veces a la semana |
Discusión
La baja diversidad de especies encontrada en los sitios de muestreo manifiesta que los hábitats estudiados no poseen la suficiente heterogeneidad, por lo que no proveen una mayor disponibilidad de micro-hábitats que permitan una mayor riqueza y colonización de especies (Walter & Haynes, 2006; Matthews, 2012). La heterogeneidad estructural del hábitat moldea la estructura de la comunidad de peces tropicales, influyendo en la distribución de especies y en los patrones de segregación a diversas escalas temporales (Ault & Johnson, 1998; Palmer, Menninger & Bernhardt, 2010). Las especies encontradas son propias de aguas mansas y con poca corriente, presentan una preferencia por ambientes profundos y anchos, con una menor velocidad de agua (Bart, 1989; Matthews, 2012). Los hábitats profundos proporcionan mayor estabilidad que los que presentan rápidos, debido a que atenúan cambios en la velocidad de corriente, profundidad y diferencias en el sustrato; lo cual pudo influenciar las diferencias entre la laguna y la quebrada (Schlosser, 1982; Beeton, 2002).
A diferencia de quebradas y ríos, donde las especies base en la cadena trófica, como A. aeneus, tienden a refugiarse en rápidos para evitar ser depredadas; en las lagunas, estas se distribuyen en aguas abiertas. Lo anterior disminuye la probabilidad de encontrarse con depredadores acuáticos que acechan en la maleza, como el P. dovii (Bussing, 1998; Mehner et al., 2005). Los peces de la familia Characidae son capaces de habitar regiones con mayor elevación y flujo de caudal, permitiéndoles un mayor ámbito de distribución en los cuerpos de agua dulce (Bussing & López-Sánchez, 1977; Azevedo, Melo & Young, 2011; Kasumyan & Marusov, 2015). A. aeneus además es muy importante en la red trófica, dado su papel en el ecosistema como uno de los principales recicladores de nutrientes (nitrógeno y fósforo), en hábitats donde hay poco aporte de los mismos (Small et al., 2011).
Los poecílidos presentes siguen la tendencia de esta familia de peces, predominando las hembras en número y tamaño (Chapman et al., 1991; Jennions & Kelly, 2002). El hábitat de preferencia es consistente con el reportado (Bussing, 1998), P. amates solo se encontró en la vegetación acuática; P. gillii es más adaptable, pudiendo vivir en aguas rápidas con caudales moderados (Burcham, 1988; Lorion, Kennedy & Braatne, 2011). A pesar de que se reporta un mayor tamaño para hembras de P. gillii (Bussing, 1998), estas observaciones muestran que algunos machos sobrepasan el tamaño máximo de las hembras. Respecto a las demás especies, sus tamaños máximos y mínimos están dentro de los rangos previamente reportados (Angulo et al., 2013). Para P. dovii no se contaron con muchos datos, debido a la imposibilidad de muestrear lejos de la orilla (ausencia de botes y sus respectivos permisos para utilizarlos).
Un hallazgo importante para el Refugio Bosque Alegre es el primer reporte de la presencia de Cynodonichthys, cuyo hábitat tipo consiste en charcos lodosos, pantanos, acequias y aguas de poca a moderada velocidad de corriente; coincidiendo con el tipo de hábitat donde fue encontrado en este trabajo (charcos lodosos de origen volcánico de poca profundidad cercanos a la laguna) (Bussing 1998).
Se encontró que la población de B. olomina poseía longitudes totales que superan las reportadas por Bussing (1998) (60mm frente a 75mm); diferencia que puede atribuirse a la escasez de depredadores. Aunque las poblaciones de Brachyrhaphis ocupen distintos hábitats, las diferencias ecológicas más pronunciadas entre ellas son causadas por la presencia o ausencia de peces depredadores. A excepción de R. laticauda, no se encontró ningún pez en la quebrada que pudiese alimentarse de esta especie, por lo que la mortalidad debe estar asociada a otros factores ecológicos o depredadores terrestres (Johnson, 2002). Se observó la presencia de parásitos dérmicos en un 25% de la población de B. olomina, que pudiesen ser los encargados de controlar en parte el tamaño de su población.
La Laguna de Hule presenta diversas problemáticas ambientales. La falta de vigilancia y estricta demarcación de la zona son las causas principales de la pesca y contaminación. Este es un tema complicado ya que la naturaleza mixta público-privado del refugio no permite el suficiente control sobre los asuntos relacionados a su manejo. En los múltiples muestreos realizados se encontró una gran cantidad de basura dispersa dentro de la laguna y en sus alrededores, patrón de comportamiento común en varios lagos (Corona-Lisboa, 2013). Se propone la colocación de rótulos, que delimiten el estatus de protección de la zona. Dichas señalizaciones deberían distribuirse a lo largo del camino, en distintas ubicaciones, para evitar que sean robados o removidos. Además, se propone la creación de un grupo comunal que vigile la laguna varias veces a la semana.
Se encontraron personas realizando actividades de pesca durante todos los muestreos. La pesca del P. dovii se lleva a cabo sin regulaciones. Tabash, Farid & Guadamuz (2000) mostraron que un incremento en la pesca deportiva de esta especie en la laguna podría afectar la capacidad de recuperación de la población. Esto puede propiciar fluctuaciones peligrosas en las poblaciones, ocasionadas por el descenso del número de individuos maduros y la consecuente baja en su capacidad reproductiva (Murawski, 2000; Jackson et al., 2001; Allan et al., 2005). Esta pesca casi siempre es acompañada de la extracción de sardinas (A. aeneus) para utilizarlas como carnada, con el consecuente efecto sobre esta especie.
Los habitantes constataron la liberación de especies invasoras en este cuerpo de agua. Puntualmente se liberaron especies como el machín (Agonostomus montícola) y sábalo (Brycon guatemalensis), lo cual no se pudo corroborar en los muestreos. Esta situación podría llevar a un desbalance en la cadena trófica, poniendo en riesgo la supervivencia de varias especies (Pimentel, Zuñiga & Morrison, 2005). Se propone impartir talleres en la zona, advirtiendo los riesgos que implica introducir especies en la laguna (Benayas, Gutiérrez, & Hernández, 2003).

