











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkFrom 1979 to 1983 ecological surveys were conducted in the Gulf of Nicoya estuary, Pacific coast of Costa Rica to provide baseline information in support of government management strategies (see references in Vargas 1995, and Vargas& Mata, 2004). These studies were followed by surveys (1984-1987 and 1994-1998) of an intertidal flat in the upper estuary. The results of the first survey were previously published by Vargas (1987, 1988a,b, 1989, 1996) and the structure of this benthic community was discussed.
More recently we have focused on the abundances of the main taxonomic groups found at the intertidal flat, including cephalochordates (Vargas& Dean, 2010), echinoderms (Vargas& Solano, 2011), mollusks (VargasZamora & Sibaja-Cordero, 2011), and crustaceans (Vargas-Zamora, Sibaja-Cordero &Vargas-Castillo (2012). The availability of user-friendly software and expanded computer memory facilitates the analyses of these data sets to find previously hidden patterns, or to confirm previously described patterns, as illustrated early by Stephenson, Williams & Cook (1970). One set of programs to analyze temporal patterns is the Generalized Additive Models (GAM) which has been applied to temperate infaunal communities by Stoner, Manderson, & Pessutti, (2001) and Jennings, Nicholson, Dinmore & Lancaster, (2002) among others. The review of the methods by Wood (2006) and by Wood, Goude & Shaw (2015) have facilitated its use. At tropical latitudes we used GAM to describe temporal oscillations of the molluscan and crustacean species from the Punta Morales flat (vargasZamora & Sibaja-Cordero, 2011; Vargas-Zamora, SibajaCordero &Vargas-Castillo, 2012). Dean (2009) updated the list of polychate worms reported from Costa Rica. Polychaeta was the most important taxonomic group in terms of number of species at the Punta Morales flat (Vargas, 1987, 1988a, 1889). Thus, a closer look of its population patterns was in order.Therefore, the objective of this note is to make accesible an updated list of the polychaete species found at the flat, and the results of the application of GAM and multivariate statistical methods to the updated polychaete data set.
Materials and methods
Study site: The intertidal flat is located on the South shore of the Punta Morales peninsula on the mid-upper region of the Gulf of Nicoya estuary (10oN-85oW) Pacific coast of Costa Rica. The sand-mud flat is limited to the North by a white sand beach. Rocky outcrops and mangrove stands are also prominent features of the peninsula (Figure 1). A dry season (December to April) and a rainy season (May to November) mark the seasonality of nutrient dynamics and water salinity, which have been described by Epifanio, Maurer & Dittel (1983) and Voorhis, Epifanio, Maurer, Dittel &Vargas (1983) as part of the ecological surveys mentioned before. Freshwater inputs from seasonal rains and runoff carried by the Tempisque and Tarcoles rivers (Figure 1) produce strong vertical and horizontal salinity gradients that drive estuarine circulation. Near Punta Morales the Largarto river (Figure 1) contributes suspended sediment loads during the rainy season.
Field and laboratory methods: The sampling protocols are outlined in Vargas (1987, 1988a, 1989) and in Vargas-Zamora & Sibaja-Cordero (2011).In summary 14 cores (17.7 cm2- 15 cm long) were collected at monthly intervals from muddy sands during low tide (mean tidal range: 3 m), from a 400 m2 plot located 20 m from a sandy beach (Figure 1). The original data set of Vargas (1987, 1988a, 1989) was expanded with additional data from MarchApril, 1987, for a total of 49 dates (February, 1984 to April, 1987) and with data collected at the site by Solano-Ulate (2007) from July 1994 to September 1996 (28 dates) using the same methodology. This sampling effort (77 dates) is unique in tropical intertidal soft-sediment studies. At the laboratory core samples were fixed in Rose
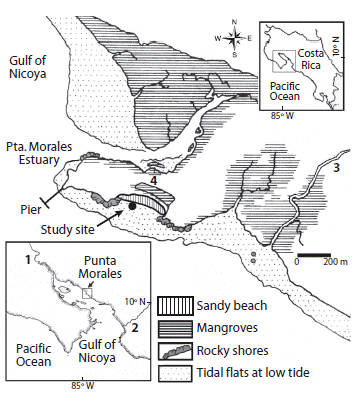
Figure 1: Location of the study site at the Punta Morales peninsula, Gulf of Nicoya estuary (10o 04'N-84o 58'W), Pacific coast of Costa Rica. 1: Tempisque river. 2: Tárcoles river. 3: Lagarto river. 4. Marine field station.
Bengal stained formalin in sea water and later sieved (500 micron mesh). Organisms were sorted and stored in ethanol filled shell vials. The original list (Vargas, 1986) of polychaete species was updated based on Dean (2009) and by inspection of the collections of voucher specimens deposited at the Museum of Zoology (University of Costa Rica) and at the Museum of Comparative Zoology (Harvard University). The presently valid names of species correctly identified in Vargas (1986), as well as those assigned to new taxa, were verified by accesing the web page w.w.w.WORMS (World Register of Marine Species).
Statistical methods: A Principal Component Analysis (PCA) was conducted on the log10 (x+1) transformed abundance data of the 43 polychaete taxa (1984-1987). Additionally, a two-way ANOSIM (Clarke & Warwick, 1994) of the same data based in the Euclidean distance, was done to test seasonal effects (as first factor) on the assemblage of polychaetes. As a second factor the data series was evaluated for changes before and after the red tide events that were frequent in the upper Gulf of Nicoya in 1985 and reported by viquez & Hargraves (1995)
Temporal trends were analyzed for the two more abundant species found during all dates of the 19841987 and 1996-1998 surveys (the capitellid Mediomastuscaliforniensis and the spionid Paraprionospio alata) with the Generalized Additive Model (GAM) of the free mgcv package in The R Project for Statistical Computing, with abundances log10 (x+1) transformed to homogenize variances (Stoner, Manderson, & Pessutti, 2001). The GAM was carried out with the subroutine quasi (Wood, 2006). The mean abundances per season (dry vs rainy, 95% confidence limits) of these species were computed also with log10 (x+1) transformed data and back to the original scale for graphical display.
Results
Information on environmental data is included in Vargas (1987, 1988, 1989, 1996), Vargas& Solano (2011), and Vargas-Zamora & Sibaja-Cordero (2011). In summary, sediment composition of the sand-mud flat averaged 65% sand and 32% silt+clay. Seasonal trends in sediment and water temperatures were not found by Vargas (1987, 1988a). Water temperatures above 30oC were characteristic of this region of the estuary, with a maximum of 40oC on April 1984 (Vargas 1987). Water salinities during the survey ranged from as low as 22 ppt (rainy season of 1984) to 34 ppt (dry seasons). The reader is referred to the original work of Solano-Ulate (2007) for more information on the 1994-1998 macrofaunal survey.
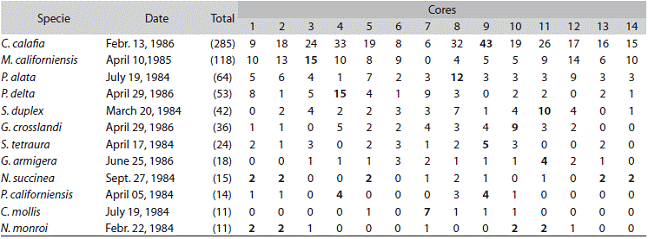
A total of 43 polychaete species were found at the site of which 15 species names (35%) were updated and eight taxa await further taxonomic work (Table 1). During the 1984-1987 survey the 43 species were represented by a total of 6 600 individuals collected in a total sample area of 1.21 m2. The 43 polychaete species were distributed among 25 families (Table 1), with the Spionidae (6), Phyllodocidae (4), and Nereididae (3) as the most speciose families. Six species were represented by a single individual and the following seven comprised 80% (Table 1) of the total number of individuals: the capitellid Mediomastus californiensis (2140 ind., 32.4%), the spionids Caraziella calafia (1338 ind., 20.3%), Paraprionospio alata (606 ind., 9.2%) and Spiophanex duplex (252 ind., 3.8%), the lumbrinerid Scolotema tetraura (394 ind., 5.9%), the nereidid Gymnonereis crosslandi (327 ind., 4.9%), and the goniadid Glycinde armigera (235 ind., 3.5%). The number of species collected per date ranged from 10 to 23, and the number of polychaetes ranged from 47 to 397 (Table 2). The capitellid M. californiensis was the numerically dominant worm for 34 of the 49 dates, followed by the spionids C. calafia (9 dates), P. alata (5) and S.duplex (1). Numerical dominance of M. californiensis ranged from 23 to 74 % (Table 2).
Table 1: Left columns : codes for the 43 polychaete (Annelida: Polychaeta) species as identified by 1987. Codes 01-95 as in Vargas (1986,1987,1988a,1989). Codes 102-122, this study. Original species identifications. Center and Right columns : updated (this study) species identifications, polychaete families and total number of individuals collected. Intertidal sand-mud flat, Gulf of Nicoya estuary, Pacific coast of Costa Rica

Table 2: Date code (1 to 49). Date (1984-1987).Total number of polychaete species (S). Total number of polychaete individuals (N). The species with the maximum number (n) of individuals and its code (as in Table 1). Percentage (%) of N represented by that species. Punta Morales sand-mud flat. Gulf of Nicoya estuary, Pacific. Costa Rica
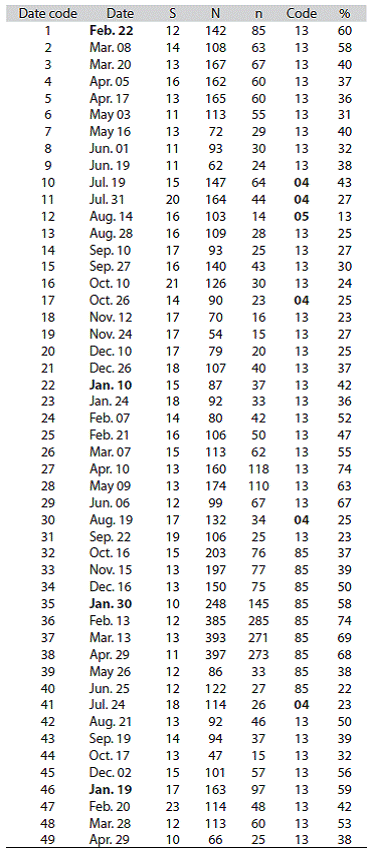
From the 1994-1998 survey we focused here on the population oscillations of P. alata and M. californiensis only. Thus, the abundance fluctuations of these two polychaete wormsduring the 77 sampling dates are included in Figure 2, Figure 3, and Figure 4 (Totals: 1078 cores, 1.90 m2 ).
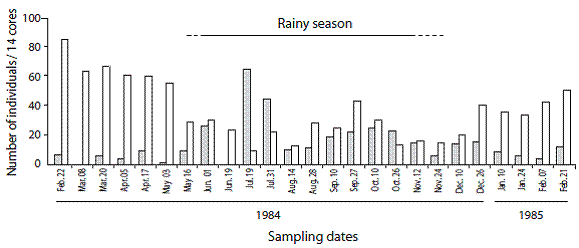
Figure 2: Numbers of Paraprionospio alata (gray bars) and Mediomastus californiensis (white bars). The extent of the rainy season (lower salinities) is indicated. Gulf of Nicoya estuary, Pacific coast of Costa Rica. Punta Morales intertidal sand-mud flat. (February 1984 to February 1985).
Figure 3: Numbers of Paraprionospio alata (gray bars) and Mediomastus californiensis (white bars). The extents of the rainy seasons (lower salinities) are indicated. Gulf of Nicoya estuary, Pacific coast of Costa Rica. Punta Morales intertidal sand-mud flat. (March 1985 to April 1987).
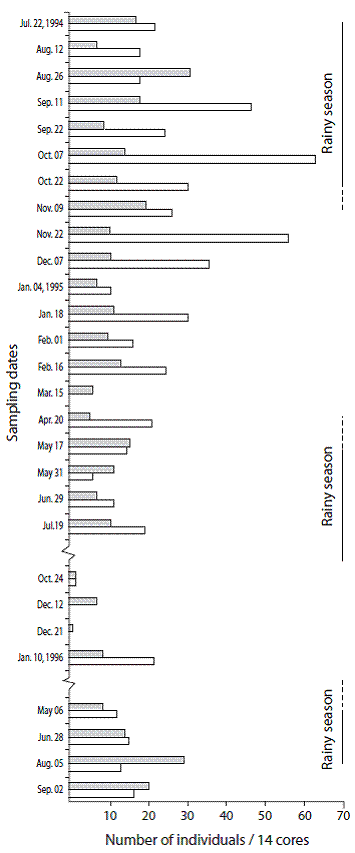
Figure 4: Numbers of Paraprionospio alata (gray bars) and Mediomastus californiensis (white bars). The extents of the rainy seasons (lower salinities) are indicated. Gulf of Nicoya estuary, Pacific coast of Costa Rica. Punta Morales intertidal sand-mud flat. (July 1994 to September 1996).
The abundance fluctuations of nine selected species from different families of worms are illustrated in Figure 5. for the 1984-1987 survey. In general, abundances were patchy in time. However, higher abundances were observed during the first half of 1984 and declined afterwards, with the exception of P. delta which had a peak after the red tides of 1985. The spatial distribution among the 14 cores of these nine species, and those of M. californiensis, C. calafia and P. alata, is illustrated in Table 3 for the date when the highest abundance of each species was found. Patchiness is space was also evident.
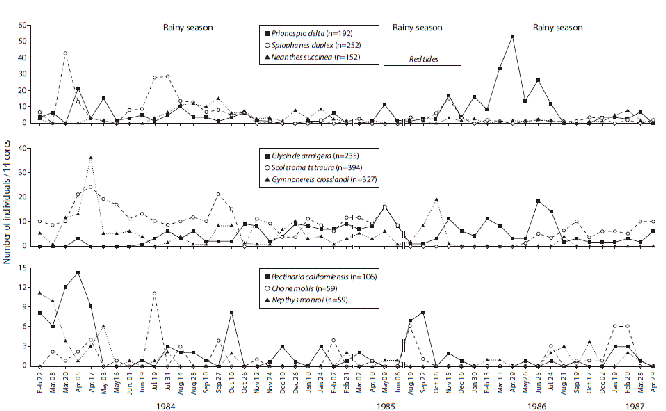
Figure 5: Numbers of nine selected species of polychaete worms. The extents of the rainy seasons (lower salinities) and red tide outbreaks are indicated. Gulf of Nicoya estuary, Pacific coast of Costa Rica. Punta Morales sand-mud flat (February 1984 to April 1987, 49 dates).
Table 3: Examples of spatial patchiness: species name and date with the maximum (within brackets) number of individuals found. Distribution of individuals among the 14 cores (core area 17.7 cm2 - core depth 15 cm). Punta Morales sand-mud flat. Gulf of Nicoya, Costa Rica. 1984-1987
Results of a Principal Components Analysis (Figure 6) based on the abundances of the 43 species during 1984-1987 evidenced a change in the composition of the polychaete fauna: Dry season dates 1-6, and rainy season dates 10-16 of 1984 were grouped apart. Similar results occurred with rainy season dates of 1985 (28-34) and dry season dates of 1986 (33-38). Moreover, dates previous to the red tide events (starting around date 29) were scored at positive values of PC-1, and show less dispersion on the axes than dates after the red tide event (gray shaded in Figure 6). At around the mid rainy season of 1986 (date 42) the polychate composition became more similar (dates 43-49 grouped with pre blooms dates) to that of pre-red tide events. A two way ANOSIM indicated a small seasonal change in the composition of the polychaetes (R=0.10, p=0.022), and supported the changes after date 29 (June, 1985) when red tide events started (R=0.49, p<0.001).
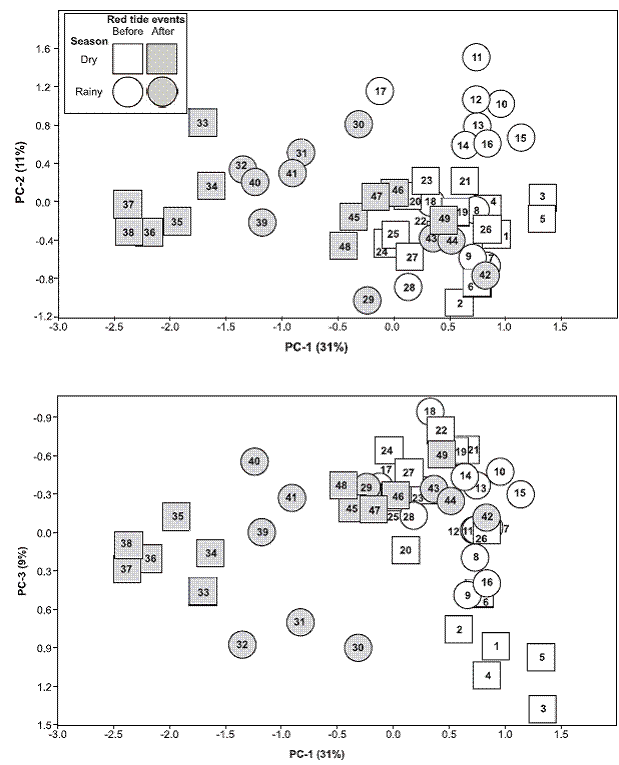
Figure 6: Principal Component Analysis (PCA) based on log10 (x+1) transformed abundances of 43 species of polychate worms collected during 49 dates (labelled 1 to 49). PC1, PC2 and PC3 explain 51 % of the variance. Squares (dry season dates), circles (rainy season dates). Gray shaded figures represent dates after the red tide outbreaks of 1985. All dates listed in Table 2. Gulf of Nicoya estuary, Pacific coast of Costa Rica. Punta Morales sand-mud flat, February 22, 1984 (Date 1) to April 29, 1987 (Date 49).
Results of the application of the General Additve Models to the M. californiensis 1984-1987 data indicates higher abundances during the dry season (Figure 7A, B) of each year (F=17.75, d.f.=1, p=0.001) and confirmed observations reported by Vargas (1987,1988a,1989) but no additional temporal trend was found during the study period (F=0.16, d.f.=1, p=0.692) (Figure 7C). During 19941996, this species lacked seasonal peaks in abundance (F=2.18, d.f.=1, p=0.153)(Figure 7D, E), but presented a long temporal trend (F=3.61, d.f.=3.4, p=0.024), with decreasing in abundances from July 1994 to December1995. Abundances increased again in later sampling dates (Figure 7F). On the other hand, P. alata had peaks of abundance during the 1984-1987 rainy season (Figure 8A, B) of each year (F=8.57, d.f.=1, p=0.005); and no additional temporal trends were found (F=0.86, d.f.=1, p=0.358) (Figure 8C). During 1994-1996 no seasonal peaks were evident (F=2.14, d.f.=1, p=0.157) (Figure 8D, E), but the population decreased from July of 1994 to December 1995, and then subsequently increased (F=3.61, d.f.=3.4, p=0.024), Figure 8F.
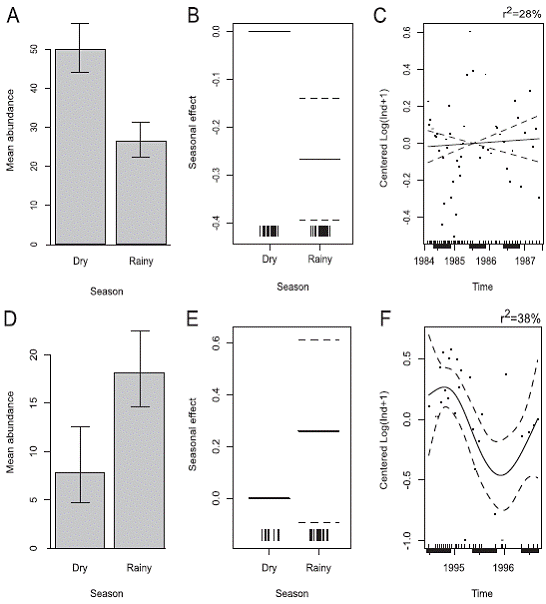
Figure 7: Outputs of the Generalized Additive Models (GAM) for the abundances of the capitellid Mediomastus californiensis (1984-1987, 49 dates). D, E, F: (1994-1996, 28 dates). A, D: Seasonal (dry vs rainy) mean abundance with 95% confidence limits. B, E:Seasonal effect. C, F: Remnant long term temporal trend. Solid line is the fitted value for the model. Broken lines are the Bayesiancredible intervals. Dark bars at the bottom = rainy seasons. Punta Morales sand-mud flat, Gulf of Nicoya estuary, Pacific, Costa Rica.
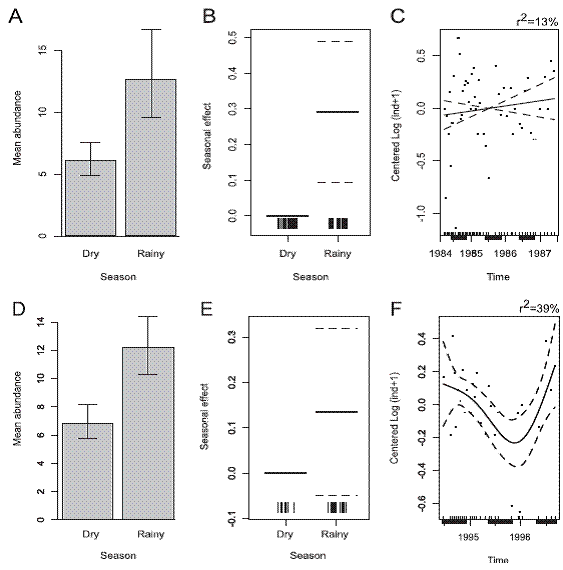
Figure 8: Outputs of the Generalized Additive Models (GAM) for the abundances of the spionid Paraprionospio alata (1984-1987, 49 dates).D, E, F: (1994-1996, 28 dates). A, D: Seasonal (dry vs rainy) mean abundance with 95% confidence limits. B, E: Seasonal effect.C, F: Remnant long term temporal trend. Solid line is the fitted value for the model. Broken lines are the Bayesian credible intervals.Dark bars at the bottom = rainy seasons. Punta Morales sand-mud flat, Gulf of Nicoya estuary, Pacific, Costa Rica.
Discussion
Vargas (1987, 1988a, 1989, 1996) discussed the results of his survey of the intertidal benthos focusing on structural aspects (fluctuations in the numbers of individuals and species in space and time) of the benthic community as an ecological unit. His research was followed by more detailed evaluations of the data sets focusing on the population dynamics of the main taxonomic groups found at the site (Vargas& Dean, 2010; Vargas& Solano, 2011; Vargas-Zamora & Sibaja-Cordero, 2011; Vargas-
Zamora, Sibaja-Cordero &Vargas-Castillo, 2012). In these later works the lists of 23 mollusks and 29 crustacean species were updated. The fauna also included several (March 1985 to April 1987).
species of flatworms, nemerteans, sipunculans, and a brachiopod (Vargas, 1987, 1988a, 1989, 1996; Dittmann &Vargas, 2001) During the 1984-1987 study an updated total of 43 species of polychaete worms were found distributed among 6 600 individuals (Table 1) bringing the total number of known macrofaunal species collected by coring at the 400m2 plot to 112. During the 1994-1987 and 1994-1998 surveys Polychata was the most important group in terms of the number of species found at the intertidal flat. The updated list of polychaetes includes 15 name corrections (Table 1). Of particular importance are the updated names of the numerically dominant spionids, C. calafia, S. duplex, and P. alata, whichwere based mainly on Blake (1979), Meisner & Hutchings (2003), and Yokoyama (2005), respectively. However, eight species remain to be identified (Table 1) The relative importance of these numbers of species and individuals in the context of discussions about tropical benthic diversity has been already addressed by Maurer &Vargas (1984), Alongi (1989), Dittmann &Vargas (2001) and Dittmann (2002), among others.
A total of 317 species of polychaetes listed in the literature for the Pacific coast of Costa Rica was reported by Dean (2009). He stated that the number of polychaetes from Central American coasts is lower when compared to other geographical regions, a fact that may be related to lower sampling efflort. Maurer,Vargas, & Dean (1988) conducted a grab survey at 41 stations in the Gulf of Nicoya ranging in depths from 1 to 46 m and a total of 125 species (retained on a 500 micron mesh sieve) of polychaetes were identified. In the deep fjord-like Golfo Dulce estuary located south of the Gulf of Nicoya a total of 47 polychaete species were collected in 139 corers (same core and mesh size as in this study) taken at nine stations ranging in depth from 43 to 200 m (León-Morales &Vargas, 1998). On the Pacific coast of Central America, Molina-Lara &Vargas-Zamora (1995) conducted an evaluation of the polychaete fauna at 15 stations along a shallow estuary in El Salvador (13oN) using the same coring device and mesh size as in this study and identified 30 species. Dittmann &Vargas (2001) compared the faunas of Punta Morales and El Salvador with those of similar habitats in Australia and although there were no polychaete species names in common certain species appear to be playing similar ecological roles at both sides of the Pacific. This similarity of ecological roles is interesting and a promising avenue for future research to understand how energy flows in tidal flats at different latitudes. Dominance by deposit feeders for instance was a feature in common between Australia and Central America.
A feature of the Punta Morales polychaete assemblage is the numerical dominance by the capitellid Mediomastus californiensis at 33 of the 49 dates (Table 2). M. californiensis, (a sub-surface deposit feeder) has been reported from both sides of the American Continent. For instance, Gaston, Lee & Nasci (1998) found no seasonal population trends in this species in a coastal lagoon on the Northern Gulf of Mexico, where it reached densities ranging from 267 to 1258 individuals / 0.5 m2. If the maximum density of 118 individuals of M.californiensis / 0.0248 m2 (14 cores) included in Table 2 is extrapolated to 0.5 m2 a value of 237 ind. / 0.05 m2 is possible for Punta Morales. This number is similar to the minimum found in the coastal lagoon. M. californiensis has been reported by Weston (1990) as a deep burrowing (2-10 cm) worm that prefers shallow fine sandy sediments with low organic carbon content in waters of Puget Sound, Pacific coast of North America. At the Gulf of Nicoya M. californiensis was found at only 19 out of 41 subtidal grab station (Maurer &Vargas, 1984). This low figure may indicate that shallow and uneven grab penetration underestimated abundances. Cores taken in Punta Morales were cut at the 15 cm depth, which may have better sampled the population of this and other deep burrowing wormsDominance at other dates was due to numbers of the surface depostit feeding spionid polychates P. alata, S. duplex and C. calafia (Table 2). Thus, the polychaete assemblage of the Punta Morales sand-mud flat is dominated by surface and subsurface deposit feeding polychaete worms. However, it is relevant the presence of other less numerically important species that could play key structuring roles due to their feeding habits as carnivores, omnivores, and suspension feeders. This variety of roles indicates the presence of a relatively well organized food web. A literature survey on tropical flats from different latitudes was conducted by Dittmann (2002) who concluded that the occurrence of macrobenthic species appears to be site specific and the composition of the local assemblages seems to be influenced by the presence of certain species (ecosystem enginneers) that through their living activities (tube building, construction of burrows, sediment reworking) influence the presence or absence of other species.
Similarities between tidal flats seem to increase at higher taxonomic levels. Dittmann &Vargas (2001) found that there were 26 genera, and 30 families in common between Austrlia and Central America. A total of 25 families are included in Table 1. As a comparison, 35 polychaete families were found by Barrio-Frojan, Kendall, Paterson, Hawkins, Nimsantijaroen & Aryuthaka (2006) at five nonvegetated intertidal habitats in Thailand (9o N) in sediments composed of similar percentages of sand (51 to 71%) to that of Punta Morales (65%). With the exception of the Pectinaridae and Trochochaeidae found in Punta Morales, the remaining 23 families were also found in Thailand. Moreover, in Thailand nine families were represented by less than 5 individuals, while 10 families were so in Punta Morales. Both sedimentary environments were dominated by deposit feeders.
The polychaete fauna of the Punta Morales estuarine intertidal sand-mud flat appears patchy both in time and space. Data included in Figs. 2, 3, 4 and 5 provide evidence of patchiness on a temporal scale, while data included in Table 3 do so at the spatial scale. Spatial patchiness may be due to several factors ranging from settling preferences of larvae to survival of individuals after predation or other disturbances (Thrush, 1991). This area of research remains little addressed in tropical benthic studies. Polychaete worms usually comprise the highest percentage of macroinvetebrates found in sediment surveys and have quick population response to environmental disturbances at least in temperate latitudes (see Gray & Elliot, 2010).
The populations of polychaetes from the Punta Morales sand-mud flats may have reacted to two environmental disturbances of different spatial and temporal scales: At the regional scale and as previously discussed by Vargas-Zamora, Sibaja-Cordero &Vargas-Castillo (2012) sampling at the flat started in February 1984, several months after the strong (October 1982 - July 1983) El Niño Southern Oscillation (ENSO) high water temperatures returned to near normal values on the Pacific coast of Central America. The impact of this ENSO on sandy beach communities of Peru was reported by Arntz, Brey, Tarazona & Robles (1987). They found that invasions of macrobenthic species were more frequent during ENSO, and after November 1983 spionid polychaetes (Dispio and Scolelepis) became members of the community. The polychaete population patterns illustrated in Figs. 2, 3, 4, 5, 6, 7, 8 provide evidence in support of the hypothesis that the benthic macrofauna of the Punta Morales flat was impacted by the 1982-1983 ENSO. Abundances during the first sampling months are indicative of declining populations. Higher abundances of flatwormrs, ostracods, and cumaceans were also found early in 1984 and declined afterwards (Vargas, 1987; Vargas-Zamora, Sibaja-Cordero &Vargas-Castillo, 2012), However, these changes were not as evident with the mollusks (vargasZamora & Sibaja Cordero, 2011).
At the Gulf of Nicoya scale the Punta Morales data set provide evidence of the response of the polychaete assemblage to an environmental disturbance that took place in this estuary in 1985. The results of a PCA (Figure 6) indicate that a change in polychaete diversity and abundance is evidenced by the separation of the sets of cores collected in May and June, 1985 (dates 28 and 29) from the cluster of previous dates. This separation was clear by date 30 (August 19) and continued to around date 41 (July 24, 1986), when faunal composition returned to be similar to that of the dates from early 1985 and before. Of particular relevance to the observed changes in the polychate assemblage detected by PCA are the ocurrences from June to November (rainy season) of 1985 of red tide patches in the mid upper Gulf of Nicoya.
Viquez & Hargraves (1995) described the occurence of dinoflagellate bloms (Cochlodinium catenatum, Gymnodinium catenatum, and the non-toxic Prorocentrum balticum) in the Gulf of Nicoya estuary from January 1985 to March 1986. These patches were frequent and dominated by C. catenatum until November, when G. catenatum became dominant. Dense patches of C. catenatum were found within the Punta Morales estuary, a few hundred meters North from the intertidal flat (Figure 1). Although no sediment disturbances were observed at the time, a few dying Tellina clams were found on the sediment surface on August, 1985 (Vargas-Zamora & Sibaja-Cordero, 2011). Moreover, fish mortalities restricted to species of the family Sciaenidae were reported in the inner Gulf of Nicoya during late September and mid October 1985 (Szelistowski & Garita. 1989). Red tides apparently also reduced the densities of eggs and larvae of anchovies (Engraulidae) in and adjacent to the Punta Morales estuary (Ramírez, Szelistowski & López, 1989).
The impact of microalgal blooms on the benthos has been reported previously only from a few locations in temperate and subtropical latitudes. The results of the PCA based on polychaete abundances at the Punta Morales site are the first for this region of the tropical Pacific.
Dauer & Simon (1976) studied the impact of the 1971 dinoflagellate (Gymnodinium breve) bloom which caused anaerobic conditions in waters of the shallow Tampa Bay. Florida (28oN). Among the 22 more abundant species, the bloom reduced the total numbers of individuals and species by 97% and 77%, respectively. Some species with important densities (ind/m2) in 1970 dissapeared (0 ind/ m2) after the 1971 event and were found again in 1973, for instance: the onuphids Onuphis eremita (283 ind. in 1970- 0 ind. after -3 ind. in 1973) and Diopatra cuprea (81-0-1), the nereidid Nereis succinea (33-7-3), and the glycerids Glycera capitata (16-0-2) and G. ameriana (3-045). Species like Scolelepis texana had a diffeent patern (16-13-2). A total of 54 polychate species was found during their study and nine of them represented 79.6% of the total number of individuals. Among these nine species, Paraprionospio pinnata, Polydora ligni, Nereis succinea, Magelona pettiboneae, Eteoneheteropoda and Capitita ambiseta, have representative genera in the Punta Morales intertidal flat (Table 1), indicating similar trophic roles of the polychaete assemblages.
Olsgard (1993) found significant reductions in the number of individuals and species in one year following a bloom of a toxic dinoflagellate (Chrysochromulina polylepis) in the coastal waters of Norway (580N). The change in faunal composition of the subtidal benthos became evident after a multivariate technique (DCA) was applied to the data set. This technique separated his data sets of 1987-1988 from those of 1989-1990. DCA also placed the 1990 set closer to that of 1987, indicating a tendency of the fauna to return to pre-bloom struture. When abundances before and after the bloom were compared, certain polychaete species showed reductions in abundance ranging from 25 to more than 80%. After the bloom many species of polychaetes present in low numbers dissapeared from more of 30% of the stations sampled. The capitellid Heteromastus filiformis was first among the ten more abundant species during the study period and this rank was not affected by the bloom. No increases in abundance of opportunistic species (including H. filiformis and Spiophanes kroyeri) was observed after the bloom.
Wear & Gardner (2001) studied the impact of the 1998 bloom caused by the toxins of the naked dinoflagellate Karenia brevisulcata in Wellington Harbour (41o S), New Zealand. They found that the four main groups of organisms (polychaetes, mollusks, crustaceans and echinoderms) were equally affected. However, at one of the subtidal (11 m) stations at the entrance to the harbour the polychaetes were affected most, in comparison with the other three groups that had small increases or decreses in the number of individuals or species. In this context it is noteworthy that the spionid P. delta reached its peak of abundance in Punta Morales after the red tide oubreaks (Figure 5).
Of special interest are the oscillations in abundance of the spionid Polydora citrona (now Carazziela calafia, Table 1) described by Vargas (1989) C. calafia was first recorded at the site in December of 1984 (3 individuals) and appeared sporadically until September 1985, when numbers increased from 18 to peaks of 145, 285, 271, and 273 individuals in January, February, March and April of 1986, respectively. Numbers declined to 33 by May and returned to single digits by August, 1986. It is noteworthy that numbers of this spionidstarted to increase late (June to November of 1985) during the period of red tides and reached its maximum several months later.
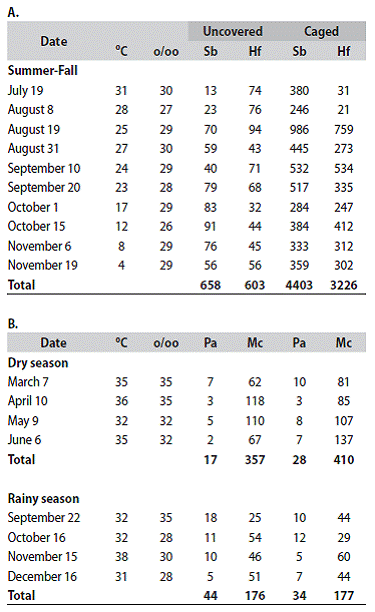
At the micro-scale level no significant changes in abundance of C. calafia were evidentin responseto the deployment of wire mesh cages on the sediment during the dry (Totals: 7 ind. outside vs 9 ind. inside) and rainy (Totals: 247 outside vs 147 inside) seasons of 1985 (Vargas, 1988a, 1996) Several species of the Spionidae and Capitellidae polychaete families are known to increase in abundance (Table 4A) in temperate latitudes inside wire mesh cages deployed to ameliorate the impact of macro-predators (see Reise, 1985). At the tropical site, however, the population changes inside vs outside cages of the spionid P. alata and the capitellid M. californienesis were negligible (Table 4B).
In addition to these disturbances the Gulf of Nicoya and its biota is under the influence of the more predictable seasonal salinity cycle. Vargas (1987, 1988) indicated that the whole benthic macrofaunal community had a seasonal response as evidenced by the results of Cluster and Multiple Discriminant Analysis. Moreover, at the population level several species presented seasonal oscillations, being more evident in M. californiensis and P. alata. Three main results of the application of General Additive Models (GAM) analysis to the abundances of M. californiensis and P. alata (Figs. 7, 8) during the period of 1984-1987 and 1994-1998 were obtained: First, it confirmed Vargas (1987, 1998) observations that M. californiesis was more abundant during the dry seasons, while P. alata was so during the rainy seasons. Second: seasonality of these species was not detected in the 1994-1996 data set. Third: previously unnoticed underlying patterns of unknown origin were detected by the application of GAM.
Table 4: Water temperature (oC), salinity, (o/oo) and total number of individuals per date for: A. Streblospio benedicti (Sb) and Heteromastus filiformis (Hf ) in cores from uncovered and coveredsediments (cage deployed June 17, 1977). Rehoboth bay shallow sandy coastal lagoon, Delaware (39º N). Atlantic. U.S.A. (Data from Vargas, 1979). B. Paraprionospio alata (Pa) and Medionastus californiensis (Mc) in cores from uncovered and covered sediments (cages deployed Feb. 7, May 9, Aug. 19 , Nov. 15, 1985). Punta Morales (10o N) intertidal sand-mud flat, Pacific coast, Costa Rica
Most of the ecological theories that guide the interpretation of the results of marine sediment surveys have been developed based on data from temperate latitudes. The books by Little (2000) on soft shores and estuarine habitats, and by Gray & Ellliot (2010) on marine sediments in general address those theories. A better understanding of the ecology of tropical benthos could benefit from more long-term monitoring and the aplication of user-friendly statistical methods in search for patterns. Data collected over periods of more than a year continue to be rare in tropical marine benthic literature. Moreover, climatic change and increased costal development make these data highly valuable for future comparisons. The Gulf of Nicoya estuary has been heavily influenced by sedimentation, mangrove forest alteration, pollution, and fishery methods leading to bottom damage (shrimp trawling), removal of top predators (shark fishing) and extraction of filter-feeder shellfish (Vargas& Mata, 2004). Borja, Dauer & Elliott (2010) point out that recovery of estuarine components may take from less than five years to more than 25 depending on the nature of the impacted component. In this context a clear knowledge of the abundance patterns of polychaetes and other groups must be at hand to evaluate potential recoveries after stressors have been removed or their effects ameloriated.

