











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
El sector cacaotero representa uno de los rubros económicos más importantes del Ecuador (García-Cruzatty et al., 2020). El material genético CCN-51, está entre los cultivares con mayor extensión en el país por su alta productividad y adaptabilidad a las diversas condiciones climáticas (Boza et al., 2014). Sin embargo, este clon es susceptible a patógenos como Moniliophthora roreri, Phytophthora palmivora y Moniliophthora perniciosa, cuya prevalencia se incrementa con una elevada humedad relativa, baja temperatura y las deficiencias nutricionales del cultivo (Gutiérrez et al., 2016; Villamizar-Gallardo et al., 2019).
El empleo de insecticidas (Carbofuram, Metomyl, Phoxim) y fungicidas (cúpricos y triazoles) mitiga las plagas y enfermedades en el cultivo de cacao (Boateng et al., 2023). Además, para suplir las necesidades nutricionales del cultivo se utilizan fertilizantes nitrogenados, fosforados y potásicos (Yaseen et al., 2021). Sin embargo, se ha reportado que el uso prolongado de estos insumos químicos desencadena una reducción en la eficiencia de sus componentes activos (Chandini et al., 2019), así como la resistencia de plagas y patógenos (Akrofi et al., 2017; Roman et al., 2021).
El empleo de rizobacterias promotoras de crecimiento vegetal (plant growth promoting rhizobacteria, PGPR por sus siglas en inglés) es una alternativa para controlar patógenos y mejorar la eficiencia en la absorción de nutrientes (Babu et al., 2015). Una de sus principales características es la acción solubilizadora de nutrientes y biocontroladora por la liberación de componentes enzimáticos y ácidos orgánicos (Devi et al., 2022; Jha & Subramanian, 2018). Ejemplo de ello es la producción de exoquitinasas chiA, implicada en la degradación de quitina, componente principal de la pared celular de hongos fitopatógenos como ascomicetos y basidiomicetos (Morales-Ruiz et al., 2021). Esta funcionalidad permite que exista una mayor utilización de las PGPR, sobre todo en problemas nutricionales y en el control de enfermedades; sin embargo, su multiplicación masiva es una de las limitantes para su producción (Tirado-Gallego et al., 2016; Tyśkiewicz et al., 2022).
Los bioformulados PGPR de bajo costo son una alternativa para aumentar la producción de microorganismos con una menor inversión (Mishra et al., 2020; Sarmiento-López et al., 2022). El desarrollo de esta tecnología se concentra en disponer fuentes nutricionales adecuadas en la multiplicación y protección de la biomasa bacteriana (Lobo et al., 2019), dado que los microorganismos requieren de medios específicos (Anith et al., 2017). La efectividad de los productos biológicos de origen bacteriano depende del número de colonias viables en el tiempo y del pH. Por tal motivo, el contenido de carbohidratos, proteínas y macro-microelementos es esencial para una óptima multiplicación y crecimiento (Chandra & Sharma, 2021).
Las aplicaciones de bioformulaciones de PGPR en la agricultura generan una aceptación por mejorar las características morfológicas y productivas (Sriwati et al., 2019). Así por ejemplo, se aplicaron formulaciones de Bacillus sp., para promover el sistema radicular de plántulas de cacao (Thomas et al., 2011). Otras investigaciones evidenciaron la eficacia de Pseudomonas aeruginosa y Chryseobacterium proteolyticum en la reducción de P. palmivora, donde se confirmó una inhibición del 100 % en mazorcas de cacao (Alsultan et al., 2019), y que las aplicaciones de microorganismos al suelo incrementan la solubilización de minerales y mejoran las características morfológicas y productivas en T. cacao (Hipólito-Romero et al., 2017).
Con el fin de resolver los problemas de manejo nutricional y fitosanitario del cultivo de cacao, además de promover el manejo sostenible del cultivo con alternativas inocuas y de bajo costo, la presente investigación tuvo como objetivo evaluar el efecto de los bioformulados sobre la viabilidad celular de las rizobacterias y su influencia en T. cacao L.
Materiales y métodos
Descripción de área de estudio
La investigación se ejecutó en dos etapas. La primera fase se llevó a cabo en el Laboratorio de Microbiología y Biología Molecular de la Universidad Técnica Estatal de Quevedo (UTEQ), en el Campus Universitario Manuel Haz Álvarez, ubicado en el km 1,5 vía Quevedo-Santo Domingo, Ecuador (01° 00’ 44” latitud Sur y 79° 28’ 22” de longitud Occidental), a 73 m s. n. m. La segunda fase se desarrolló en la finca Ignolia, localizada en el recinto el Moral, cantón La Maná, provincia de Cotopaxi, Ecuador (0° 57’ 38” latitud sur y 79° 16’ 22” longitud occidental), con una altitud de 187 m s. n. m.
Caracterización bioquímica de la capacidad solubilizadora de minerales
Las cepas bacterianas (Cuadro 1) se reactivaron de forma individual en medio de cultivo King B líquido (KBL) a 28 °C a 180 rpm por 24 h, y se usaron para determinar la capacidad hidrolítica de urea a dióxido de carbono por el método de Christensen (1946), la actividad fosfatasa (Pikovskaya, 1948) y la producción de ácidos orgánicos para solubilización de potasio (Hu et al., 2006) y de zinc (Saravanan et al., 2004). La generación de biofilm se determinó de forma cualitativa por el método de cristal violeta (Alexander & Zuberer, 1991) y producción de quitinasa (Moon et al., 2017). En cada prueba se empleó como control negativo agua desionizada estéril.
Cuadro 1 Características de las rizobacterias con producción de metabolitos antagónicos, sideróforos, producción de ácido indol-3-acético y cultivo de procedencia. Ecuador, 2020.
| Organismo | Cepas | Metabolitos antagónicos | Sideróforos | Producción de AIA | Cultivo de procedencia | |||
| PR | HCN | Prn | DAPG | |||||
| A. calcoaceticus | BMR2-12 | + | + | Banano | ||||
| S. marcescens | PM3-8 | + | + | + | Banano | |||
| P. protegens | CHA0 | + | + | + | + | + | + | Tabaco |
| P. veronii | R4 | + | + | + | + | Uva | ||
| E. asburiae | PM3-14 | + | + | + | + | Banano | ||
PR: proteasa. HCN: cianuro de hidrógeno. Prn: Pirrolnitrina. DAPG: 2,4-diacetilfloroglucinol, sideróforos. AIA: producción de ácido indol-3-acético (Chávez Arteaga et al., 2020; Montes et al., 2016; Peñafiel Jaramillo et al., 2016). / PR: protease. HCN: hydrogen cyanide. Prn: pyrrolnitrin. DAPG: 2,4-diacetylphloroglucinol, siderophores. IAA: production of Indole-3-acetic Acid (Chávez Arteaga et al., 2020; Montes et al., 2016; Peñafiel Jaramillo et al., 2016).
La producción de ureasa, fosfatasa, ácidos orgánicos y quitinasa se evaluó por el tamaño del halo. La presencia de biofilm se determinó por la aparición de biopelículas formadas en los tubos de ensayo por la adhesión de bacterias. Se definieron valores por la intensidad del anillo en el tubo. Los valores se registraron de acuerdo con la metodología definida por Holt et al. (1994), utilizando los siguientes valores cualitativos: mayor presencia (++), presencia (+) y ausencia (-).
Determinación de la presencia del gen chiA por PCR
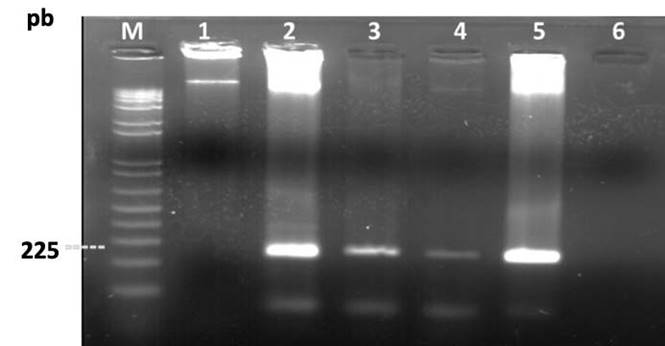
El ADNg bacteriano se obtuvo al emplear PureLink™ Genomic DNA Mini Kit - Thermo Fisher Scientific, según el protocolo descrito por el fabricante. Para la amplificación molecular, se emplearon los primers específicos chiAF (5’GATATCGACTGGGAGTTCCC3’) y chiAR (5’CATAGAAGTCGTAGGTCATC3’), con producto de amplificación 225 pb (Ramaiah et al., 2000). La reacción de PCR se realizó en 20 µL, con 5,0 μL Buffer (1X); 1,2 μL de MgCl2 (6 mM); 1,0 μL de DNTP`s (0,2 mM); 1 μL / cada primer (0,2 mM); 0,2 μL de Taq polimerasa (Promega®); 1 μL de ADN (50 ng/mL), y 11,6 μL de H2O. Se empleó un termociclador (TECHME®) y se ajustó su perfil térmico a 94 °C durante 4 min; 35 ciclos de desnaturalización a 92 °C; alineamiento a 58 °C, y polimerización a 72 °C por 1 min, seguidos de 7 min de extensión a 72 °C. Los productos de amplificación se verificaron por electroforesis en gel de agarosa al 1,5 % y teñidos en bromuro de etidio. Los fragmentos se determinaron por comparación con el marcador de peso molecular 100 pb (Invitrogen®).
Bioformulaciones líquidas para la determinación de la viabilidad celular de las rizobacterias
Las cepas Acinetobacter calcoaceticus BM2-12, Serratia marcescens PM3-8, P. protegens CHA0, P. veronii R4 y Enterobacter asburiae PM3-14, provenientes del banco de germoplasma de la Universidad Técnica Estatal de Quevedo, seleccionadas por su propiedad antifúngica, nematicida y promotoras de crecimiento vegetal (Cuadro 1), fueron reactivadas. Luego, se cultivaron de forma individual en 100 mL de KBL ((g/L): peptona de carne 20,0; glicerol, 15,0 mL; fosfato dipotásico, 1,5; sulfato de magnesio heptahidratado, 1,5; agua destilada (pH 7,2)), suplementado con chloramphenicol (13 μg/mL), ampicilina (40 μg/mL) por 48 h (King et al., 1954) (Figura 1A).

Figura 1 Proceso de multiplicación celular de las rizobacterias en formulaciones líquidas. A) Crecimiento del preinóculo bacteriano. B) Fermentación de las bacterias en tanques de 50 L. C) Concentración celular 24 horas después del análisis microbiológico. Quevedo, Ecuador, 2020.
El preinóculo de las cinco bacterias se recuperó por sedimentación y se aplicó de forma individual un volumen de 100 mL a una concentración de 1,0E+8 UFC/mL en 40 L de medio de cultivo alternativo (Cuadro 2). Se procedió a su respectiva fermentación y/o masificación de forma individual mediante fermentadores con capacidad de 50 L. Se mantuvieron en agitación con una presión de aire de 0,007 MPa durante 24 h; cada fermentador contenía un filtro de nitrocelulosa de 0,45 micras x 47 mm a entrada y salida del flujo de aire (Figura 1B). Al finalizar las 24 h de agitación, se colectaron tres muestras de 50 mL de cada tratamiento en tubos Falcon. El número de muestras forma parte de las repeticiones; se tomaron en consideración tres unidades experimentales por cada repetición y se conservaron a temperatura ambiente (25 °C) durante 26 días.
Cuadro 2 Composición de los diferentes formulados para el crecimiento y viabilidad de rizobacterias. Ecuador, 2020.
| Formulado | Ingrediente | |
| BIO-M3 | Melaza (g/L) | 20 |
| Harina de maíz (g/L) | 20 | |
| Sal en grano (g/L) | 5,0 | |
| Glicerina (mL) | 7,0 | |
| pH | 6,5 | |
| PGPRs-4N | Melaza (g/L) | 20 |
| Harina de maíz (g/L) | 20 | |
| Sal en grano (g/L) | 5,0 | |
| Roca fosfórica (g/L) | 1,5 | |
| Glicerina (mL) | 7,0 | |
| pH | 6,5 | |
| BIO-IMPULSE | Melaza (g/L) | 20 |
| Harina de maíz (g/L) | 20 | |
| Sal en grano (g/L) | 5,0 | |
| Roca fosfórica (g/L) | 1,5 | |
| Glicerina (mL) | 7,0 | |
| Aceite mineral (mL) | 5,0 | |
| pH | 6,5 | |
| BIO-QPGPRs | Melaza (g/L) | 10 |
| Harina de maíz (g/L) | 20 | |
| Sal en grano (g/L) | 5,0 | |
| Roca fosfórica (g/L) | 1,5 | |
| Glicerina (mL) | 7,0 | |
| Fijador (mL) | 5,0 | |
| Lixiviado de raquis de banano (mL) | 15 | |
| pH | 6,5 |
Variables evaluadas en los bioformulados
Para determinar la viabilidad celular de las rizobacterias se recuperaron muestras de 100 ul a (1, 6, 11, 16, 21 y 26 días) después de su proceso de fermentación. Se evaluaron las siguientes variables: unidades formadoras de colonias por mililitro (UFC/mL) en tiempo y potencial de hidrógeno (pH), con base en las metodologías de Dickson (1993) y Sharpe y Kilsby (1971) (Figura 1C).
Aplicación de bioformulados en plantaciones de T. cacao CCN-51
El bioformulado se seleccionó por su mayor viabilidad celular de las rizobacterias evaluadas de la fase in vitro, para la multiplicación y/o masificación. El bioformulado se fermentó durante 24 h y se homogenizó en relación v/v al 20 %. Se obtuvo una concentración bacteriana de 7,6E+6 UFC/mL para su aplicación en plantación de CCN-51 de cuatro años de edad. La forma de aplicación de los bioformulados con sus respectivos controles se describe en el Cuadro 3.
Cuadro 3 Descripción de la forma de aplicación de los bioformulados. Ecuador, 2020.
| Aplicación | Descripción | Frecuencia de aplicación durante el año 2020 |
| Edáfico | Se realizó una limpieza de 0,60 m desde el centro del tallo hacia la corona de la planta, y se aplicaron, vía edáfica, 250 mL/planta de bioformulados al 20 % en relación v/v, con la ayuda de un pulverizador de mochila Cooper Pegler CP3 Classic Knapsack Sprayer CP846320. | Cuatro veces (enero, marzo, julio, septiembre) |
| Foliar | Se esparcieron al área foliar 300 ml/planta de bioformulados al 20 % v/v mediante una fumigadora. | |
| Inyección | A una altura de 0,20 m desde la base del tallo, se perforó un hoyo de 8 cm de profundidad con un diámetro de 1,2 cm y una inclinación de 45°, con 10 mL/planta de bioformulado al 20 % en relación v/v. La perforación se selló con parafilm. | |
| Químico | Se aplicó fungicida de forma foliar 3 kg/ha de oxicloruro de cobre (Oxithane) y 1 kg/ha de fertilizante completo soluble 13-40-13 (kristalon - YaraTera). | Tres veces (enero, abril, agosto) |
| Control | De forma foliar, se aplicaron 200 ml de agua destilada |
Variables evaluadas en el cultivo
Las variables evaluadas en CCN-51 fueron las siguientes: a) número de emisión foliar (brote joven), para lo cual se contabilizaron los brotes dos meses después de la poda de formación y mantenimiento; b) incidencia de Phytophthora spp., M. roreri; c) rendimiento del cultivo (kg/ha), con base en la metodología descrita por Anzules Toala (2019), y d) número de granos, que se obtuvo al contar el total de semillas de cada mazorca. Estas variables fueron obtenidas cada mes (temporada lluviosa y seca), durante un año.
Labores culturales
Como labores culturales se realizaron dos podas (formación y mantenimiento) durante el año, en los meses de enero y junio. Además, se realizó un control de maleza cuatro veces al año, con aplicaciones de paraquat (2,0 L/ha) y desmalezadora a motor. La fertilización química se realizó en tres aplicaciones fraccionadas de urea (N46 %) 150 g/planta; DAP (N 18 %, P 46 %) 70 g/planta; KCl (K 60 %) 150 g/planta y sulfato de Mg (Mg 27% y S 20%) 29 g/planta. La aplicación del fertilizante se realizó al voleo, mediante aplicación directa al suelo, desde la base de la planta a un metro a la redonda lo que cubrió la corona de forma homogénea (Ruales et al., 2011).
Diseño experimental y análisis de datos
En el experimento de viabilidad celular de rizobacterias se aplicó un diseño completamente al azar (DCA) con arreglo factorial conformado por formulados y cepas. Se realizaron tres réplicas con tres unidades experimentales. A nivel de campo, en la aplicación de bioformulados en plantaciones de T. cacao CCN-51, se empleó un diseño factorial en bloques completamente al azar (DBCA), para su temporada y formas de aplicación. Cada tratamiento contenía cuatro repeticiones con cuatro unidades experimentales; se evaluaron dieciséis plantas por tratamiento.
Para garantizar la confiabilidad en la estadística paramétrica como análisis de varianza (ANDEVA), los datos fueron sometidos a normalidad mediante Kolmogorov-Smirnov y se aplicó la prueba de Bartlett para verificar el supuesto de homogeneidad de la varianza. Los datos fueron sometidos al ANDEVA y separados por procedimiento de comparación múltiple de Tukey, al nivel de significancia de p ≤ 0,05. Para el análisis se utilizó el software estadístico Infostat 2020 y la versión 19.1.0 del software Minitab (Di Rienzo et al., 2020; Minitab, 2021).
Resultados
Caracterización bioquímica con potencial a ser considerada
Las cinco rizobacterias mostraron la capacidad de hidrolizar urea y producir quitinasa, solubilizadora de fósforo, potasio y zinc de una fuente insoluble presente en el medio. Además, cuatro cepas fueron productoras de biofilm (Cuadro 4; Figura 2).
Cuadro 4 Selección de rizobacterias con capacidad para producir quitinasa, ureasa, fosfatasa, ácidos orgánicos y biopelículas. Ecuador, 2020.
| Cepas | Caracterización bioquímica | |||||
| Quitinasa | Fosfatasa | Ácido orgánico (potasio) | Ureasa | Ácido orgánico (zinc) | Biofilm | |
| BM2-12 | ++ | ++ | ++ | + | ++ | + |
| PM3-14 | + | + | ++ | + | + | - |
| PM3-8 | ++ | + | + | + | ++ | + |
| CHA0 | ++ | ++ | + | ++ | + | ++ |
| R4 | ++ | + | + | ++ | ++ | ++ |
Los signos indican: (-) ausencia, (+) presencia, (++) mayor presencia. / Signs indicate: (-) absence, (+) presence, (++) increased presence.
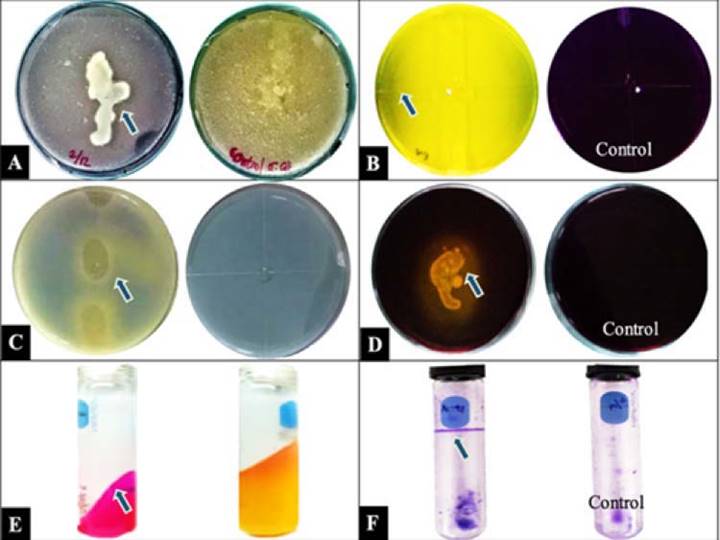
Figura 2 Producción enzimática de las PGPR para la solubilización de cada componente. A. Quitinasa. B. Ácidos orgánicos para la solubilización de potasio. C. Fosfatasas. D. Ácidos orgánicos para la solubilización de zinc. E. Ureasa. F. Biofilm. Flechas indican el sitio de liberación de compuestos por las PGPR. Ecuador, 2020.
Identificación del gen chiA por PCR
Se determinó la presencia de chiA, gen que codifica el dominio catalítico de las quitinasas bacterianas con un producto de amplificación de 225 pb en las rizobacterias (PM3-8, PM3-14, BM2-12, CHA0). No existió amplificación en la cepa R4 ni en el control negativo (agua) (Figura 3).
Determinación de la viabilidad celular y plataforma de multiplicación de rizobacterias
La generación de los bioformulados define valores significativos al proceso de multiplicación de rizobacterias, viabilidad celular y sus interacciones (p < 0,001) en los días (1, 6, 11, 16, 21 y 26). En cuanto a la viabilidad celular al día 1, los bioformulados BIO-QPGPRs, BIO-M3 y PGPRs-4N alcanzaron la concentración celular de CHA0 con 7,87E+11, 6,63E+9 y 4,80E+8 UFC/mL, respectivamente. La disponibilidad de los nutrientes del bioformulado BIO-IMPULSE mantuvo la viabilidad celular en las rizobacterias (R4 y PM3-8), con 1,05E+10 UFC/mL. Esto indica que el alto contenido de UFC/mL está relacionado con la alta disponibilidad de los nutrientes, que mejora la actividad biológica del bioformulado (Cuadro 5).
Cuadro 5 Unidades formadoras de colonias en bioformulaciones de rizobacterias durante 26 días de evaluación. Ecuador, 2020.
| Formulados | Bacterias | Día 1 | Día 6 | Día 11 | Día 16 | Día 21 | Día 26 | ||||||||
| BIO-QPGPRs | CHA0 | 7,87E+11 | A | 3,83E+8 | AB | 4,70E+7 | BC | 1,57E+6 | DEF | 1,60E+5 | DE | 1,83E+5 | A | ||
| BIO-QPGPRs | BM2-12 | 1,11E+10 | B | 5,50E+7 | EFG | 4,37E+7 | BCD | 3,23E+6 | CDE | 1,73E+5 | DE | 6,30E+4 | B | ||
| BIO-QPGPRs | PM3-14 | 6,17E+9 | B | 1,90E+8 | DE | 5,40E+7 | BC | 1,10E+7 | B | 1,43E+6 | C | 1,80E+5 | A | ||
| BIO-QPGPRs | PM3-8 | 1,27E+9 | B | 2,57E+8 | BCD | 6,07E+7 | B | 1,53E+6 | DEF | 2,10E+5 | DE | 1,63E+5 | A | ||
| BIO-QPGPRs | R4 | 1,27E+9 | B | 2,57E+8 | BCD | 6,07E+7 | B | 1,47E+6 | EF | 2,43E+5 | DE | 1,63E+5 | A | ||
| BIO-IMPULSE | CHA0 | 3,53E+8 | B | 1,27E+7 | FG | 3,73E+5 | F | 0 | F | 0 | E | 0 | C | ||
| BIO-IMPULSE | BM2-12 | 8,70E+7 | B | 3,90E+6 | G | 5,17E+5 | F | 0 | F | 0 | E | 0 | C | ||
| BIO-IMPULSE | PM3-14 | 6,07E+9 | B | 3,07E+6 | G | 2,67E+6 | F | 2,10E+4 | F | 0 | E | 0 | C | ||
| BIO-IMPULSE | PM3-8 | 1,05E+10 | B | 1,53E+8 | DEFG | 4,00E+5 | F | 0 | F | 0 | E | 0 | C | ||
| BIO-IMPULSE | R4 | 1,05E+10 | B | 1,53E+8 | DEFG | 4,00E+5 | F | 0 | F | 0 | E | 0 | C | ||
| BIO-M3 | CHA0 | 6,63E+09 | B | 4,77E+8 | A | 2,03E+8 | A | 2,70E+7 | A | 3,60E+6 | A | 0 | C | ||
| BIO-M3 | BM2-12 | 1,27E+08 | B | 1,60E+8 | DEF | 1,87E+7 | DEF | 4,83E+5 | F | 9,60E+4 | DE | 2,53E+4 | C | ||
| BIO-M3 | PM3-14 | 1,00E+9 | B | 3,50E+8 | ABC | 1,80E+7 | DEF | 2,70E+6 | CDE | 2,63E+6 | B | 0 | C | ||
| BIO-M3 | PM3-8 | 1,27E+9 | B | 2,27E+8 | CD | 2,93E+7 | CDE | 2,47E+6 | CDE | 2,13E+5 | DE | 0 | C | ||
| BIO-M3 | R4 | 1,27E+9 | B | 2,27E+8 | CD | 2,93E+7 | CDE | 2,93E+6 | CDE | 2,40E+5 | DE | 0 | C | ||
| PGPRs-4N | CHA0 | 4,80E+8 | B | 2,57E+7 | FG | 2,93E+7 | CDE | 2,30E+6 | CDE | 2,57E+5 | D | 1,60E+4 | C | ||
| PGPRs-4N | BM2-12 | 3,73E+7 | B | 1,20E+7 | FG | 6,63E+5 | F | 3,30E+5 | F | 2,73E+4 | DE | 0,00E+0 | C | ||
| PGPRs-4N | PM3-14 | 1,13E+8 | B | 7,13E+6 | G | 6,10E+6 | EF | 3,27E+6 | CD | 2,10E+5 | DE | 1,63E+4 | C | ||
| PGPRs-4N | PM3-8 | 4,73E+8 | B | 4,10E+7 | EFG | 1,23E+7 | EF | 3,00E+6 | CDE | 1,13E+5 | DE | 1,63E+4 | C | ||
| PGPRs-4N | R4 | 4,73E+8 | B | 4,10E+7 | EFG | 1,23E+7 | EF | 3,47E+6 | C | 1,10E+5 | DE | 1,63E+4 | C | ||
| P (formulados) | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | |||||||||
| P (bacterias) | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | |||||||||
| P (interacción) | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | |||||||||
Los valores de p indican el efecto de los factores sobre la variable dependiente mediante un análisis de varianza de dos vías (formulado × bacteria). Los valores con letras similares no presentan diferencias estadísticas significativas al nivel de p ≤ 0,05, según el procedimiento de comparación múltiple de Tukey. Al tiempo de muestreo (1, 6, 11, 16, 21 y 26 días). / The p values indicate the effect of the factors on the dependent variable using a two-way analysis of variance (formulated × bacterial). Values with similar letters do not present significant statistical differences at the level of p ≤ 0.05, according to Tukey’s multiple comparison procedure. At sampling time (1, 6, 11, 16, 21 and 26 days).
En cuanto a la viabilidad celular al día 6, los bioformulados BIO-M3 y BIO-QPGPRs lograron mantener la UFC/mL en 4,77E+8 y 3,83E+8 en la rizobacteria CHA0, respectivamente. La viabilidad celular se redujo en BIO-IMPULSE con 1,53E+8 UFC/mL, de las rizobacterias PM3-8 y R4 (Cuadro 5).
Viabilidad celular al día 11, disminuyó el número de células UFC/mL en BIO-IMPULSE, BIO-M3 y PGPRs-4N entre 3,73E+5 y 1,87E+7; a diferencia de BIO-QPGPRs que mantiene su concentración celular de 4,37E+7 y 6,07E+7 para las rizobacterias CHAO, M2-12, BM3-14, PM3-8 y R4. Sin embargo, el bioformulado BIO-M3 mantiene la concentración celular en 2,03E+8 UFC/mL para CHA0 (Cuadro 5).
Para el día 16, los bioformulados BIO-M3 y BIO-QPGRs mantuvieron la concentración celular en CHA0 y PM3-14 en 2,70E+7 y 1,10E+7 UFC/ml. La disponibilidad de células vivas para las rizobacterias CHAO, M2-12, PM3-8 y R4 tuvo un efecto negativo en BIO-IMPULSE que decreció en su totalidad la presencia de células vivas (Cuadro 5).
Al día 21, el bioformulado BIO-IMPULSE evidenció una respuesta negativa al proceso de almacenamiento de rizobacterias con la disminución de su totalidad de células vivas para BM3-14. Al contrario, BIO-QPGPRs y BIO-M3 mantuvieron la mayor viabilidad celular entre 1,60E+5 y 1,43E+6 UFC/ml, y 9,6E+4 y 3,60E+6 UFC/ml, respectivamente.
Por último, al día 26, el bioformulado BIO-QPGPRs suministró las condiciones óptimas para una mayor estabilidad celular a su almacenamiento en las rizobacterias CHA0, BM2-12, BM3-14, PM3-8 y R4 con 1,83E+5, 6,30E+4, 1,80E+5, 1,63E+5 y 1,63E+5 UFC/ml, respectivamente (Cuadro 5). Este bioformulado BIO-QPGPRs es la alternativa viable para masificar y preservar células bacterianas hasta los 26 días. Además, es un sustituto viable para la masificación de PGPR, por su bajo costo y viabilidad en el tiempo, para la aplicación en cultivos agrícolas en la segunda fase de investigación (aplicación en campo).
Determinación del pH de los formulados
Los niveles de pH en procesos de biofermentación y conservación de las células bacterianas determinaron valores significativos para los bioformulados, las rizobacterias y sus interacciones (p < 0,001) en los días (1, 6, 11, 16, 21 y 26). Se deduce que el cultivo bacteriano presenta una disminución considerable del pH en los días 6, 11, 16, 21 y 26, en comparación con el día 1. En la evaluación del pH al día 21, la composición del bioformulado BIO-M3 logró mantener un pH entre (4,07, 4,13 y 4,22) para las rizobacterias (CHA0 BM2-12 y PM3-8), respectivamente (Cuadro 6).
Cuadro 6 Potencial de hidrógeno (pH) en bioformulaciones de rizobacterias durante 26 días de evaluación. Ecuador, 2020.
| Formulados | Bacterias | Día 1 | Día 6 | Día 11 | Día 16 | Día 21 | Día 26 | |||||||
| BIO-QPGPRs | CHA0 | 4,29 | HI | 3,88 | E | 3,81 | EFG | 3,77 | K | 3,77 | G | 3,81 | EFG | |
| BIO-QPGPRs | BM2-12 | 4,21 | IJK | 3,66 | G | 3,37 | I | 3,31 | O | 4,34 | A | 4,43 | A | |
| BIO-QPGPRs | PM3-14 | 4,26 | HIJ | 3,98 | D | 3,93 | DEF | 3,98 | G | 3,99 | E | 3,92 | DE | |
| BIO-QPGPRs | PM3-8 | 4,35 | H | 3,84 | EF | 3,89 | EF | 3,94 | H | 3,95 | E | 3,85 | DEF | |
| BIO-QPGPRs | R4 | 4,13 | K | 3,84 | EF | 3,76 | EFGH | 3,81 | J | 3,81 | FG | 3,73 | FGHI | |
| BIO-IMPULSE | CHA0 | 4,30 | H | 3,78 | F | 3,92 | DEF | 3,90 | I | 3,85 | F | 3,74 | FGH | |
| BIO-IMPULSE | BM2-12 | 5,10 | C | 4,78 | B | 4,23 | BC | 4,12 | D | 4,07 | J | 3,74 | FGH | |
| BIO-IMPULSE | PM3-14 | 4,20 | JK | 3,86 | EF | 3,92 | DEF | 3,49 | M | 3,76 | G | 3,58 | IJ | |
| BIO-IMPULSE | PM3-8 | 4,86 | EF | 3,97 | D | 3,68 | FGH | 3,72 | L | 3,53 | H | 3,57 | J | |
| BIO-IMPULSE | R4 | 4,74 | G | 3,62 | G | 3,50 | HI | 3,46 | N | 3,42 | I | 3,33 | K | |
| BIO-M3 | CHA0 | 5,23 | B | 5,04 | A | 4,78 | A | 4,29 | B | 4,13 | C | 3,72 | FGHI | |
| BIO-M3 | BM2-12 | 5,05 | C | 3,91 | DE | 3,98 | CDE | 4,07 | E | 4,07 | D | 3,96 | CDE | |
| BIO-M3 | PM3-14 | 5,43 | A | 4,65 | C | 4,40 | B | 4,01 | F | 3,95 | E | 3,63 | B | |
| BIO-M3 | PM3-8 | 5,12 | C | 5,05 | A | 4,92 | A | 4,25 | C | 4,22 | B | 4,18 | HIJ | |
| BIO-M3 | R4 | 5,03 | CD | 4,97 | A | 4,88 | A | 4,67 | A | 3,81 | FG | 3,72 | FGHI | |
| PGPRs-4N | CHA0 | 5,04 | CD | 4,61 | C | 4,19 | BCD | 4,08 | E | 3,83 | F | 3,66 | HIJ | |
| PGPRs-4N | BM2-12 | 5,03 | CD | 3,67 | G | 3,54 | GHI | 3,88 | I | 3,24 | K | 3,35 | K | |
| PGPRs-4N | PM3-14 | 4,83 | F | 3,91 | DE | 4,32 | B | 4,08 | E | 3,99 | E | 3,68 | GHIJ | |
| PGPRs-4N | PM3-8 | 4,95 | DE | 3,87 | E | 3,80 | EFG | 4,23 | C | 4,00 | E | 3,98 | CD | |
| PGPRs-4N | R4 | 5,28 | B | 3,88 | E | 4,01 | CDE | 3,84 | J | 3,83 | F | 4,10 | BC | |
| P (formulados) | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | ||||||||
| P (bacterias) | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | ||||||||
| P (interacción) | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | < 0,0001 | ||||||||
Los valores de p indican el efecto de los factores sobre la variable dependiente mediante un análisis de varianza de dos vías (formulado × bacteria). Los valores con letras similares no presentan diferencias estadísticas significativas al nivel de p ≤ 0,05, según el procedimiento de comparación múltiple de Tukey, al tiempo de muestreo (1, 6, 11, 16, 21 y 26 días). / The p values indicate the effect of the factors on the dependent variable using a two-way analysis of variance (formulated × bacterial). Values with similar letters do not present significant statistical differences at the level of p ≤ 0.05, according to Tukey’s multiple comparison procedure, at sampling time (1, 6, 11, 16, 21 and 26 days).
La acidificación continua del medio causa la muerte celular de los microorganismos, por efecto de los ácidos orgánicos que afectan la adecuada estabilidad de pH en el medio. Al día 26, se tomó en consideración que algunos valores de pH se mantienen y otros se reducen de forma sustancial; situación que presentó BIOQ-PGPRs con BM2-12, donde se mantiene el nivel de pH entre (3,73 y 4,43). La reducción del pH está sujeta a los compuestos acidificantes que formaron las bacterias por la disponibilidad de azúcares y proteínas, así como su capacidad de liberar enzimas, como los ácidos orgánicos, que disminuyen los niveles de pH del medio.
Incidencia del bioformulado BIOQ-PGPRs en CCN-51 según la forma de aplicación en temporada seca y lluviosa
Las rizobacterias crecieron en el bioformulado BIOQ-PGPRs. Su aplicación a campo se realizó en consorcio en parcelas de CCN-51, que define valores significativos a la forma de aplicación (edáfica, foliar e inyección), la temporada (lluviosa y seca) y sus interacciones para las variables morfológicas y productivas.
La evaluación de emisión foliar de las aplicaciones de rizobacterias en CCN-51, tuvo un aumento del número de hojas con similar respuesta a su aplicación edáfica e inyección en temporada lluviosa y seca, con 100 y 52, y 108 y 53, respectivamente (Cuadro 7). Hubo un incremento de la masa foliar por las aplicaciones de las rizobacterias, a diferencia de las parcelas de CCN-51 tratadas con control químico y sin aplicación; se redujo la emisión foliar con 73 y 35 en temporada lluviosa y seca, respectivamente (Cuadro 7).
Cuadro 7 Emisiones foliares, número de granos y rendimiento de T. cacao durante la época seca y lluviosa. Ecuador, 2020.
| Forma de aplicación | Época | Número de emisiones foliares | Número de granos | Rendimiento kg/ha | |||||||||
| Edáfico | Lluviosa | 100 | ± | 11,9 | AB | 51 | ± | 2,2 | A | 157,3 | ± | 43,6 | D |
| Edáfico | Seca | 52 | ± | 3,9 | DE | 42,2 | ± | 4,5 | BCD | 1270,6 | ± | 235 | A |
| Foliar | Lluviosa | 97 | ± | 17,9 | AB | 45,2 | ± | 1,4 | ABC | 117,4 | ± | 39,5 | D |
| Foliar | Seca | 61 | ± | 17,2 | CDE | 44,8 | ± | 3,5 | ABC | 486 | ± | 146 | C |
| Inyección | Lluviosa | 108 | ± | 18,2 | A | 46,2 | ± | 3,1 | ABC | 85,6 | ± | 25,9 | D |
| Inyección | Seca | 53 | ± | 10,6 | DE | 36,8 | ± | 3,7 | D | 804 | ± | 129 | B |
| Químico | Lluviosa | 73 | ± | 13,9 | BCD | 46,8 | ± | 3,3 | AB | 94,3 | ± | 15,1 | D |
| Químico | Seca | 54 | ± | 5,9 | DE | 45 | ± | 1,3 | ABC | 723 | ± | 103 | B |
| Testigo | Lluviosa | 84 | ± | 16,6 | ABC | 40,2 | ± | 2,1 | CD | 60,7 | ± | 20,5 | D |
| Testigo | Seca | 35 | ± | 6,5 | E | 36,4 | ± | 3,6 | D | 473 | ± | 63,9 | C |
| P (forma de aplicación) | 0,001 | <0,0001 | <0,0001 | ||||||||||
| P (época) | <0,0001 | 0,0005 | <0,0001 | ||||||||||
| P (interacción ) | 0,0255 | 0,0001 | <0,0001 | ||||||||||
Los valores de p indican el efecto de los factores sobre la variable dependiente mediante un análisis de varianza de dos vías (forma de aplicación × época). Los valores con letras similares no presentan diferencias estadísticas significativas al nivel de p ≤ 0,05, según el procedimiento de comparación múltiple de Tukey. Los valores de desviación estándar ± corresponden a n = 16. / The p values indicate the effect of the factors on the dependent variable using a two-way analysis of variance (application method × time). Values with similar letters do not present signif-icant statistical differences at the level of p ≤ 0.05, according to Tukey’s multiple comparison procedure. Standard deviation ± values correspond to n = 16.
La formación de granos en CCN-51, bajo aplicaciones edáficas de las rizobacterias, generó un promedio anual de 47 semillas/mazorca entre las temporadas (lluviosa y seca). Con similar respuesta, las parcelas de CCN-51 tratadas con fertilizante foliar y fungicidas (químico), obtuvieron un promedio de 46 semillas/mazorca entre las dos temporadas (Cuadro 7). Este resultado evidencia que el proceso de colonización de las rizobacterias en CCN-51 ejerce cambios a nivel fisiológico que permiten asimilar los nutrientes para favorecer la formación de mazorcas.
El rendimiento de CCN-51 disminuyó en la temporada lluviosa, característica de este clon. En esta investigación, la aplicación constante de rizobacterias a nivel edáfico ejerció un incremento de 1270,6 kg/ha en la época seca, y se disminuyó a 157 kg/ha en la época lluviosa. Las parcelas de CCN-51 tratadas con fertilización foliar y fungicidas (químico) alcanzaron 723 kg/ha en la temporada seca. Las aplicaciones de microorganismos potenciaron el mecanismo de protección de este clon y mejoraron el proceso de asimilación de minerales, lo que contribuyó al incremento en la producción de semillas de cacao (Cuadro 7). En la temporada lluviosa, se redujo la producción de mazorcas por efecto de las condiciones climáticas, que disminuyen la formación de flores e incrementan las enfermedades.
Incidencia de enfermedades en mazorcas de CCN-51 según la forma de aplicación del bioformulado en temporada seca y lluviosa
La aplicación del bioformulado BIOQ-PGPRs mostró diferencias significativas en la interacción de formas de aplicación y temporadas (p < 0,0001). Las aplicaciones edáficas del bioformulado BIOQ-PGPRs disminuyeron la incidencia de Phytophthora spp., en mazorcas de cacao, donde las rizobacterias activan el mecanismo de protección a nivel fisiológico en CCN-51 para responder de manera más eficiente a enfermedades. La mayor incidencia de la enfermedad fue evidente en las parcelas de CCN-51 no tratadas con fungicidas químicos con un incremento de infección en mazorcas del 9,7 % (Cuadro 8).
Cuadro 8 Incidencia de Phytophthora spp. y M. roreri durante la época seca y lluviosa. Ecuador, 2020.
| Forma de aplicación | Temporada | Incidencia en mazorca (%) | |||||||
| Phytophthora spp. | M. roreri | ||||||||
| Edáfico | Lluviosa | 3,6 | ± | 0,5 | CD | 10,9 | ± | 1,4 | B |
| Edáfico | Seca | 0,8 | ± | 0,1 | AB | 34,1 | ± | 3 | F |
| Foliar | Lluviosa | 5,4 | ± | 1,1 | D | 13,6 | ± | 1,4 | BC |
| Foliar | Seca | 7,8 | ± | 0,8 | E | 13,6 | ± | 1,4 | BC |
| Inyección | Lluviosa | 8,3 | ± | 1,2 | E | 30,7 | ± | 1,6 | EF |
| Inyección | Seca | 0 | ± | 0 | A | 20,9 | ± | 3,4 | D |
| Químico | Lluviosa | 2,7 | ± | 0,8 | BC | 3,61 | ± | 0,4 | A |
| Químico | Seca | 4,0 | ± | 1,0 | CD | 28,0 | ± | 3,3 | E |
| Control | Lluviosa | 4,0 | ± | 1,1 | CD | 17,7 | ± | 1,8 | CD |
| Control | Seca | 9,7 | ± | 1,5 | E | 42,9 | ± | 3 | G |
| P (forma de aplicación) | <0,0001 | <0,0001 | |||||||
| P (época) | 0,1985 | <0,0001 | |||||||
| P (interacción) | <0,0001 | <0,0001 | |||||||
Los valores de p indican el efecto de los factores sobre la variable dependiente mediante un análisis de varianza de dos vías (forma de aplicación × época). Los valores con letras similares no presentan diferencias estadísticas significativas al nivel de p ≤ 0,05, según el procedimiento de comparación múltiple de Tukey. Los valores de desviación estándar ± corresponden a n = 16. / The p values indicate the effect of the factors on the dependent variable using a two-way analysis of variance (application method × time). Values with similar letters do not present significant statistical differences at the level of p ≤ 0.05, according to Tukey’s multiple comparison procedure. Standard deviation ± values correspond to n = 16.
La proliferación de M. roreri en mazorcas de cacao disminuyó con la aplicación del bioformulado BIOQ-PGPRs a nivel foliar, con niveles de infección del 13,6 % para sus dos temporadas, actuó sobre la mazorca de cacao. La aplicación edáfica del bioformulado PGPRs en temporada seca se incrementó un 34 % de infección en mazorca de CCN-51 (Cuadro 8). Estos resultados sugieren que la efectividad de las diferentes formas de aplicación para el control de Phytophthora spp. y M. roreri puede variar según la temporada del año.
Discusión
Las PGPR en esta investigación presentaron capacidad de producir biopelículas o exopolisacáridos, enzimas quitinasa y ureasa, diferentes tipos de fitohormonas, metabolitos fijadores de hierro (sideróforos) y solubilizadores de minerales del suelo. Las bacterias del género Pseudomonas, Acinetobacter, Serratia, Enterobacter y Aeromonas son productoras de ácidos orgánicos, controlan hongos fitopatógenos por actividad de la enzima quitinasa, disminuyen el estrés y estimulan el crecimiento de plantas hospederas mediante la producción de fitohormonas y enzimas ACC desaminasa, precipitan metales pesados por la producción de biopelículas (Cadirci & Yilmaz, 2023; Haque et al., 2023; Nagrale et al., 2023; Wang et al., 2023). Esta particularidad mejora el estado fisiológico de las especies vegetales, ya sea directa o indirecta, incrementa la productividad de cultivos agrícolas y minimiza la dependencia de los agroquímicos (Mishra et al., 2020).
Se determinó la presencia del gen chiA en las cepas PM3-8, PM3-14, BM2-12 y CHA0, con el producto de amplificación 225 pb, que codifica el dominio catalítico a la producción de quitinasas bacterianas. Se han reportado cepas de los géneros Serratia, Pseudomonas, Acinetobacter y Enterobacter con presencia del gen chiA, como un indicativo para el control de enfermedades en cultivos agrícolas (Cucu et al., 2019; Medina-de la Rosa et al., 2016; Okay & Alshehri, 2020; Verma et al., 2023).
Con el bioformulado BIO-QPGPRs, se pudieron masificar y preservar células bacterianas hasta los 26 días. Sin embargo, la reducción de las UFC día 26 está influenciada por el metabolismo de los microorganismos y la dosis de cada ingrediente; en especial de los azúcares que pueden formar alcoholes, elemento que minimiza la presencia del oxígeno y acelera la muerte celular (Arriel-Elias et al., 2018).
Las fuentes de nutrientes, adherentes y estabilizadores juegan un rol fundamental en la protección y viabilidad de las formulaciones líquidas de bacterias (Lee et al., 2016). Esto indica que el bioformulado tiene componentes activos con la capacidad de preservar células bacterianas por un tiempo determinado. Por tal motivo, es relevante conocer las UFC/mL en los bioformulados, ya que son indicativas de calidad del producto (Lobo et al., 2019).
Los bioformulados en estudio presentaron una disminución del pH del día 1 al 26, con valores inferiores a 5. Las PGPR se desarrollan en medios con melaza a pH 4-9; sin embargo, el pH óptimo para la mayoría de los aislados bacterianos está entre 6,5 y 7 (Mustapha et al., 2022). No obstante, existen reportes que indican que altas concentraciones de azúcares desarrollan una inestabilidad en las características químicas (pH) de las formulaciones líquidas que ocasionan la reducción y muerte bacteriana (Guerra López & Zúñiga Dávila, 2018). Por tal motivo, los componentes utilizados en las formulaciones podrían influir en la disminución celular. En este contexto, las características de tolerancia a las condiciones ácidas del formulado podrían ser una alternativa para usar en suelos con estas mismas condiciones (Ma et al., 2020).
La aplicación a nivel de campo del bioformulado BIO-QPGPRs a manera de consorcio de PGPR, en plantación de T. cacao CCN-51, promovió las emisiones foliares durante la época lluviosa, en su aplicación foliar. La aplicación del bioformulado con P. aeruginosa aumentó el número de brotes en manzanas (Malus domestica L.) en condiciones de campo (Sharma & Sharma, 2022). La aplicación de bioformulados PGPR en Solanum tuberosum mostró mayor biomasa y área foliar en comparación con las plantas sin aplicación (Batool et al., 2020). Las PGPR tienen un efecto en el crecimiento foliar de T. cacao (Loguercio et al., 2023), el cual se origina porque las rizobacterias estimulan una mayor densidad de cloroplastos, clorofila y nutriente por unidad de área foliar, por liberación de hormonas citoquininas.
El incremento en el número de granos presentado en la interacción edáfica y la época seca está regulado por el proceso de disponibilidad de nutrientes, en el cual las rizobacterias juegan un rol sustancial en la capacidad de solubilizar minerales y que estén disponibles para CCN-51, en comparación con el control sin aplicaciones. El bioformulado con P. fluorescens promovió un mayor número y peso de semillas por planta de Brassica juncea en comparación con el control (Mishra et al., 2020). Las PGPR mejoran la solubilización y eficiencia en la absorción de nutrientes que retribuyen en la producción y calidad de semillas (Hakim et al., 2021). Esto demuestra que las aplicaciones edáficas de la bioformulación son eficientes para incrementar el número de semillas en los cultivos.
La forma de aplicación edáfica en la época seca incrementó el rendimiento del cultivo de cacao (1270,6 kg/ha), en comparación con las aplicaciones por inyección y foliar. La aplicación de la formulación de la emulsión de aceite con P. fluorescens FP79, productora de AIA y citoquininas, aumentó significativamente el rendimiento del plátano (68,12 t/ha) en comparación con el control (66,49 t/ha) (Peeran et al., 2014). Se reportó que la aplicación de formulaciones de Pseudomonas y Azospirillum generó un incremento en el rendimiento y la calidad del fruto de tomate (Pérez-Rodriguez et al., 2020). Esta alternativa mejora la eficiencia de los agroquímicos y el rendimiento en el cultivo de cacao.
En cuanto al control fitosanitario de Phytophthora spp. y M. roreri en mazorcas de cacao, se observó una disminución bajo la aplicación del bioformulado BIOQ-PGPRs tanto a nivel foliar como edáfico. Se han reportado estudios en los que la formulación orgánica basada en PGPR ocasionó un efecto biocontrolador por la acción de PGPR, generando un efecto antagónico frente a hongos patógenos en especies vegetales (Chakravarty, 2023; Meena et al., 2020; Pathma et al., 2021).
Existen pocos reportes sobre el empleo de formulaciones PGPR al control biológico de infecciones ocasionadas por hongos en CCN-51. Sin embargo, la alternativa es viable según Waisen et al. (2023) y Anderson y Kim (2020), quienes utilizaron el producto comercial P. chlororaphis AFS009 (Howler™, AgBiome Inc., USA) para la protección en plantas contra los fitopatógenos Alternaria sp., Botrytis sp., Colletotrichum sp., Fusarium sp., Phytophthora sp., Pythium sp., Rhizoctonia sp. y Sclerotinia sp., por la producción de fenazinas antimicrobianas, insecticidas y nematicidas (Anderson y Kim, 2020; Waisen et al., 2023). La producción de compuestos como el 2,4 diacetilfloroglucinol (DAPG), fenazinas, pioluteorina y cianuro de hidrógeno podrían estar ligada a la acción de biocontrol en enfermedades fúngicas (Balmes et al., 2022; Chávez Arteaga et al., 2020). Además, mediante pruebas bioquímicas y moleculares, se encontró que la mayoría de las PGPR son productoras de quitinasas, lo cual es fundamental para el control de hongos con estructura quinolitica, como M. roreri.
Conclusiones
Los bioformulados de rizobacterias presentaron variabilidad en las unidades formadoras de colonias (UFC) y el potencial de hidrógeno (pH) durante el periodo de evaluación, ubicándose el bioformulado BIOQPGPRs con las cepas P. protegens CHA0, M2-12, E. asburiae BM3-14, S. marcescens M3-8 y P. veronii R4 en UFC/mL como el medio con mayor estabilidad con respecto a las variables antes mencionadas, durante los 26 días de evaluación.
En el cultivo de cacao, la aplicación por inyección en la época lluviosa incrementó el número de emisiones foliares (108), mientras que la aplicación edáfica en la misma época aumentó el número de granos (51). En época seca, el rendimiento del peso seco del cacao (1270 kg/ha) mejoró cuando las aplicaciones de los formulados se realizaron de forma edáfica. Además, en la temporada seca, se presentó una disminución total de Phytophthora (0 %) por el efecto de las aplicaciones por inyección.

