











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
Los frijoles (Phaseolus vulgaris L.) son semillas comestibles e importantes a nivel económico, se cultivan y consumen en todo el mundo. El frijol se destaca como fuente de proteína, almidón y fibra dietética, necesaria para las personas en países en desarrollo (Gomes Basso Los et al., 2018). Colombia no logra suplir la demanda interna de leguminosas, por lo que importa de China, Bolivia, Perú, Ecuador y Estados Unidos (Federación Nacional de Cultivadores de Cereales, Leguminosas y Soya de Colombia (FENALCE), 2022). En el año 2022 este país importó 47 324 t de leguminosas, lo cual significó un incremento de 58,70 %, en comparación con el año 2020 (FENALCE, 2022). En cuanto a producción, el departamento del Norte de Santander ocupó el cuarto lugar a nivel nacional con 10 990 t en el año 2013. En el año 2017 la región disminuyó su producción a 4923,78 t en un área de 2646,75 ha (Cámara de Comercio de Bogotá, 2015).
La producción agrícola está sometida a diversas limitaciones. Las arvenses, las plagas, el desequilibrio en la nutrición y los factores abióticos, son las principales amenazas (Tahseen et al., 2015). Las plantas arvenses presentan mayor crecimiento y tolerancia a diferentes factores ambientales y su reproducción es más eficiente (Ibanhes Neto et al., 2020). Las plantas compiten por la luminosidad (Araniti et al., 2015) y pueden sufrir otros efectos perjudiciales, como los alelopáticos de un cultivo sobre el otro (Huang et al., 2015). La presencia de plantas arvenses, entre otros factores fitosanitarios, es la principal limitación del cultivo de frijol, por lo que representan un obstáculo para lograr mayores rendimientos, limitan la producción de grano e incrementan los costos de producción (Pratibha et al., 2021).
La alelopatía involucra fitoquímicos producidos por las plantas con impacto en el crecimiento y el desarrollo de la planta receptora (Hussain et al., 2020). La intensidad y la naturaleza de los fitotóxicos producidos por la planta, afecta el área donde se desarrollan las receptoras (Hussain et al., 2020). Altera la estructura y la elongación celular, cambia su sistema y altera las reacciones enzimáticas de la planta receptora (Ibanhes Neto et al., 2020). Las arvenses liberan al medio ambiente una gran diversidad de metabolitos primarios y secundarios a partir de las hojas, tallo y raíz (Taiz et al., 2017). Se han observado diferencias sobre el efecto alelopático de extractos acuosos de Argemone ochroleuca, según el órgano de la planta utilizado (Mlombo et al., 2021).
Investigaciones han identificado funciones biológicas complejas de los metabolitos secundarios. Los compuestos que se han detectado incluyen una variedad de clases químicas como ácidos fenólicos, cumarinas, benzoquinonas, terpenoides, glucosinolatos y taninos (Putnam & Duke, 1978). Los compuestos fenólicos están presentes en las arvenses (Nunes Lopes et al., 2022; Vargas Martínez et al., 2018) con propiedades alelopáticas, como la lignina (Jaimes-Cruz et al., 2021), que es un metabolito secundario con efecto fitotóxico sobre otras especies (Macías et al., 2019). Cynara cardunculus posee propiedades fitotóxicas, debido a la presencia de un anillo aromático en su composición, con múltiples grupos hidroxilo (Ben Kaab et al., 2020).
Las propiedades fitotóxicas de los compuestos fenólicos destruyen la membrana plasmática y contribuyen al efecto oxidativo (Hasan et al., 2021). Las moléculas modificadas de siloxanos (Burbano Salas, 2019), son utilizadas como herbicidas, por su acción fitotóxica contra las malezas (Undabeytia et al., 2021). Estos y otros aleloquímicos se encuentran en diversas especies, desde leñosas hasta herbáceas, pastos, hojas anchas, arvenses y cultivos (Tahseen et al., 2015). Existen arvenses con capacidad de inhibir la germinación de semillas, la emergencia y el crecimiento de plántulas de frijol (Ibanhes Neto et al., 2020). Investigaciones previas reportaron la disminución de la germinación de P. vulgaris por el efecto alelopático de A. ochroleuca. Los extractos acuosos de hoja, presentaron mayor efecto alelopático, en comparación con el tallo y la raíz (Mlombo et al., 2021).
Es necesario conocer la biología y la ecología de las especies arvenses, para realizar un correcto manejo integral hacia su control (Moreno-Preciado & Balaguera-López, 2021). La gran diversidad de plantas arvenses con efecto alelopático en campos agrícolas, justifica la necesidad de investigación en el área (Flores Córdova et al., 2015). El municipio Pamplona en Norte de Santander, presenta escasos estudios sobre el potencial alelopático de C. clandestinus (Hochst. ex Chiov.), el cual puede interferir en la germinación, crecimiento y desarrollo de P. vulgaris, de allí la importancia del estudio. El objetivo de la presente investigación fue evaluar la alelopatía del pasto kikuyo (C. clandestinus) en la germinación de frijol (P. vulgaris) variedad ICA Cerinza.
Materiales y métodos
Sitio de estudio
La investigación se realizó durante los meses de mayo a agosto de 2022, en el laboratorio de Control de Calidad de la Universidad de Pamplona, Campus Central (7°23’9,41” N, 72°38’59,98” O, 2300 m s. n. m.), Pamplona, Norte de Santander, Colombia.
Población y muestra de estudio
Para evaluar el efecto alelopático de C. clandestinus en la germinación, se utilizaron 432 semillas homogéneas en forma y tamaño de frijol de la variedad ICA Cerinza, seleccionada por su adaptación a las zonas de producción (2000 a 2300 m s. n. m.). Las semillas de frijol fueron lavadas con agua potable y sometidas a desinfección con hipoclorito de sodio (3 % v/v), durante 10 min, seguido de dos enjuagues con agua destilada (Castillo-Quiroz et al., 2018).
Toma de muestras de las plantas de kikuyo
Las plantas de C. clandestinus, se recolectaron en el campo del Centro de Investigación en Sanidad Vegetal y Bioinsumos (CISVEB) de la Universidad de Pamplona, durante el período de floración. Esta arvense se considera invasora en el territorio de Pamplona, según estudios previos del Programa de Ingeniería Agronómica de la Universidad de Pamplona (Labrada, s.f; Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO), 2006). Las plantas de C. clandestinus se identificaron en el Herbario Catatumbo Sarare de la Universidad de Pamplona, Colombia.
Después de la colecta, las plantas se lavaron con agua destilada y se secaron en oscuridad, durante dos semanas. Luego, la materia seca fue separada por órgano (hoja, tallo y raíz) y triturada en un procesador de alimentos para la obtención del polvo (Sithara et al., 2017).
Preparación del extracto
El extracto acuoso se obtuvo a partir de 5 g de polvo de las muestras trituradas (hoja, tallo y raíz) de C. clandestinus, los cuales se agregaron en 100 mL de agua destilada (Arora, 2015), relación masa/volumen (g/mL). La mezcla se homogeneizó en un agitador mecánico a 100 rpm, durante 20 min y luego se dejó reposar durante 24 h a temperatura ambiente (23 ± 2 °C) y oscuridad.
Los extractos se filtraron en papel filtro (Whatman nº 1) mediante una bomba de vacío, capacidad 2,00 m3/h. Los tratamientos se constituyeron en las proporciones 1 %, 2,5 % y 5 %, de extracto de C. clandestinus y uno adicional, compuesto por agua destilada (0 %), para cada uno de los órganos (hoja, tallo y raíz), con un total de doce tratamientos (Cuadro 1). Los extractos acuosos se conservaron en envases de plástico protegido de la luz hasta su uso.
Cuadro 1 Tratamientos generados por la combinación de niveles de los factores órgano de la planta y concentración del extracto acuoso de C. clandestinus, aplicados a las semillas de P. vulgaris, variedad ICA Cerinza. Laboratorio de Control de Calidad de la Universidad de Pamplona, Campus Central, Pamplona, Norte de Santander, Colombia, mayo-agosto 2022.
| Órgano de la planta | Concentración (%) | |||
| Hoja | 0,0 | 1,0 | 2,5 | 5,0 |
| Tallo | 0,0 | 1,0 | 2,5 | 5,0 |
| Raíz | 0,0 | 1,0 | 2,5 | 5,0 |
Obtención del extracto acuoso liofilizado
Para la obtención del extracto acuoso liofilizado, se colocaron 30 mL de extracto de C. clandestinus en bandejas de liofilización y se llevaron a un liofilizador a 40 mbar de presión. El proceso de liofilización consistió en seis segmentos, de acuerdo con el Cuadro 2.
Germinación
La prueba de germinación se realizó para los doce tratamientos (Cuadro 1). Las semillas de frijol se dispusieron en cajas de Petri (12 cm Ø) con papel esterilizado (Whatman No. 1), doce semillas por repetición (Abril-Saltos et al., 2023; Morales-Santos et al., 2017) y tres repeticiones por tratamiento. El papel esterilizado se humedeció con 6 mL del extracto acuoso (Rodríguez-Gutiérrez et al., 2016). El extracto acuoso se aplicó a las cajas de Petri en días alternos, a razón de 6 mL del extracto, durante la evaluación.
Las cajas de Petri con los tratamientos se mantuvieron en condiciones de laboratorio con temperatura de 21 ± 1 °C, 48 ± 2 % de humedad relativa y 24 h de oscuridad. El recuento de la prueba de germinación se ralizó a diario; como semilla germinada se consideró la semilla que presentó 1 cm de raíz primaria. La lectura se realizó hasta que la estabilización de la germinación alcanzó el valor máximo, a los doce días después de la siembra (Hartmann & Kester, 1988).
Para el análisis de la dinámica de germinación se consideraron las variables: número de semillas germinadas (NSG), índice de tasa de germinación (ITG) e índice de germinación de Scott (IG). El NSG se registró como número de semillas germinadas. El ITG representó el tiempo transcurrido en germinar las semillas (Clavijo & Baker, 1988; Quevedo García et al., 2015), calculado según la ecuación 1.
ITG = (G1/T1 + G2/T2 ……+ Gn/Tn) / % G (1)
Donde G1 es el porcentaje de semillas germinadas en T1, G2 el porcentaje de semillas germinadas entre T1 yT2, Gn el porcentaje de semillas germinadas al final, T1 corresponde a los días al primer conteo, T2 los días al segundo conteo, Tn los días al conteo final y % G es el porcentaje total de germinación.
El IG provee una medida del tiempo de germinación en relación con la capacidad de germinación (González-Zertuche & Orozco-Segovia, 1996; Scott et al., 1984), calculado según la ecuación 2.
IG = ∑ (ni * ti) / N (2)
Donde ni es el número de semillas germinadas el i-ésimo día, ti el número de días después de la siembra y N el número total de semillas sembradas.
Obtención de espectros infrarrojos mediante espectroscopía
Los espectros de las muestras secas y el extracto acuoso liofilizado de C. clandestinus (hoja, tallo y raíz), se realizaron en el laboratorio de Biocalorimetría de la Universidad de Pamplona, con la técnica de espectroscopía infrarroja transformada de Fourier (FTIR, por sus siglas en inglés), acoplada con reflectancia total atenuada (ATR, por sus siglas en inglés).
La medición por espectroscopía ATR-FTIR se realizó en un espectrofotómetro FTIR, provisto de un detector DLATGS (sulfato de triglicina dopado con L-alanina deuterada) equipado con un accesorio ATR, con cristal de diamante. Los espectros se obtuvieron en la región del infrarrojo medio, con un barrido de 4000 hasta 500 cm-1, a una resolución de 4 cm-1 y 48 scans. El espectro de cada muestra se analizó por triplicado de manera independiente.
Diseño y análisis estadístico
Se utilizó un arreglo factorial (3x4) en un diseño totalmente al azar, con medidas repetidas en la unidad experimental, con enfoque univariado, donde los factores intersujetos fueron el órgano de la planta (A3= hoja, tallo y raíz) y la concentración del extracto acuoso de C. clandestinus (B4= 0 %, 1 %, 2,5 % y 5 %), mientras que el factor intrasujeto fue el tiempo o días después del inicio del experimento (doce mediciones).
Para el análisis estadístico de la variable número de semillas germinadas, se realizó un análisis exploratorio de la normalidad, mediante la prueba de Shapiro-Wilk y homogeneidad de varianzas, con la prueba de levene, encontrándose que la variable NSG no siguió una distribución normal, por ello, se utilizó la transformación √x+1 y se aplicó luego el análisis de la varianza (ANDEVA) de medidas repetidas. Las medias se compararon por el test de Bonferroni, al 5 %. Las variables ITG e IG, cumplieron con los supuestos de normalidad y homogeneidad de varianzas, por lo que se realizó el ANDEVA en el diseño completamente aleatorio con arreglo factorial (3x4), con los factores órgano de la planta, concentración y su interacción. En las comparaciones de medias de los índices, se utilizó la prueba de comparaciones múltiples de Tukey al 5 %, con el programa estadístico SPSS (International Business Machine, 2015).
Resultados
Número de semillas germinadas (NSG)
Las estadísticas descriptivas del número de semillas germinadas (NSG) de frijol en las diferentes concentraciones de extracto acuoso de C. clandestinus, se presentan en el Cuadro 3. La mayor media en el NSG se observó para las tres concentraciones del extracto acuoso en comparación con el testigo que presentó un valor de 1,27.
Cuadro 3 Estadísticas descriptivas del número de semillas germinadas (NSG) de Phaseolus vulgaris variedad ICA Cerinza, en las diferentes concentraciones de extracto acuoso de Cenchrus clandestinus. Laboratorio de Control de Calidad de la Universidad de Pamplona, Campus Central, Pamplona, Norte de Santander, Colombia. Mayo-agosto 2022.
| Concentración (%) | Media/SE | Límite inferior | Límite superior | Rango |
| 0 | 1,27±0,004 | 1,26 | 1,28 | 0,02 |
| 1 | 1,30±0,004 | 1,29 | 1,31 | 0,02 |
| 2,5 | 1,29±0,004 | 1,29 | 1,30 | 0,01 |
| 5 | 1,30±0,004 | 1,29 | 1,31 | 0,02 |
* Intervalo de confianza 95 %. SE: error estándar. / 95 % confidence interval. SE: Standard error.
Cuando se evaluó la fitotoxicidad de los extractos acuosos de C. clandestinus en la germinación de frijol, en el análisis de varianza se encontraron diferencias estadísticas significativas (p<0,001) en el NSG entre los tratamientos y el testigo, por efecto de la interacción tiempo después de la siembra por concentración del extracto acuoso, pero no para el órgano de la planta.
La germinación se inició con la imbibición de frijol, a partir del segundo día de evaluación, con la emergencia de la radícula, y se incrementó hasta el día cinco, a partir del cual empezó a descender (Figura 1). La media más alta en el NSG se registró en los días tres y cinco del experimento (3,75 y 1,03, respectivamente), con una tendencia cuadrática, donde se cruzaron dichas líneas, lo que identificó el efecto de la interacción entre el tiempo (días después de la siembra) y la concentración del extracto acuoso. El mayor NSG de frijol con los extractos acuosos de C. clandestinus se registró el día 4, en el orden de concentración 1, 5 y 2,5 % (Figura 1), mientras que para el testigo fue el día 3.
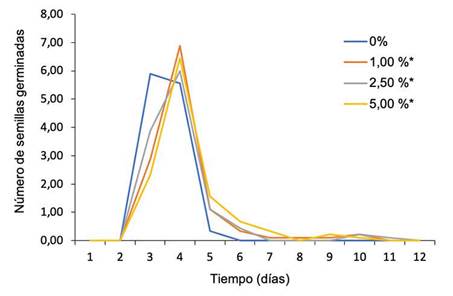
Figura 1 Perfil del tiempo y el número de semillas germinadas (NSG) de Phaseolus vulgaris en las diferentes concentraciones de extracto acuoso de kikuyo (Cenchrus clandestinus). Laboratorio de Control de Calidad de la Universidad de Pamplona, Campus Central, Pamplona, Norte de Santander, Colombia. Mayo-agosto 2022.
Índices de germinación
En relación con los índices de germinación evaluados, el análisis de la varianza detectó diferencias para el ITG por efecto de los factores órgano (p<0,05) y concentración (p<0,001) y en el IG para la concentración (p<0,001).
Índice de tasa de germinación (ITG)
El aumento de la concentración del extracto acuoso de C. clandestinus disminuyó el ITG en frijol, con interacción entre todos los factores evaluados (Figura 2).
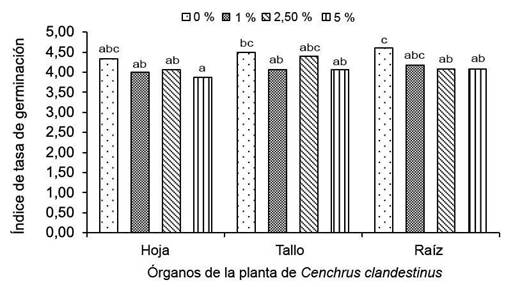
Figura 2 Índice de tasa de germinación (ITG) de semillas de Phaseolus vulgaris tratadas con extracto acuoso de Cenchrus clandestinus (hoja, tallo y raíz) en diferentes concentraciones (0 %, 1 %, 2,5 % y 5 %). Laboratorio de Control de Calidad de la Universidad de Pamplona, Campus Central, Pamplona, Norte de Santander, Colombia. Mayo-agosto 2022.
La mayor velocidad de germinación se observó para las semillas del control (4,61), la cual disminuyó con el aumento de la concentración de los tres extractos acuosos en comparación con el testigo. La aplicación de extracto acuoso del tallo de C. clandestinus al 2,5 %, fue la que menos afecto el ITG (4,40), pero sin exceder al testigo. El mayor y menor ITG se observó con 1 % y 5 % de concentración del extracto de hoja (4,00 % y 3,87 %, respectivamente), tallo (4,07 % y 4,06 %, respectivamente) y raíz (4,18 % y 4,08 %, respectivamente). La menor media en el ITG fue para las semillas de frijol tratadas con el extracto acuoso de hoja al 5 % (3,87).
Índice de germinación (IG)
La mayor media en el IG (Figura 3), fue para las semillas de frijol tratadas con el extracto acuoso de C. clandestinus al 5 % (4,30), lo que indica que la semilla requirió mayor número de días promedio para alcanzar la máxima germinación, en comparación con el testigo que fue menor (3,39). Para el extracto acuoso de hoja al 1 %, se registró mayor valor (4,17) de IG en comparación con la concentración al 2,5 %, que fue 4,14.
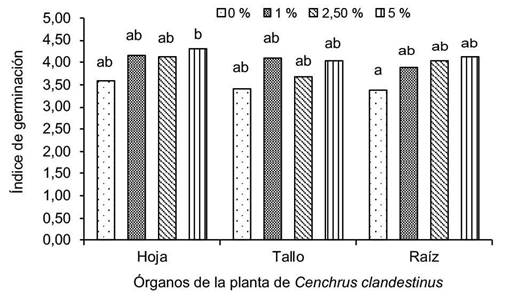
Figura 3 Índice de germinación (IG) de semillas de Phaseolus vulgaris tratadas con extracto acuoso de Cenchrus clandestinus (hoja, tallo y raíz) en diferentes concentraciones (0 %, 1 %, 2,5 % y 5 %). Laboratorio de Control de Calidad de la Universidad de Pamplona, Campus Central, Pamplona, Norte de Santander, Colombia. Mayo-agosto 2022.
El menor IG de semillas de frijol tratadas con los extractos acuosos de tallo y raíz de C. clandestinus, fue para el testigo pero el resultado no difirió de las concentraciones evaluadas (1 %, 2,5 % y 5 %).
Análisis de FTIR y figura del espectro de C. clandestinus
Los datos del espectro ATR-FTIR se obtuvieron en la región infrarroja entre 4000 y 500 cm-1. La Figura 4 muestra los espectros obtenidos a partir del análisis de los extractos de las muestras de hoja, tallo y raíz de C. clandestinus. Cada una de ellas está representada por la absorbancia versus el número de onda (cm-1), en los cuales se identificaron las bandas más significativas, que representan las vibraciones de los grupos funcionales presentes en las biomoléculas de esta planta.
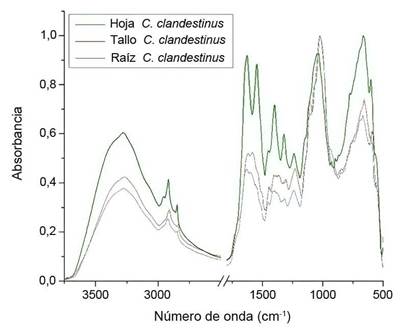
Figura 4 Espectro infrarrojo obtenido por ATR-FTIR para hoja, tallo y raíz de Cenchrus clandestinus en la región espectral de 4000 a 500 cm-1. Laboratorio de Biocalorimetría de la Universidad de Pamplona, Norte de Santander, Colombia. Mayo-agosto 2022.
El C. clandestinus es una arvense perenne baja, que en el análisis ATR-FTIR proporcionó un espectro con bandas características de compuestos lipídicos en la banda de 2913 cm-1 (Figura 4, Cuadro 4), luego en la banda de muy baja intensidad a 2326 cm-1, indicó la presencia de nitrilos especialmente en las muestras de tallo y hoja, ya que en la raíz no se presentó dicha banda.
Cuadro 4 Caracterización de bandas de absorción a partir de los espectros infrarrojos con reflectancia total atenuada en muestras de Kikuyo (Cenchrus clandestinus), en la región de 4000 a 500 cm-1. Laboratorio de Biocalorimetría de la Universidad de Pamplona, Norte de Santander, Colombia. Mayo-agosto 2022.
| Número de onda (cm-1) | Asignación |
| 3400 | Estiramiento O-H polifenoles |
| 2921 | Estiramiento simétrico CH3 |
| 2840 | Estiramiento asimétrico CH2 |
| 2320 | Estiramiento R-CºN común de nitrilos |
| 1640 | Tensión NH3 de aminoácidos |
| 1550 | Flexión NH o estiramiento CN en proteínas |
| 1400 | Tensión CH3 lípidos y aminoácidos |
| 1330 | Tensión C-O ácidos grasos |
| 1241 | Tensión CO éter |
| 1031 | Tensión Si-O-Si en siloxanos |
En el rango espectral de 1700-1500 cm-1 se registró la banda referente a la proteína contenida por la arvense. En 1640 cm-1, se situó la banda que denota vibraciones de tensión NH3, característico en los aminoácidos; también hubo presencia de éteres en la banda 1240 cm-1, mientras que las bandas presentes en la región de 1000-1100 cm-1 fueron representativas de los enlaces Si-O-Si, propios en moléculas de siloxanos. En la banda ancha e intensa alrededor de 3400 cm-1 (Figura 4, Cuadro 4), se asignaron vibraciones de estiramiento O-H de alcoholes que pueden presentarse en compuestos fenólicos.
Discusión
La semilla de frijol presentó diferencias en la germinación con los extractos acuosos de C. clandestinus. El incremento en la concentración de los extractos acuosos redujo el número de semillas germinadas (NSG) de frijol (Lessa et al., 2017). El extracto acuoso de Amburana cearensis y Plectranthus barbatus disminuyó la germinación de Amaranthus deflexus, 51,5 y 99,3 %, respectivamente (Lessa et al., 2017). El efecto alelopático de Sphaeranthus suaveolens en la germinación de semillas y el crecimiento de plántulas de P. vulgaris, fue reportado. El efecto alelopático de diferentes concentraciones del extracto pudo resultar en las diferentes anomalías del metabolismo (Laizer et al., 2021). Estudios previos señalaron el efecto inhibitorio de extractos acuosos de Parthenium hysterophorus, debido a la presencia de algunos aleloquímicos (Tahseen et al., 2015).
La velocidad de germinación disminuyó con el aumento de la concentración de los extractos acuosos en comparación con el testigo y la alelopatía ocurrió en función de la concentración del producto en el medio (de Araujo Barbosa et al., 2018; Tahseen et al., 2015). La disminución pudo estar relacionada con el efecto del extracto acuoso sobre la permeabilidad y selectividad de las membranas celulares de las semillas de frijol (Abugre et al., 2011; Carillo et al., 2010). En P. vulgaris se reportó una reducción en la velocidad de germinación con el aumento de extracto acuoso de hoja de Brassica napus (Rigon et al., 2012). La germinación fue menos sensible a los efectos aleloquímicos cuando se comparó con la velocidad de germinación (Rigon et al., 2012). Los extractos de hoja exhibieron efectos alelopáticos más altos sobre la velocidad de germinación, en comparación con el tallo y la raíz (Mlombo et al., 2021). El efecto de los extractos acuosos depende de la concentración, del órgano utilizado y del estadio fenológico de la planta (Lessa et al., 2017), y las plantas de C. clandestinus se cosecharon durante floración. El extracto acuoso de las flores de Parthenium hysterophorus ocasionaron mayor inhibición de la germinación de P. vulgaris, debido a la presencia de aleloquímicos (Tahseen et al., 2015). La adición de 50 % de extractos acuosos de raíz de Digitaria insularis, solo redujeron 12,50 % la germinación de P. vulgaris (Ibanhes Neto et al., 2020). El mayor índice de germinación de las semillas de frijol con el incremento de la concentración de los extractos acuosos de C. clandestinus, indicó que hubo necesidad de mayor tiempo para alcanzar el máximo incremento de la germinación. La modificación de los patrones de germinación de las semillas de frijol, pudo ser debida a los aleloquímicos presentes en los extractos, aun cuando el número de semillas germinadas durante la evaluación no fue afectado (Santana et al., 2006). Las diferencias mostraron que los índices de tasa de germinación y de germinación son marcadores más sensibles a los aleloquímicos que el número de semillas germinadas.
El análisis ATR-FTIR proporcionó un espectro con bandas características de compuestos nitrilos, solo en las muestras de tallo y hoja de C. clandestinus (Blažević et al., 2019), con efecto alelopático sobre la germinación de las semillas (Brown & Morra, 1995). Los nitrilos son algunos de los productos bioactivos (Frisch et al., 2015; Mocniak et al., 2020) que se originan a través de la hidrólisis de los glucosinolatos, por las enzimas mirosinasas (Frisch et al., 2015). Esto puede suceder en el tejido dañado o tras la reorganización de los compartimentos celulares en el tejido intacto, donde la hidrólisis de glucosinolatos no ocurre, debido a la compartimentación en la célula o por factores aún no identificados (Lv et al., 2022).
La técnica FTIR permite determinar los grupos funcionales presentes en los diferentes fitoquímicos de las muestras de plantas (Kumar Patle et al., 2020). La mayor intensidad de las bandas en el espectro de C. clandestinus, puede relacionarse cuantitativamente con la presencia de fitoquímicos (Beć et al., 2020). La mayor intensidad de las bandas en la muestra de hoja de C. clandestinus, se asoció al mayor efecto alelopático (Beć et al., 2020; Laizer et al., 2021). Mientras que la menor fitotoxicidad de la raíz, coincidió con la menor intensidad de sus bandas de absorción (Beć et al., 2020). Condición de la raíz que se asoció a la menor presencia de los compuestos fitoquímicos determinados en la muestra de estudio (Smith, 2002).
En el presente trabajo se demostró que C. clandestinus es una fuente potencial de aleloquímicos que influyen en la germinación del frijol. El NSG, ITG e IG se vieron afectados en semillas de frijol, germinadas en presencia de extractos acuosos de C. clandestinus (Pérez-Peralta et al., 2019). Los ácidos fenólicos determinados en los extractos acuosos son tipos comunes de aleloquímicos (Shankar et al., 2009), los cuales interrumpen los procesos fisiológicos de la planta receptora, inhiben la actividad enzimática necesaria durante la fotosíntesis, la respiración, el balance hídrico y hormonal de la planta receptora (Soltys et al., 2013). Además, puede alterar importantes vías metabólicas del proceso de germinación y el desarrollo temprano de las plántulas (de Araujo Barbosa et al., 2018).
Conclusiones
El efecto alelopático del tallo y la raíz de C. clandestinus sobre el número de semillas germinadas de frijol, fue menor en comparación con el extracto de hoja. En contraposición, las semillas de frijol fueron afectadas por los extractos de la hoja de C. clandestinus.
El índice de tasa de germinación (ITG) indicó una reducción de la velocidad de germinación con el aumento de la concentración del extracto, con mayor valor para el extracto acuoso de hoja de C. clandestinus al 5 %. Mientras que el índice de germinación (IG) se asoció al mayor número promedio de días para alcanzar el máximo incremento de la germinación de las semillas de frijol, conforme la concentración del extracto acuoso aumentó.
El espectro infrarrojo del extracto reflejó la presencia de polifenoles, nitrilos y xilosanos, de acuerdo con las bandas observadas. El espectro de la hoja de C. clandestinus, presentó las mayores intensidades en sus bandas en comparación con el tallo y la raíz.

