










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgronomía Mesoamericana
On-line version ISSN 2215-3608Print version ISSN 1659-1321
Agron. Mesoam vol.25 n.1 San Pedro Jan./Jun. 2014
Cruzamiento natural en frijol común en Costa Rica
Natural outcrossing in common bean in Costa Rica
Natural outcrossing in common bean in Costa Rica
*Dirección para correspondencia:
Resumen
El objetivo de esta investigación fue estimar el porcentaje de alogamia entre dos variedades de frijol común en Fraijanes y Alajuela, localidades del Valle Central de Costa Rica. La parcela experimental consistió en tres bloques de diez lomillos de 15 m de largo espaciados 0,8 m. En el bloque central se ubicó la variedad de grano color negro, UCR 55 en Fraijanes y Guaymí en Alajuela, y en los laterales la variedad de grano color blanco PAN 68. Los ensayos se llevaron a cabo del 2002 al 2003 en Fraijanes y del 2002 al 2007 en Alajuela, y se evaluó el efecto de la dirección predominante del viento, la estación (seca y lluviosa) y la distancia de la fuente de polen. En ambas localidades se presentó sincronía floral de las variedades de frijol. Se cosecharon y numeraron todas las plantas de PAN 68 en las cinco hileras próximas al bloque central. El cruzamiento natural se estimó como el porcentaje de plántulas con hipocotilo morado en la progenie F1 de la variedad PAN 68, con hipocotilo verde. En Alajuela el cruzamiento natural varió de 0,000 a 0,096% y en Fraijanes de 0,004 a 0,033%. Se obtuvo una alogamia general de 0,031%, baja en comparación con estudios realizados en otros países. La polinización cruzada natural no fue afectada por la condición de clima (seca o lluviosa) o la dirección predominante del viento, pero fue afectada inversamente por la distancia desde la fuente de polen en Alajuela (r2=0,9622, p≤0,05), donde se acercó a cero después de 4 m.
Palabras clave: Phaseolus vulgaris, biología floral, flujo de genes, híbridos naturales, marcador morfológico, polen, polinizadores.
Abstract
The objective of the research was to evaluate the natural outcrossing rate between two varieties of common bean in Alajuela and Fraijanes, localities in the Central Valley of Costa Rica. The experimental plot included three blocks with ten 15.0 m long rows, 0.8 m The black-seeded varieties UCR 55 and Guaymí were growing in Fraijanes and Alajuela in the central block respectively; while the white-seeded variety PAN 68 was planted in the two lateral blocks. The experiments were carried out from 2002 thru 2003 in Fraijanes and from 2002 thru 2007 in Alajuela. The variables: wind direction, seasonal effect and distance from the pollen source were evaluated. There was a synchrony in the flowering time of the varieties in both localities. All plants of the white -seeded variety were numbered and harvested in the five nearest rows to the black-seeded varieties. Natural outcrossing was recorded as a percentage of seedlings with purple hypocotyl in the F1 progeny of PAN 68 (variety that shows green hypocotyls). In Alajuela the rate of natural outcrossing varied from 0,000 to 0.096%, while in Fraijanes it did from 0.004 to 0.033%. The general allogamy was of 0.031%, a lower outcrossing if compared to trials carried out in other countries. The natural outcrossing was not affected by the dry or rainy season nor the wind direction, but it was inversely affected by the distance from the pollen source in Alajuela (r2=0.9622, p≤0.05); It was nil from 4 m away from the pollen source.
Keywords: Phaseolus vulgaris, floral biology, gene flow, natural hybrids, morphological marker, pollen, pollinators.
Introducción
El frijol común (Phaseolus vulgaris L.) es predominantemente autógamo, con una incidencia de polinización cruzada natural muy baja (Bliss, 1980; León, 1987). Su estructura floral protege a los órganos masculinos y femeninos con los pétalos unidos por la quilla, y los granos de polen empiezan a caer sobre el estigma antes de la antesis, lo que le confiere al frijol común la característica de autopolinización (Webster et al., 1977; Ferreira et al., 2007). Sin embargo, la presencia de un nectario activo entre el tubo estaminal y el ovario puede atraer insectos, lo que da lugar a diferentes tasas de polinización cruzada natural en este cultivo (Webster et al., 1982).
La tasa de alogamia está influenciada por muchos aspectos, entre ellos el genotipo y la metodología utilizada para estimarla (Wells et al., 1988; Ramírez et al., 2000; Ferreira et al., 2007), el hábito de crecimiento (Brunner y Beaver, 1989; Ramírez et al., 2000) y la estructura floral de los genotipos empleados (Miranda, 1971; Ramírez et al., 2000). También, las condiciones ecológicas y climáticas específicas de los sitios de evaluación afectan la cantidad y diversidad de polinizadores (Ramírez et al., 2000; Ferreira et al., 2007). Otro factor que influye en flujo de genes en frijol es si el cruzamiento natural se da entre formas cultivadas, o entre estas y las formas silvestres, y en el caso de estas últimas también es importante su origen (Triana et al., 1993).
El cruzamiento natural entre cultivares de frijol común se ha estudiado en países como México, Puerto Rico, Estados Unidos, Brasil, Etiopía, España, Venezuela y Costa Rica. Los valores extremos de máxima alogamia se obtuvieron en trabajos realizados en Estados Unidos, donde se obtuvo el valor más bajo: 0,007% (Tucker y Harding, 1975) y el más alto 66,8% (Wells et al., 1988). En contraste, con excepción de Puerto Rico, donde Brunner y Beaver (1989) obtuvieron 39,3%; los porcentajes máximos de polinización cruzada natural fueron menores: 4,19; 4,50 y 7,45% en México (Crispín, 1960; Miranda, 1971; Ramírez et al., 2000); 4,80% en Etiopía (Stoetzer, 1984); 0,14% y 1,66% en Brasil (Royer et al., 2002; Ferreira et al., 2007); 0,73 y 6,90% en Estados Unidos (Mackie y Smith, 1935; Ibarra et al., 1997); 0,74% en España (Ferreira et al., 2000) y 0,49% en Venezuela (Ortega, 1974). En Costa Rica, el cruzamiento natural entre formas cultivadas de frijol común fue estudiado por Alan y Moh en 1966 y por Pinchinat en 1967, los primeros determinaron una alogamia máxima de 0,20% en Alajuela, mientras que en la segunda investigación se obtuvo 0,10% de cruzamiento natural.
En estudios sobre el flujo de genes entre accesiones silvestres de frijol, se encontró una alogamia promedio de 1,5% en Cerrito, Valle, Colombia (Triana et al., 1993), y entre formas silvestres y cultivadas de 20 a 55% en la Sierra Norte de Puebla, México (Gepts et al., 2000). En el caso de Gepts et al. (2000), estos determinaron el porcentaje de alogamia mediante técnicas moleculares y midieron los eventos de cruzamiento natural acumulados durante años, por lo que obtuvieron porcentajes de esa magnitud. En Costa Rica, Chaves et al. (2009) determinaron una alogamia máxima de 0,199% entre formas silvestres y cultivadas de frijol común.
Los cultivos y sus parientes silvestres pueden cruzarse libremente, lo que da origen a progenies fértiles y viables (Harlan y De Wet, 1971). En relación con el entrecruzamiento, en frijol común se presentan dos situaciones a considerar, por un lado que el flujo de genes es mayor de las formas cultivadas a las silvestres (Gepts y Papa, 2003; Papa y Gepts, 2003), y por otro lado, a pesar del éxito limitado en la transformación genética del frijol (Broughton et al., 2003) y de que la tecnología aún se encuentra en fase experimental, ya se han producido plantas transgénicas (Aragão et al., 2002; Bonfim et al., 2007; Aragão y Faria, 2009; Kwapata et al., 2012). Tanto la presencia de cultivares mejorados como la presencia potencial de transgénicos cerca de poblaciones silvestres de frijol en los centros de origen, representa un riesgo ambiental (Hails, 2000; Hails, 2002), porque se desconoce el efecto que pueda tener la transferencia accidental de genes en la capacidad de adaptación de las formas silvestres.
El impacto de la alogamia sobre la industria de semillas también es un factor importante de considerar. Aunque en cultivos de autopolinización los niveles de cruzamiento son muy bajos, y no deberían existir problemas para la producción de semilla genéticamente pura (Ferreira et al., 2007), se deben conocer los niveles de flujo de genes entre formas cultivadas y las distancias mínimas requeridas para cumplir con los requisitos de pureza en la certificación de semilla. En Costa Rica, la Oficina Nacional de Semillas (ONS) vela por el cumplimiento del “Reglamento técnico para la producción de semilla certificada de frijol”, donde se indica el aislamiento mínimo exigido para la producción de las distintas categorías de semilla (ONS, 2012); sin embargo, no existen estudios de campo que validen las distancias ahí establecidas. Con base en estos antecedentes, se desarrolló el presente trabajo, cuyo objetivo fue determinar el porcentaje de alogamia entre dos variedades de frijol común en Alajuela y Fraijanes, localidades del Valle Central de Costa Rica.
Materiales y métodos
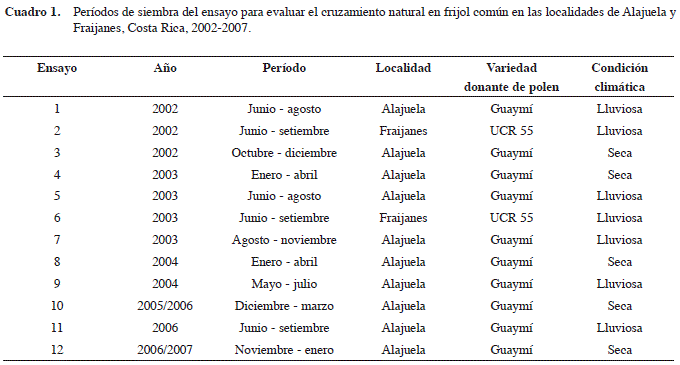
La investigación se llevó a cabo en Costa Rica, entre junio del 2002 y enero del 2007 en la Estación Experimental Agrícola Fabio Baudrit Moreno (EEAFBM) ubicada en San José, Alajuela (10º01’N; 84º16’O; 840 msnm), y entre junio del 2002 y setiembre del 2003 en la Finca Experimental Fraijanes (FEF), ubicada en Fraijanes, Alajuela (10º05’N; 84º16’O; 1764 msnm); ambas pertenecientes a la Universidad de Costa Rica. El ensayo se sembró en la EEAFBM durante diez períodos, cinco en época lluviosa y cinco en época seca; mientras que en la FEF, se sembró durante dos períodos, ambos en época lluviosa (Cuadro 1). El ensayo realizado de octubre a diciembre del 2002 en la EEAFBM, a pesar de ser sembrado en época lluviosa, se consideró de época seca porque fue la condición imperante desde antes de la floración y durante el resto del ciclo de cultivo.
En la EEAFBM, localizada en la zona de vida bosque húmedo premontano con un clima subhúmedo caliente (Holdridge, 1982), la época lluviosa se extiende aproximadamente de mayo a mediados de diciembre y la seca de mediados de diciembre a abril, donde los meses de setiembre y octubre son los más lluviosos (>300 mm mensuales) y enero el más seco (<10 mm mensuales). La precipitación promedio anual es de 1940 mm, que se reduce significativamente en años con influencia del fenómeno de “El Niño”. Durante la época lluviosa, la evaporación promedio mensual presenta valores inferiores a 150 mm, la radiación solar incidente valores inferiores a 17 MJ/día y la humedad relativa promedio es mayor al 85%; mientras que, en la época seca la evaporación promedio mensual llega hasta los 250 mm mensuales, la radiación solar alcanza valores superiores a 19 MJ/día y la humedad relativa promedio es menor al 70%. La temperatura promedio para la EEAFBM es de 21,9°C, con mínimas y máximas promedio de 17,8 y 28,5°C, respectivamente. Los meses más frescos corresponden a diciembre y enero, y los más calientes a marzo y abril (Gutiérrez et al., 1997).
En la FEF, localizada en la zona de vida bosque húmedo montano bajo con un clima (Holdridge, 1982), la época lluviosa se extiende de mayo a noviembre y la seca de enero a marzo; abril y diciembre son considerados meses de transición. Los meses de setiembre y octubre son los más lluviosos (~ 450 mm mensuales) y de enero a marzo los más secos (<100 mm mensuales). La precipitación promedio anual es de 3150 mm. Durante la época lluviosa, la radiación solar presenta valores menores a 12 MJ/día y la humedad relativa promedio es cercana a 90%; mientras que, en la época seca la radiación solar alcanza valores superiores a 16 MJ/ día, la humedad relativa promedio ronda el 80% y la evaporación promedio mensual fluctúa entre 135 y 180 mm mensuales. La temperatura promedio para la FEF es de 16,1°C, con mínimas y máximas promedio de 12,9 y 21,4°C respectivamente. Los meses más frescos corresponden a diciembre y enero, y los más calientes a marzo y abril (Gutiérrez et al., 1999-2002).
Se utilizó el color del hipocotilo como marcador morfológico para identificar las progenies originadas del cruzamiento entre dos variedades de frijol contrastantes para este caracter. El cruzamiento se evaluó en forma unidireccional de una variedad de grano negro con el caracter dominante para el color de hipocotilo (morado) a una variedad de grano blanco con el carácter recesivo para el color de hipocotilo (verde). En Alajuela se utilizó como donante de polen la variedad comercial Guaymí y en Fraijanes la variedad UCR 55 (Cuadro 2), debido a su adaptación diferencial a suelos de origen volcánico con baja disponibilidad de fósforo como los de la FEF (Araya y Acosta, 2000). En ambas localidades se utilizó la variedad PAN 68 como variedad receptora de polen y se evaluó el porcentaje de alogamia en las semillas que ella produjo.
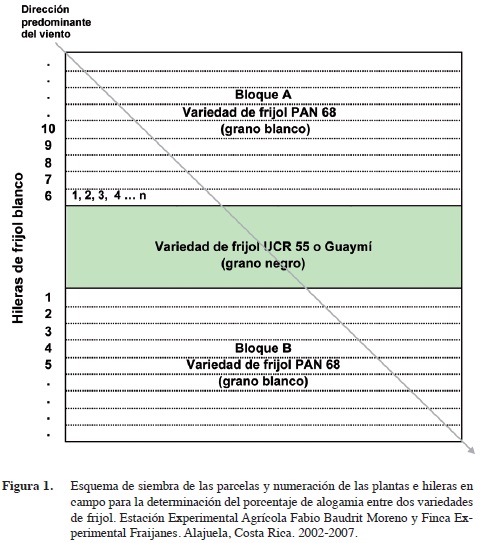
La parcela experimental consistió en tres bloques de diez lomillos de 15 m de largo espaciados 0,8 m. En el bloque central se ubicó el frijol negro (UCR 55 o Guaymí) y en los laterales el frijol blanco (PAN 68) (Figura 1). Para la siembra de las parcelas del ensayo se utilizó una sembradora manual que coloca 10-12 semillas por metro, para una densidad aproximada de 200 000 plantas/ha. El manejo del cultivo se llevó a cabo de acuerdo a las normas de producción comercial. A la siembra se aplicó fertilizante 10-30-10 (150kg/ ha) e insecticida granulado clorpirifós. Las arvenses se controlaron con aplicaciones de paraquat y glifosato, las plagas insectiles con deltametrina e imidacloprid, las enfermedades fungosas con benomil y azoxistrobina, y la fertilización al suelo se reforzó foliarmente con un multimineral (9,1% N; 6,6% P; 5,0% K; 1250 ppm S, 332 ppm B; 17 ppm Co, 664 ppm Zn; 332 ppm Cu; 42 ppm Mo; 207 ppm Ca; 332 ppm Mn; 415 ppm Fe; 207 ppm Mg). Las parcelas del ensayo siempre se ubicaron en un lugar completamente aislado para evitar contaminación con polen proveniente de otras siembras de frijol cercanas, y se realizaron en terrenos provenientes de una rotación con maíz o en descanso, para evitar la aparición de plantas voluntarias de frijol.
Durante la realización de los ensayos se llevó registro de las fechas de inicio y final de las principales etapas fenológicas del cultivo. Durante la floración se evitó la aplicación de insecticidas y se observó cualitativamente la actividad (visita a flores) y tipo de polinizadores que visitaron la parcela. También se obtuvo información de la dirección del viento predominante y las condiciones climáticas imperantes durante la realización del ensayo (temperatura promedio, humedad relativa y precipitación) a través del Instituto Meteorológico Nacional de Costa Rica.
Solamente se cosecharon las plantas de las primeras cinco hileras de cada bloque de la variedad blanca (PAN 68) y el resto fungieron como borde. Se numeraron las hileras de uno a cinco en el bloque B (de acuerdo con la dirección predominante del viento, este atravesó primero la fuente de polen – variedad UCR 55 o Guaymí – y luego pasó por la variedad blanca) y de seis a diez en el bloque A (de acuerdo con la dirección predominante del viento, este atravesó primero la variedad blanca y luego la fuente de polen). Las hileras uno y seis correspondieron a las más cercanas al bloque de frijol negro. A su vez, se numeraron todas las plantas de cada hilera de uno hasta n (Figura 1) y se identificaron con una cinta adhesiva blanca, donde se indicó su posición dentro de la hilera y número de esta. Con base en la numeración efectuada en campo, se procedió a determinar la ubicación aproximada dentro de la parcela del ensayo de cada planta donde pudo ocurrir un evento de cruzamiento y de la progenie resultante.
Las plantas numeradas se trasladaron a un área bajo techo para completar el secado. Una vez secas, se desgranaron manual e individualmente, y el número de semilla se obtuvo con el contador automático Multi Auto Counter, Model DC1, No. 03031, Everwell Corporation. Las semillas se almacenaron en sobres de papel cerrados e identificados. Luego, se pusieron a germinar en bandejas plásticas a razón de una semilla por celda, en una casa de mallas. Cada grupo de semillas se identificó por medio de una paleta plástica con el número de planta e hilera correspondientes. Como sustrato, se utilizó suelo mezclado con granza (cascarilla) de arroz (proporción de 3:1) durante el primer año y peat moss (Plug Mix Perlite VJ PMP 4CF) para el resto del estudio. El sustrato se trató con benomil (80 g de i.a./16 l) adicionado con bomba de espalda para prevenir la incidencia de patógenos. Entre los siete y nueve días después de la siembra, cuando las plántulas alcanzaron aproximadamente ocho centímetros de altura, se evaluó el color del hipocotilo y la venación de las hojas cotiledonares, para determinar si presentaban cruzamiento o no.
Tanto en Alajuela como en Fraijanes, el cruzamiento natural (alogamia) se determinó al obtener el porcentaje de plántulas que germinaron con hipocotilo morado con respecto al total de plántulas germinadas en la F1 de la variedad PAN 68 con grano blanco e hipocotilo verde, de la misma forma en que se estimó en otras investigaciones sobre hibridación natural en frijol (Alan y Moh, 1966; Ortega, 1974; Ramírez et al., 2000; Royer et al., 2002). Mediante una prueba de t se procedió a determinar la influencia de la condición climática sobre la tasa de cruzamiento natural en Alajuela. Para determinar el efecto de la dirección del viento sobre la tasa de alogamia, los datos se analizaron como bloques completos al azar, donde la época de siembra se consideró como el factor de bloqueo. En Alajuela, primero se analizó el efecto de la dirección del viento para cada condición climática (seca o lluviosa) y luego en forma combinada. Además, se consolidaron los datos para cada distancia (posición de las hileras) respecto a la fuente de polen para los diez ensayos en Alajuela y los dos en Fraijanes, y se hizo un análisis de regresión con el fin de determinar la influencia de la distancia de la fuente de polen sobre el porcentaje de cruzamiento natural y la ecuación de la curva de mejor ajuste que la describe. Para el análisis estadístico de los datos se utilizaron los paquetes Microsoft Excel y Statistica 6.0 StatSoft, Inc. 1984-2001.
Resultados y Discusión
En las dos localidades donde se realizaron los ensayos, Alajuela y Fraijanes, y en todas las épocas de siembra (Cuadro 1), siempre se dio sincronía en la floración de las variedades de frijol negra y blanca. Esta condición garantizó que la receptividad de los estigmas de las flores de la variedad de hipocotilo claro (PAN 68) y la viabilidad del polen de las flores de la variedad negra (UCR 55/Guaymí) ocurrieran al mismo tiempo, lo que pudo permitir eventos de polinización cruzada natural.
En Alajuela, la entomofauna que visitó las parcelas experimentales durante el período de floración fue abundante y diversa, dentro de los posibles polinizadores los de mayor cantidad y frecuencia fueron las abejas melíferas (Hymenoptera, Apis mellifera L.), que se indican como uno de los polinizadores más importantes en frijol por su adaptación a la estructura floral de esta planta (Free, 1966). También fue posible observar con frecuencia trigonas (Trigona spp) y en forma ocasional chiquizás (abejorros) (Bombus spp), estos últimos conocidos por su eficiencia como agentes polinizadores (Corbet et al., 1991; Ibarra et al., 1999; Ahrné et al., 2009). En Fraijanes, el principal polinizador fue el chiquizá, del que fue posible apreciar una mayor cantidad de individuos y frecuencia de visita a flores en el ensayo del 2002 que en el del 2003. En esta localidad también se observaron abejas melíferas en ambos períodos, pero en poca cantidad. Para ambas localidades la mayor actividad de posibles polinizadores se dio hacia horas del mediodía con el incremento de la temperatura y decreció cuando el día se nubló o iniciaron las precipitaciones.
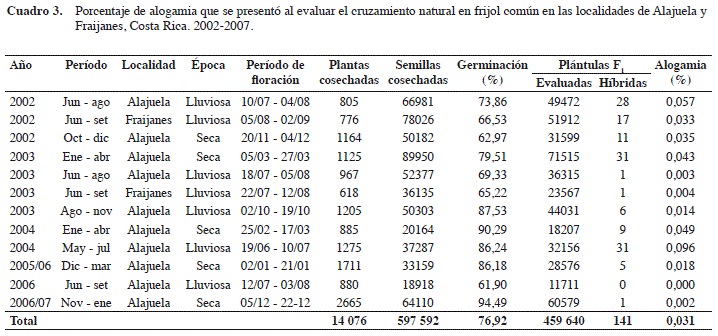
En Alajuela el cruzamiento natural varió desde 0,000 hasta 0,096%, mientras que en Fraijanes varió de 0,004 hasta 0,033% (Cuadro 3). En los doce ensayos realizados en ambas localidades se cosecharon 597 592 semillas de testa blanca, de las que germinó un 76,92%. Se obtuvieron 459 640 plántulas de las que 141 tuvieron hipocotilo morado e indicaron que efectivamente se produjo eventos de polinización cruzada en campo. Con base en el número de individuos híbridos en la F1 de la variedad PAN 68, se calculó el porcentaje de alogamia en las plántulas germinadas (Cuadro 3). Aunque el porcentaje de germinación de la semilla utilizada para evaluar el cruzamiento natural se podría considerar bajo, este se acerca al mínimo requerido (80%) para una semilla de categoría Certificada (ONS, 2012). En todo caso, la tasa de alogamia determinada en este trabajo fue muy baja (0,031%) por lo que el efecto del bajo porcentaje de germinación en su estimación sería mínimo.
En general, se estimó una alogamia de 0,031% y valores máximos de 0,096% en Alajuela y 0,033% en Fraijanes (Cuadro 3). Estos resultados son bajos en comparación con investigaciones similares realizadas en otros países para determinar el flujo de genes entre cultivares de frijol común, donde se indican valores de cruzamiento natural desde 0,14% en Brasil (Ferreira et al., 2007) hasta 7,45% en México (Miranda, 1971). A pesar de que los valores de polinización cruzada natural obtenidos en esta investigación son muy bajos, no distan mucho de los determinados anteriormente en Costa Rica, donde Alan y Moh (1966) obtuvieron una alogamia máxima de 0,20% en Alajuela y Pinchinat (1967) estimó 0,10% de cruzamiento natural, valor similar al máximo determinado en Alajuela para este trabajo.
La estimación de la tasa de alogamia puede verse afectada por la metodología empleada, pero también por factores relacionados con el genotipo y germoplasma utilizado, así como las condiciones agroecológicas y climáticas específicas de los sitios donde se realizó el trabajo (Miranda, 1971; Wells et al., 1988; Brunner y Beaver, 1989; Triana et al., 1993; Ramírez et al., 2000; Ferreira et al., 2007; Chaves et al., 2009). Por ello, en esta investigación se evaluó la influencia de las condiciones climáticas, la dirección predominante del viento y la distancia de la fuente de polen sobre el cruzamiento natural del frijol.
Con respecto a la metodología empleada, en esta investigación se utilizó un esquema de siembra en campo similar al empleado por Ferreira et al. (2007), quienes colocaron una parcela de una variedad de grano color negro (fuente de polen contaminante) rodeada por hileras de una variedad de grano blanco, y estimaron el porcentaje de alogamia por hilera y respecto a la distancia de la fuente de polen externo, excepto que ellos utilizaron el color de flor como mar-cador morfológico para distinguir las plantas híbridas en la F1. El sistema de siembra en campo empleado en esta investigación permitió simular de mejor manera lo que sucedería en una condición de siembra comercial, donde se presentarían parcelas de distintas variedades de forma contigua. En este trabajo se evaluó un mayor número de ensayos y semillas que en otras investigaciones sobre alogamia en frijol común (Alan y Moh, 1966; Miranda, 1971; Ortega, 1974; Wells et al., 1988; Brunner y Beaver, 1989; Triana et al., 1993; Ramírez et al., 2000; Royer et al., 2002; Ferreira et al., 2007). El cálculo del porcentaje de cruzamiento natural fue idéntico al utilizado por Alan y Moh (1966), Ortega (1974), Ramírez et al. (2000) y Royer et al. (2002), quienes se basaron en la cantidad de plántulas con hipocotilo morado respecto al número total de plántulas germinadas.
En estudios de polinización cruzada natural en frijol se ha utilizado como marcadores morfológicos para determinar la tasa de alogamia el color de hipocotilo (Alan y Moh, 1966; Ortega, 1974; Triana et al., 1993; Ibarra et al., 1997; Ramírez et al., 2000; Royer et al., 2002; Chaves et al., 2009), el color de la flor (Miranda, 1971; Ibarra et al., 1997; Royer et al., 2002; Ferreira et al., 2007), el color de la semilla (Stoetzer, 1984; Wells et al., 1988; Brunner y Beaver, 1989), el color y forma de la hoja (Brunner y Beaver, 1989), y el hábito de crecimiento de la planta (Crispín, 1960; Tucker y Harding, 1975). En esta investigación se utilizó el color del hipocotilo para estimar el porcentaje de polinización cruzada debido a que es parte integral del embrión producto de la polinización (autopolinización o polinización cruzada), mientras que el tegumento de la semilla corresponde a un tejido maternal (Weinstein, 1926). Además, el color de hipocotilo es una característica dominante, estable, independiente del ambiente y que puede distinguirse fácilmente en los primeros estadíos de desarrollo de la planta (Alan y Moh, 1966). Al respecto, Basset (1996) e Ibarra et al. (1996) indican que este color está determinado por un solo locus genético (P/p), así el homocigoto recesivo (pp) es verde (sin color) mientras que el dominante (PP) y el heterocigoto (Pp) son morados. Por su parte, Ramírez et al. (2000) indican que el uso de este marcador permite manejar un número mayor de individuos e incrementa la precisión de la estimación.
En cuanto a la condición climática, no hubo diferencias significativas en el porcentaje de cruzamiento natural entre los ensayos que fueron sembrados durante la estación lluviosa (0,038%), con respecto a aquellos sembrados durante la estación seca (0,027%) en Alajuela. Los resultados obtenidos fueron más bajos pero coinciden con lo indicado por Vieira (1960), quien no encontró diferencias significativas en el porcentaje de alogamia estimado para la época seca (0,18%) con respecto a la época lluviosa (0,70%) en Viçoza, Minas Gerais, Brasil. Al respecto, Royer et al. (2002), en Maringá, Paraná, Brasil, determinaron promedios de hibridación natural de 1,23; 1,34; 1,25 y 0,71% para los cultivares Jalo Precoce, Pérola, Carioca y Michelite respectivamente en la época de lluvias, y de 0,82; 0,75; 0,79 y 0,76% para Jalo Precoce, Pérola, Carioca y Michelite, respectivamente en el período seco. Sin embargo, no encontraron diferencias significativas en la tasa de alogamia para las épocas seca y lluviosa en ninguno de los cultivares sembrados a cuatro distancias distintas, excepto para el cultivar Pérola a un espaciamiento de 0,2 m. Por su parte, Crispín (1960) en México, indica que el porcentaje de polinización cruzada natural fue mayor en las siembras donde no se necesitó riego que en las que se hicieron con riego, y Ortega (1974) en Venezuela, determinó una alogamia de 0,22% en época seca y de 0,49% en época de lluvias, pero estos autores no indican si las diferencias encontradas fueron significativas o no.
Otro de los factores estudiados en este trabajo fue la influencia de la dirección predominante del viento sobre la polinización cruzada natural en frijol común. En ninguna de las dos localidades donde se realizó el estudio se encontraron diferencias significativas en la tasa de alogamia de aquellas hileras que se situaron en el bloque B, a favor de la dirección predominante del viento, con respecto a las ubicadas en el bloque A, en contra de la dirección predominante del viento (Figura 1). En Fraijanes se tuvo una alogamia de 0,021% en el bloque B y de 0,027% en el bloque A, mientras que en Alajuela 0,030% en el bloque B y de 0,034% en el bloque A. En Alajuela tampoco se encontraron diferencias significativas en el porcentaje de cruzamiento natural entre un bloque y otro durante la época lluviosa o la seca, a pesar de que en época seca (diciembre a marzo) la dirección del viento es más definida y las ráfagas son de mayor velocidad (Gutiérrez et al., 1997). Los resultados obtenidos indican que el viento no ejerce influencia alguna sobre la polinización cruzada en frijol y lo único que podría favorecer es el arrastre de los polinizadores.
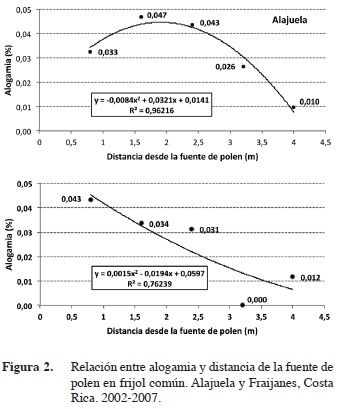
El factor que fue determinante en la tasa de alogamia en frijol común es la distancia respecto a la fuente de polen. La tasa de polinización cruzada natural decreció según aumentó la distancia respecto a la fuente de polen y se obtuvo una correlación inversa de tipo polinomial en las dos localidades donde se sembraron los ensayos (Figura 2). Solo en Alajuela la correlación fue significativa (p<0,05; r2=0,9622), mientras que en Fraijanes, solo se mantuvo la tendencia con una fuerte asociación entre las variables (r2=0,7624), talvez por falta de un mayor número de ensayos que le dieran más consistencia a los datos. El resultado obtenido en Alajuela, concuerda con Miranda (1971), Royer et al. (2000) y Ferreira et al. (2007), quienes indican que el cruzamiento natural en frijol decrece conforme aumenta la distancia de la fuente de polen.
El porcentaje de polinización cruzada disminuyó casi hasta cero a los 4 m de la fuente de polen en Alajuela (Figura 2), resultado que concuerda con lo encontrado por Ferreira et al. (2007), quienes determinaron que la alogamia en frijol es prácticamente cero (0,0045%) a una distancia de 3,25 m de la fuente de polen. El resultado obtenido en la EEAFBM (Alajuela), donde se produce semilla categoría Fundación de frijol, coincide con la norma de aislamiento mínimo exigida por la Oficina Nacional de Semillas (ONS), que estipula 3 m para campos de producción de semilla de frijol categoría Certificada, 5 m para la categoría Registrada y 10 m para la categoría Fundación (ONS, 2012). De acuerdo con los datos obtenidos en este estudio para la variedad Guaymí, una de las variedades negras de uso comercial en Costa Rica, si se quisiera evitar por completo la contaminación genética de los campos de producción de semilla Certificada a través del fenómeno de polinización cruzada, se debería respetar al menos 5 m de aislamiento, como se estipula para las categorías Registrada y Fundación. Sin embargo, con 3 m de aislamiento se garantizaría cumplir con el 98% de pureza genética mínima exigida por la norma de la ONS para todas las categorías de semilla de frijol (ONS, 2012).
Agradecimientos
Al German Federal Ministry for Economic Cooperation and Development (BMZ), al Centro Internacional de Agricultura Tropical (CIAT) y a la Universidad de Costa Rica (Proyecto 736-A1-553 inscrito en Vicerrectoría de Investigación), instituciones que financiaron el desarrollo de esta investigación. Al Instituto Meteorológico Nacional de Costa Rica por facilitar los datos de las condiciones climáticas imperantes durante la realización de los ensayos. Por su valiosa colaboración durante la realización de este trabajo a los señores Catalino Sevilla y Greivin Soto; a la señorita Laura Barboza; a las ingenieras Marcela Acuña y Sofía Rodríguez, al MSc. Félix Zumbado; a los señores Isauro González, Francisco González, Luis Cascante (qdDg), Marvin Arias y José Oreamuno del Programa de Leguminosas de la Estación Experimental Fabio Baudrit de la Universidad de Costa Rica; al Ing. Juan Carlos Hernández y a los señores Ronald Campos, Javier Campos, Jonathan Campos y Jesús Arroyo del Instituto Nacional de Innovación y Transferencia en Tecnología Agropecuaria (INTA) de Costa Rica.
Notas y Citas
1. Trabajo conjunto entre el Centro Internacional de Agricultura Tropical (CIAT) y la Universidad de Costa Rica (Proyecto V.I. 736-A1-553).
Literatura Citada
Ahrné, K., J. Bengtsson, y T. Elmqvist. 2009. Bumble bees (Bombus spp) along a gradient of increasing urbanization. PLoS ONE 4(5):e5574. doi:10.1371/ journal.pone.0005574. [ Links ]
Alan, J., y C. Moh. 1966. Determinación del porcentaje de cruzamiento natural en el frijol común (Phaseolus vulgaris L.) en Alajuela, Costa Rica. Turrialba 16(2):156-158. [ Links ]
Aragão, F., G. Vianna, M. Albino, y E. Rech. 2002. Transgenic dry bean tolerant to the herbicide glufosinate ammonium. Crop Sci. 42:1298-1302. [ Links ]
Aragão, F., y J. Faria. 2009. First transgenic geminivirus resistant plant in the field. Nat. Biotechnol. 27:1086-1088. [ Links ]
Araya, R., y J. Acosta. 2000. Adaptación del frijol a suelos de baja fertilidad bajo presión de enfermedades foliares en Costa Rica. Agron. Mesoam. 11(2):31-36. [ Links ]
Bassett, M. 1996. List of genes – Phaseolus vulgaris L. Annual Report of the Bean Improvement Coopertive (USA) 39:1-19. [ Links ]
Bliss, F. 1980. Common bean. En: W. Ferh, y H. Hadley, editores, Hybridization of crops plants. American Society of Agronomy and Crop Science, Wisconsin, USA. p. 273-284. [ Links ]
Bonfim, K., J. Faria, E. Nogueira, É. Mendes, y F. Aragão. 2007. RNAi-mediated resistance to Bean golden mosaic virus in genetically engineered common bean (Phaseolus vulgaris). Mol. Plant-Microbe Interact. 20:717-726. [ Links ]
Broughton, W., G. Hernández, M. Blair, S. Beebe, P. Gepts, y J. Vanderleyden. 2003. Beans (Phaseolus spp.) - model food legumes. Plant Soil 252(1):55-128. [ Links ]
Brunner, B., y J. Beaver. 1989. Estimation of outcrossing of the common bean in Puerto Rico. HortScience 24:669-771. [ Links ]
Chaves, N., R. Araya, y D. Debouck. 2009. Flujo de genes entre frijol común y silvestre en Costa Rica. Agron. Mesoam. 20:237-244. [ Links ]
Corbet, S., I. Williams, y J. Osborne. 1991. Bees and the pollination of crops and wild flowers in the European community. Bee World 72:47-59. [ Links ]
Crispín, A. 1960. Cruzamiento natural en frijol. Agricultura Técnica en México 11:38-39. [ Links ]
Ferreira, J., M. Álvarez, M. Fueyo, A. Roca, y R. Giraldez. 2000. Determination of the outcrossing rates of Phaseolus vulgaris L. using seed protein markers. Euphytica 113:259-263. [ Links ]
Ferreira, J., J. De Souza, A. Teixeira, F. De Lanes, P. Cecon, y A. Borém. 2007. Gene flow in common bean (Phaseolus vulgaris L.). Euphytica 153:165-170. [ Links ]
Free, J. 1966. The pollination of beans Phaseolus multiflorus and Phaseolus vulgaris by honeybees. J. Apicult.Res. 5(2):87-91. [ Links ]
Gepts, P., A. González, R. Papa, J. Acosta, A. Wong, A. Delgado. 2000. Outcrossing in Mexican wild and domesticated populations of common bean. Annual Report of the Bean Improvement Cooperative 43:25-26. [ Links ]
Gepts, P., y R. Papa. 2003. Possible effects of (trans)gene flow from crops on the genetic diversity from landraces and wild relatives. Environ. Biosafety Res. 2(2):89-103. [ Links ]
Gutiérrez, M., D. Soto, y M. Alpízar. 1997. Cuarenta años de observaciones meteorológicas en la Estación Experimental Fabio Baudrit Moreno. Boletín Técnico Estación Experimental Fabio Baudrit Moreno (BOLTEC) 30(2):1-14. [ Links ]
Gutiérrez, M.V., D. Soto, y M. Alpízar. 1999-2002. El clima en la Sub-Estación Experimental Fraijanes, en un ecosistema montano bajo y húmedo. Revista de Agricultura Tropical (Costa Rica) 32:39-45. [ Links ]
Hails, R. 2000. Genetically modified plants: the debate continues. Trends Ecol. Evol. 15:14-18. [ Links ]
Hails, R. 2002. Assessing the risks associated with new agricultural practices. Nature 418:685-688. [ Links ]
Harlan, J., y J. De Wet. 1971. Toward a rational classification of cultivated plants. Taxon 20:509-517. [ Links ]
Holdridge, L.R. 1982. Ecología basada en zonas de vida. Instituto Interamericano de Cooperación para la Agricultura (IICA), San José, Costa Rica. [ Links ]
Ibarra, F., N. Ellstrand, y J. Giles. 1996. Multiple paternity in common bean (Phaseolus vulgaris L., Fabaceae). Am. J. Bot. 83(6):749-758. [ Links ]
Ibarra, F., B. Ehdaie, y J. Waines. 1997. Estimation of outcrossing rate in common bean. Crop Sci. 37:60-65. [ Links ]
Ibarra, F., D. Barnhart, B. Ehdaie, K. Knio, y J. Waines. 1999. Effects of insect tripping on seed yield of common bean. Crop Sci. 39:428-433. [ Links ]
Kwapata, K., T. Nguyen, y M. Sticklen. 2012. Genetic transformation of common bean (Phaseolus vulgaris L.) with the Gus color marker, the Bar herbicide resistance, and the barley (Hordeum vulgare) HVA1 drought tolerance genes. Int. J. Agron. Volume 2012, Article ID 198960.doi:10.1155/2012/198960. [ Links ]
León, J. 1987. Botánica de los cultivos tropicales. 2 ed. Instituto Interamericano de Cooperación para la Agricultura (IICA), San José, Costa Rica. [ Links ]
Mackie, W., y F. Smith. 1935. Evidence of field hybridization in beans. J. Am. Soc. Agron. 27:903-909. [ Links ]
Miranda, S. 1971. Cruzamiento natural en frijol. Agricultura Técnica en México 3(2):48-52. [ Links ]
ONS (Oficina Nacional de Semillas de Costa Rica). 2012. Reglamento técnico para la producción de semilla certificada de frijol. http://www.ofinase.go.cr (Consultado 7 abr. 2013). [ Links ]
Ortega, S. 1974. Polinización cruzada natural de la caraota (P. vulgaris L.) en Venezuela. Agronomía Tropical 24(1):27-32. [ Links ]
Papa, R., y P. Gepts. 2003. Asymmetry of gene flow and differential geographical structure of molecular diversity in wild and domesticated common bean (Phaseolus vulgaris L.) from Mesoamerica. Theor. Appl. Genet. 106:239-250. [ Links ]
Pinchinat, A. 1967. Cruzamiento natural en el frijol común (Phaseolus vulgaris L.) en Costa Rica. En: Editorial Nuestra América, editor, VII Reunión Latinoamericana de Fitotecnia. Maracay, Venezuela. p. 208-209. [ Links ]
Ramírez, P., A. Reyes, J. Estrada, F. Castillo, S. Cruz, y L. Barrios. 2000. Cruzamiento natural de frijol común en los valles altos de México. Revista Fitotecnia Mexicana 23(2):277-292. [ Links ]
Royer, M., M. Gonçalves, C. Scapim, P. Soares, V. Filho, y Y. Terada. 2002. Outcrossing in common bean. Crop Breed. Appl. Biotechnol. 2(1):49-54. [ Links ]
Stoetzer, H. 1984. Natural cross-polinization in bean in Ethiopia. Annual Report of the Bean Improvement Cooperative 27:99-100. [ Links ]
Triana, B., M. Iwagana, H. Rubiano, y M. Andrade. 1993. A study of allogamy in wild Phaseolus vulgaris. Annual Report of the Bean Improvement Cooperative 36:20-21. [ Links ]
Tucker, C., y J. Harding. 1975. Outcrossing in common bean Phaseolus vulgaris L. J. Am. Soc. Hortic. Sci. 100:283-285. [ Links ]
Vieira, C. 1960. Sobre a hibridação natural em Phaseolus vulgaris L. Ceres 11:103-107. [ Links ]
Webster, B., C. Tucker, y S. Lynch. 1977. A morphological study of the development of reproductive structures of Phaseolus vulgaris L. J. Am. Soc. Hortic. Sci. 102:640-643. [ Links ]
Webster, B., R. Ross, y T. Evans. 1982. Nectar and the nectary of Phaseolus vulgaris L. J. Am. Soc. Hortic. Sci. 107:497-503. [ Links ]
Weinstein, A. 1926. Cytological studies on Phaseolus vulgaris. Am. J. Bot. 13(4):248-263. [ Links ]
Wells, W., W. Isom, y J. Waines. 1988. Outcrossing rates of six common bean lines. Crop Sci. 28:177-178. [ Links ]
*Correspondencia a:
Programa de Leguminosas, Estación Experimental Fabio Baudrit Moreno, Universidad de Costa Rica. Apdo. postal 183-4050 Alajuela, Costa Rica. Correo electrónico nfchaves@gmail.com
2 Programa de Leguminosas, Estación Experimental Fabio Baudrit Moreno, Universidad de Costa Rica. Apdo. postal 183-4050 Alajuela, Costa Rica. Correo electrónico avillalo2005@hotmail.com
3 Programa de Recursos Genéticos, Centro Internacional de Agricultura Tropical (CIAT). A.A.6713, km 17, recta Cali-Palmira, Cali, Colombia. Correo electrónico d.debouck@cgiar.org
Recibido: 8 de julio, 2013. Aceptado: 18 de marzo, 2014


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}