










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgronomía Mesoamericana
On-line version ISSN 2215-3608Print version ISSN 1659-1321
Agron. Mesoam vol.24 n.2 San Pedro Jul./Dec. 2013
Comunicación corta
Efecto de poblaciones de Meloidogyne sp. en el desarrollo y rendimiento del tomate
Effect of Meloidogyne sp. on growth and yield of tomato
Wilber Salazar-Antón2*, Tomás de Jesús Guzmán-Hernández3*
Effect of Meloidogyne sp. on growth and yield of tomato
Wilber Salazar-Antón2*, Tomás de Jesús Guzmán-Hernández3*
*Dirección para correspondencia
Resumen
El objetivo de este trabajo fue determinar el efecto de la población inicial de Meloidogyne sp. en el cultivo del tomate. El estudio se realizó en León, Nicaragua en el año 2011. Las variables medidas fueron número de nematodos al trasplante y al momento de la cosecha, altura de la planta al momento de la madurez fisiológica y el rendimiento en kilogramos. La extracción de nematodos se realizó mediante el método de Baermann modificado. En el estudio, la densidad inicial (Pi) presentó una correlación lineal positiva con la densidad final (Pf). Sin embargo, se observó que cuando la Pi era igual a 400 nematodos/100 g de suelo, el factor de reproducción se incrementaba a 3,64; por el contrario, cuando la Pi era igual a 700 nematodos/100 g de suelo, el factor de reproducción se reducía a 2,48. La Pi se correlacionó inversamente con el peso de frutos y altura de las plantas. Plantas con 200 nematodos/100 g de suelo presentaban peso de frutos de 2,19 kg y altura de 153,20 cm en comparación con 0,93 kg y 135,24 cm en plantas con 600 nematodos/100 g de suelo. Existió una correlación positiva entre Pi y Pf de Meloidogyne sp. en el tomate. Las poblaciones de este nematodo, se correlacionaron inversamente con las variables altura y peso de los frutos.
Palabras clave: nematodos fitopárasitos, daño radicular, nematodo agallador, reducción del peso de frutos.
Abstract
The objective of this work was to determinate the relationship between initial density and final density of Meloidogyne sp in tomato crop. The study was carried out at León, Nicaragua during 2011. The variables measured were number of nematodes at transplant, number of nematodes at harvest time, height at maturity of plants and yield in kilograms. Nematode extraction was performed through the Baermann modified method. In the study, the initial density (Pi) showed a positive linear correlation with the final density (Pf). However, it was observed that when Pi was equal to 400 nematodes/100 g of soil, the reproduction factor was increased to 3.64, on the contrary, when Pi was equal to 700 nematodes/100 g of soil, reproduction factor was reduced to 2.48. Pi was inversely correlated to weight of tomato fruits and height of plants. Plants with 200 nematodes/100 g of soil had fruit weight 2.19 kg and height 153.20 cm compared to 0.93 kg and 135.24 cm in plants with 600 nematodes/100 g of soil. There was a positive correlation between Pi and Pf of Meloidogyne sp. in tomatoes. The populations of this nematode, were inversely correlated with plant height and fruits weight.
Keywords: phytoparasitic nematodes, root damage, root knot nematode, fruit weight reduction.
Iintroducción
El nematodo Meloidogyne sp. es uno de los patógenos más nocivos del tomate a nivel mundial, debido a que afecta severamente las raíces de este cultivo (Sikora y Fernández 2005, Bhattarai et al. 2008). Se caracteriza por tener un hábito alimenticio polífago con un amplio rango de hospederos especialmente en países tropicales y subtropicales (Sikora y Fernández 2005, Moens et al. 2009). Esto ha hecho que sea considerado el nematodo fitoparásito de mayor importancia económica en el mundo.
Los síntomas característicos de este nematodo provocan en la planta diferentes grados de achaparramiento, falta de vigor, deficiencias nutricionales y marchitamiento bajo condiciones de estrés (Shurtleff y Averre III 2000, Zaqui et al. 2001). Estas afectaciones generan pérdidas a nivel mundial que se estima superan los $US 100 billones (Bird y Kaloshian 2003), siendo más de la mitad de estas pérdidas atribuidas a Meloidogyne sp (Bent et al. 2008).
En el cultivo del tomate, Meloidogyne spp. es muy importante por su rápida expansión, alta frecuencia de infestación y su capacidad para reducir su rendimiento hasta en un 68% (Esparrago y Navas 1995, Chindo y Khan 1988, Adesiyan et al. 1990). Los daños causados no solo reducen el número de frutos del cultivo, sino que también afecta la calidad de los mismos impactando de esta forma en sus precios (Sasser y Carter 1982, Moens et al. 2009).
Se ha documentado que la población inicial de Meloidogyne sp. influye en la severidad del daño, afectando el desarrollo del tomate (Mekete et al. 2003, Salazar-Antón y Guzmán-Hernández 2013) y su rendimiento (Ehwaeti et al. 1998). A pesar de la innegable importancia de Meloidogyne sp. en el cultivo del tomate, no existen estudios en Nicaragua que indiquen el tipo de relación existente entre su densidad al momento del trasplante y los rendimientos del tomate. Asimismo, se carece de información técnica que permita predecir la población final de este nematodo basado en la población inicial del mismo.
El presente estudio tuvo como objetivo determinar el efecto de la población inicial de Meloidogyne sp. en el cultivo del tomate.
Materiales y Métodos
Ubicación del estudio y condiciones agroclimáticas
El presente estudio se llevó a cabo en el campus agropecuario de la Universidad nacional autónoma de Nicaragua-León (UNAN-León), ubicado en León, Nicaragua, durante el año 2011. Las condiciones agroclimáticas fueron 1910 mm anuales, con una humedad relativa promedio de 70%, con un suelo predominantemente franco arenoso, pH de 6,7 y una temperatura promedio de 31oC (INETER 2011).
Metodología
El estudio se realizó en una parcela de 4000 m2 previamente diagnosticada con altas poblaciones de Meloidogyne sp. La variedad de tomate utilizada fue Shanty la cual se caracteriza por producir plantas vigorosas, de frutos rojo intenso en forma de óvalo con susceptibilidad a Meloidogyne spp.
La selección de plantas sujetas a muestreo nematológico se realizó mediante uno aleatorio sistemático, en el cual se eligió una planta al azar y a partir de ella, a intervalos constantes, se eligieron las demás plantas hasta completar la muestra.
La población estuvo formada por 1500 plantas con una muestra de 50 plantas. El intervalo de muestreo se obtuvo dividiendo la población en estudio entre la muestra (1500/50=30). Se eligió el elemento de inicio, tomando aleatoriamente un número entre el 1 y 30, el número seleccionado fue 25 y a partir de él se obtuvieron los restantes elementos de la muestra. Iniciando en la planta número 25 se muestrearon las plantas 25, 50, 75, 100, hasta la planta 1250.
El muestreo de nematodos se realizó en las 50 plantas previamente identificadas. La densidad inicial de Meloidogyne sp. se contabilizó al momento del trasplante del tomate y la densidad final al momento de la cosecha. Cada muestra fue de 200 g de suelo, extraídas de la rizosfera de la planta con un barreno metálico de 2,2 cm de diámetro interno, el que se introducía 30 centímetros en el suelo. Las muestras fueron empacadas en bolsas plásticas, rotuladas y enviadas al Laboratorio de Fitopatología de la UNAN-León para su análisis.
Extracción de nematodos de suelo
La extracción de nematodos se realizó mediante el método de Baermann modificado (Hooper et al.2005). Se pesaron 100 g de suelo y se colocaron en un filtro de papel toalla extendido sobre un tamiz dentro de un recipiente plástico. Luego se agregó agua hasta humedecer el nivel superior del suelo sin saturarlo. Se incubó por 48 horas y luego se filtró la suspensión dos veces a través de un tamiz de 400 mesh, colectando 20 ml de suspensión conteniendo los nematodos.
Para la identificación se colocó una gota de la suspensión conteniendo los nematodos en un porta objeto cubierto, se flameó con un mechero de alcohol por cinco segundos para inmovilizarlos. Se realizó la identificación utilizando parámetros morfométricos y la clave de identificación de nematodos diseñada por mai y mullin (1996) utilizando un microscopio. Una vez obtenida la solución conteniendo los juveniles, se homogenizó para tomar diez alicuotas de 5 ml cada uno y cuantificar la población utilizando un estereoscopio (Castro et al. 2011).
Medición de variables
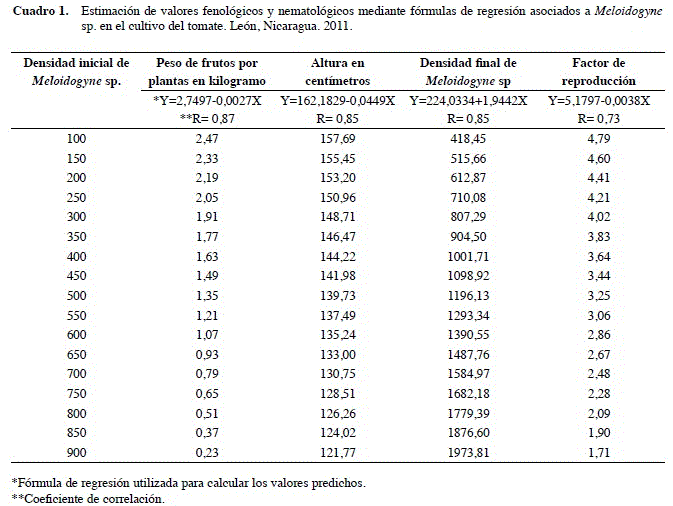
Durante la fase de campo se midió la altura de las plantas de tomate al momento de la cosecha, al mismo tiempo se cuantificó y pesó los frutos de cada planta. Finalmente, se calculó el Factor de reproducción de Meloidogyne sp., mediante la fórmula FR= Pf/Pi. Donde Pi es la densidad de nematodos al momento del trasplante y Pf es la densidad de nematodos al momento de la cosecha.
Manejo agronómico
El cultivo del tomate fue manejado convencionalmente utilizando el sistema productivo que los agricultores de la zona occidental de Nicaragua realizan. El tipo de suelo fue franco arenoso, bien drenado y profundo, con una topografía plana del 2% de pendiente. Se preparó el suelo mecánicamente, con un pase de arado y dos pases de grada. Se aplicó productos bactericidas como sulfato de cobre al pie de cada plántula a razón de 1,6 l t/ha y fungicidas Benomil del grupo de los Benzimidazol a razón de 1 kg por 50 galones de agua. Sin embargo, no se aplicó ningún producto nematicida al suelo con el fin de no interferir en la dinámica poblacional y tasa reproductiva de los nematodos.
Análisis económico y estadístico
Se cuantificó el número de frutos de tomate por planta y se calculó el peso en kilogramos de frutos que debe ser generado por planta para la obtención de beneficios económicos. Para el análisis se tomó en cuenta el número de plantas por hectárea, los costos de producción y el precio del kilogramo de frutos. Los datos obtenidos fueron analizados mediante una regresión lineal simple. Contrastándose la densidad inicial de nematodos con los datos de rendimiento, altura de la planta y densidad de Meloidogyne sp al momento de la cosecha. Para el análisis se utilizó el programa Statistical Package for Social Science (SPSS) (Esparza s.f.).
Resultados y Discusión
Relación entre la población inicial y final de Meloidogyne sp. en tomate
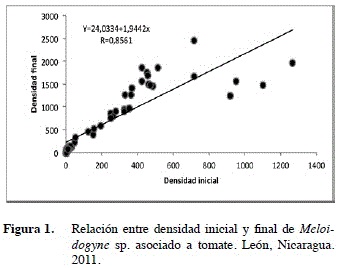
La regresión lineal realizada a la densidad inicial y final de Meloidogyne sp. en el cultivo del tomate, indicó que ambas variables están positivamente co- rrelacionadas y tienen una relación lineal significativa (Figura 1), este tipo de relación ha sido previamente reportada para Meloidogyne sp. en tomate, por Ferris (1985).
De la fórmula de la recta generada en el análisis de regresión (Y=224,03+1,944X), se deduce que por cada nematodo, encontrado al momento del trasplante del tomate, correspondió a un incremento de 1,94 nematodos al final del ciclo del cultivo. El coeficiente de correlación r=0,85 (Figura 1), muestra una estrecha relación entre ambas variables y el coeficiente de determinación de estas variables r2= 0,73 indica que el 73% de la variabilidad en la densidad final de Meloidogyne sp. es explicada por la densidad inicial.
Meloidogyne sp. experimentó un crecimiento lineal durante las diferentes etapas de crecimiento del tomate. no obstante, al correlacionarse su densidad inicial con el Factor de reproducción (FR) se obtuvo una correlación inversa (Figura 2).
Por otro lado, una alta Pi puede causar un deterioro de los sitios de infección y generar una acumulación de desechos metabólicos de estos organismos que afectarían su desarrollo (Ferris 1985), por lo que se puede afirmar que el FR de una población de Meloidogyne sp decrece cuando las poblaciones son excesivas, debido a la limitación de alimentos y espacio.
Relación entre población inicial de nematodos y su efecto sobre el rendimiento del tomate
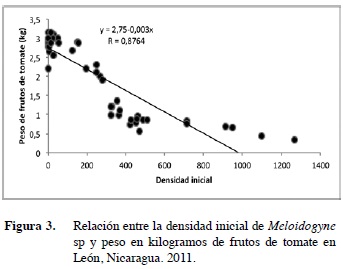
El análisis económico indicó que en promedio cada planta debió producir al menos 1,02 kg de tomate. A los datos de tomate producido se les realizó una regresión lineal para determinar el nivel de influencia de la Pi sobre ellos. La fórmula de la regresión lineal obtenida fue Y=2,755-0,003x (Figura 3), indicando que por cada nematodo encontrado al inicio del cultivo hubo una reducción en la producción de 0,003 kg por planta.
El estudio demostró que la Pi fue negativamente correlacionada con los rendimientos del tomate. Una Pi mayor a los 620 nematodos/100 g de suelo redujeron el rendimiento a menos de 1,02 kg de frutos por planta. Esta situación generó ingresos económicos menores que los costos de producción, lo que ocasiona pérdidas económicas. Este nivel de daño supera los obtenidos por otros autores que indican que con una Pi de 200 juveniles/100 g de suelo se causa daños económicos que hacen necesarias medidas de control para reducir sus poblaciones y evitar pérdidas (Chindo y Khan 1988). Igualmente, este resultado contrasta con el reportado por Di Vito y Ekanayake (1983) y Chan y López (1992) quienes obtuvieron niveles de daño para tomate con 330 nematodos/100 g de suelo. Estos datos permiten ilustrar la variabilidad que existe al realizar mediciones de estos parámetros debido a la gran variedad de factores ambientales y varietales que influyen en sus poblaciones y en sus niveles de daño (Ekanayake y Di Vito 1984).
Una población de 620 nematodos/100 g de suelo, causa daños a las raíces incluyendo severos agallamientos radiculares que impiden su funcionamiento y dificultan el normal desarrollo de la planta (Olthof y Potter 1977, Mekete et al. 2003). Por otro lado, plántulas de tomate expuestas a una alta Pi sufren mayor daño de Meloidogyne sp., del mismo modo, este patógeno ataca con mayor severidad a plántulas que a plantas adultas (Philis 1990). Esto coincide con nuestro estudio, donde una alta Pi al momento del trasplante contribuyó a una reducción significativa en los rendimientos.
Por otro lado, utilizando la fórmula de la recta generada, se pudo estimar que con una Pi igual a 200 nematodos/100 g de suelo, se obtienen rendimientos de 2,19 kg por plantas, suficientes para generar beneficios económicos. Estudios previos indican que poblaciones menores de 200 individuos por 100 g de suelo estimulan el desarrollo de la planta e incrementan los rendimientos del cultivo (Sayre y Toyama 1964). Estos resultados coinciden igualmente con lo reportado más recientemente por Fawole y Mai (1979) y por Belair y Tremblay (1995), quienes afirman que en presencia de un suelo con humedad y fertilidad adecuada para la planta afectada por Meloidogyne sp., esta genera numerosas raíces secundarias que favorecen el crecimiento y rendimiento del tomate.
Relación entre población inicial de nematodos y su efecto sobre la altura de las plantas de tomate
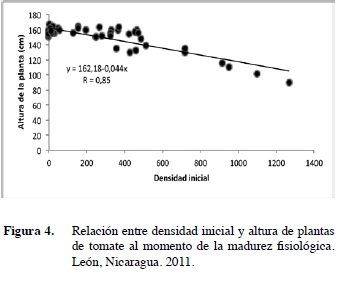
La regresión realizada a la Pi y altura de la planta indica que ambas variables están inversamente correlacionadas entre sí. La fórmula de la regresión obtenida fue Y=161,91-0,044X, lo que indica que por cada nematodo encontrado al momento del trasplante habrá una reducción en la altura de la planta de 0,044 cm (Figura 4).
Este tipo de relación ha sido previamente descrita por diversos autores (Khan et al. 2000, Ploeg 2000, Kumar y Khanna 2006) quienes afirman que un incremento en la Pi causa una reducción significativa en la altura de las plantas cuando estas alcanzan su madurez. Otros autores señalan que una alta Pi de Meloidogyne sp. genera un efecto adverso sobre el peso de las plantas (Ehwaeti et al. 1998, Mekete et al. 2003). Esto demuestra que el achaparramiento inducido por nema- todos afecta también el número de hojas y el peso de retoños y raíces (Khan et al. 2000)
Se observó que cuando Pi era igual a 200 nematodos/100 g de suelo la altura de las plantas fue de 153,20 cm (Cuadro 1). Sin embargo, cuando la Pi fue igual a 600 individuos/100 g de suelo la altura de las plantas decreció a 135,24 cm, lo que representó una reducción de un 12,37% (Cuadro 1). Este resultado coincide con lo reportado por Ahmad et al. (1988) y Zaki et al. (2001), quienes mencionaron que Meloidogyne sp. reduce significativamente la altura de plantas de tomate cuando son expuestas a altas densidades de este patógeno en su etapa de plántulas.
Al comparar la altura de plantas expuestas a Pi entre 4-250 nematodos/100 g de suelo con plantas sin afectación de nematodos, se observó que las afectadas superaron en un 4% a la altura de las plantas sanas. Este resultado coincide con estudios previos que han demostrado que bajas poblaciones de nematodos (26-184/g de suelo) estimulan el crecimiento vegetativo del tomate en comparación con altas poblaciones (612-2795/ 100 g de suelo) que reducen la altura de las plantas (Olthof y Potter 1977). Este comportamiento de las plantas se debe a que un leve daño de nematodos incrementa el tamaño del sistema radicular, debido a la formación de raíces adventicias alrededor de las áreas agalladas. Esto último, facilita la nutrición de la planta e incrementa la capacidad de la misma de tolerar el daño de los nematodos. Por el contrario, altas densidades de Meloidogyne sp. impiden la tasa de expansión de las raíces afectando el crecimiento de la planta (Olthof y Potter 1977, Chan y López 1992, Belair y Tremblay 1995). Esta aseveración es confirmada por Ravichandra (2008) quien ha indicado que hay evidencias que indican que bajas densidades de nematodos pueden reparar el daño en las raíces causados por nematodos fitoparásitos y aun estimular su crecimiento.
El incremento en la altura de las plantas en presencia de bajas poblaciones de nematodos, puede explicarse según el modelo de Seinhorst (1968), que indica que los nematodos fitoparásitos tienen dos efectos sobre las plantas, uno de estimulación y otro de inhibición. Adicionalmente, Wallace (1971) propuso que cuando el proceso estimulatorio es mayor que el inhibitorio la planta afectada por nematodos muestra un mayor crecimiento en comparación con plantas sin nematodos. Este tipo de comportamiento está acorde con lo reportado en otros cultivos como banano (Chávez-Velazco et al. 2009) y café (Morera y López 1987), los que presentan mejor desempeño en presencia de nematodos que en su ausencia.
Literatura citada
Adesiyan, SO; Caveness, FE; Adeniji, Mo; Fawole B. 1990. Nematode pests of tropical crops. Ibadan, Nigeria, Heinemann Educational Books. p. 19-26. [ Links ]
Ahmad, A; Tlyagi, SA; Alam, MM. 1988. self interaction of the root-knot nematode, Meloidogyne incognita on tomato. Nematologia Mediterranea 16:227-228. [ Links ]
Belair, G; Tremblay, N. 1995. The influence of chitinurea amendments applied to an organic soil on a Meloidogyne hapla population and on the growth of greenhouse tomato. Phytoprotection 76:75-80. [ Links ]
Bent, E; Loffredo, A; McKenry, MV; Becker, JO; Borneman, J. 2008. Detection and investigation of soil biological activity against Meloidogyne incognita. J. Nematol.40:109-118. [ Links ]
Bhattarai, KK; Xie, QG; Mantelin, S; Bishnoi, U; Girke, T; navarre, Da; Kaloshian, I. 2008. Tomato susceptibility to root-knot nematodes requires an intact jasmonic acid signaling pathway. Mol. Plant- Microbe Interact.21:1205-1214. [ Links ]
Bird, DM; Kaloshian, I. 2003. Are roots special? Nematodes have their say. Physiological and Molecular Plant Pathology 62:115-123. [ Links ]
Castro, L; Flores, L; Uribe, L. 2011. efecto del vermicompost y quitina sobre el control de Meloidogyne incognita en tomate a nivel de invernadero. Agronomía Costarricense 35(2):21-32. [ Links ]
Chan, S; López, R. 1986. Efecto de diferentes densidades iniciales de Meloidogyne incognita sobre el crecimiento del tomate. Agronomía Costarricense 16(2):165-169. [ Links ]
Chávez-Velazco, C; Solórzano-Figueroa, F; Araya-Vargas, M. 2009. Relación entre nematodos y la productividad del banano (Musa AAA) en Ecuador. Agronomía Mesoamericana 20(2):351-360. [ Links ]
Chindo, PS; Khan, FA. 1988. Relationship between initial population densities of Meloidogyne incognita race 1 and growth and yield of tomato (Lycopersicon esculentum Mill.). Pak J. Nematol. 6(2):93-100. [ Links ]
Di Vito, M; Ekanayake, HMRK. 1983. Relationship between population densities of Meloidogyne incognita and growth of resistant and susceptible tomato. Nematologia Mediterranea 11(2):151-155. [ Links ]
Ehwaeti, ME; Phillip, SM; Trudgill, DL. 1998. Dynamics of damage to tomato by Meloidogyine incognita. Fundam. appl. nematol. 21:627-635. [ Links ]
Ekanayake, MHRK; Di Vito, M. 1984. Effect of population densities of Meloidogyne incognita on growth of susceptible and resistant tomato plants. Nematologia Mediterranea 12(1):1-6. [ Links ]
El-Sherif, AG; Refaei, AR; El-Nagar, ME; Hagar, M;Salem, M. 2007. The role of eggs inoculum level of Meloidogyne incognita on their reproduction and hostreaction. African Journal of Agricultural Research 2(4):159-163. [ Links ]
Esparza, CC. s.f. Introducción al programa SPSS 13.0.Laboratorio de estadística. España, CSIC. 57 p. [ Links ]
Esparrago, RMG; Navas, A. 1995. Nematofauna fitoparásita asociada a cultivos hortícolas y tabaco en regadíos de Extremadura. Bol. San. Veg. Plagas 21:303-317. [ Links ]
Fawole, B; Mai, WG. 1979. Influence of plant age, light, intensity, nematode inoculums level, and their interactions on tomato growth and reproduction of Meloidogyne hapla. J. Nematol. 11:199-201. [ Links ]
Ferris, H. 1985. Density-dependent nematode seasonal multiplication rates and overwinter survivorship: a critical point model. J. Nematol. 17(2):93-100. [ Links ]
Hooper, DJ; Hallmann, J; Subbotin, SA. 2005. Methods for extraction, processing and detection of plant and soil nematodes. In Luc, M; Sikora, RA; Bridge, J. eds. Plant parasitic nematodes in subtropical and tropical agriculture. 2 ed. UK, CABI. p. 53-86. [ Links ]
INETER (Instituto Nicaragüense de Estudios Territoriales) 2011. Areas técnicas. Meteorología. Normas históricas (en línea). Consultado 22 dic. 2011. Disponible en http://www.ineter.gob.ni/index.html [ Links ]
Khan, RM; Khan, AM; Khan, MW. 1986. Interaction between Meloidogyne incognita, Rotylenchulus reniformis and Tylenchorhynchus brassicae on tomato. Revue de Nématologie 9(3):245-250. [ Links ]
Khan, H; Ahmad, R; Akhtar, AS; Mahmood, A; Basit, T; Niaz, T. 2000. effect of inoculum density of Meloidogyne incognita and plant age on severity of root-knot disease in tomato. Int. J. Agric. Biol. 2:360-363. [ Links ]
Kumar, S; Khanna, as. 2006. effect of neem-based products on the root-knot nematode, Meloidogyne incognita, and growth of tomato. Nematologia Mediterranea 34:141-146. [ Links ]
Mai, WF; Mullin, PG. 1996. Plant-parasitic nematodes: a pictorial key to genera. Ithaca, NY, Cornell University Press. 277 p. [ Links ]
McSorley, R; Dickson, DW; Candanedo-Lay, EM; Hewlett, TE; Frederick, J. 1992. Damage functions for Meloidogyne arenaria on peanut. J. Nematol. 24:193-198. [ Links ]
Mekete, T; Mandefro, W; Greco, N. 2003. Relationship between initial population densities of Meloidogyne javanica and damage to pepper and tomato in ethiopia. Nematología Mediterránea 31:169-171. [ Links ]
Moens, M; Perry, RN; Starr, JL. 2009. Meloidogyne species–a diverse group of novel and important plant parasites. In Perry, RN; Moens, M; Starr, JL. eds. Root knot nematodes. Wallingford, UK, CAB International. p. 1-17. [ Links ]
Morera, N; López, R. 1987. Evaluación de la virulencia de tres poblaciones de Meloidogyne exigua Goeldi, 1887, en el cafeto y de la resistencia de seis líneas de dicho cultivo a una base de esas poblaciones. Boletín de Promecafe 35:9-15. [ Links ]
Olthof, HA; Potter, JW. 1977. Effects of population densities of Meloidogyne hapla on growth and yield of tomato. J Nematol. 9(4):296-300. [ Links ]
Philis, J. 1990. The efficacy of nematode resistant tomato cultivars to Meloidogyne javanica under green house conditions. Nematologia Mediterranea 18:209-11. [ Links ]
Ploeg, T. 2000. Effects of amending soil with Tagetes patula cv. single gold on Meloidogyne incognita infestation of tomato. Nematology 2:289-493. [ Links ]
Ravichandra, NG. 2008. Plant nematology. I. K. International New Delhi, India, Publishing House. 693 p. [ Links ]
Salazar-Antón, W; Guzmán-Hernández, TJ. 2013. Nematodos fitoparásitos asociados al tomate en la zona occidental de Nicaragua. Agronomía Mesoamericana 24(1):27-36. [ Links ]
Sasser, JN; Carter, CC. 1982. Overview of the international Meloidogyne project rational, goals, implementation and progress to date. Proceeding of Conference on Root-knot nematodes Meloidogyne spp. (region 111). Brasillia, Brazil. p. 3-13. [ Links ]
Sayre, RM; Toyama, TK. 1964. The effect of root-knot nematodes on the yield of processing tomatoes. can. J. Plant Sci. 44: 265-267. [ Links ]
Seinhorst, HW. 1968. A model for the relation between nematode density and yield of attacked plants including growth stimulation at low densities. Reports of the eighth international symposium of nematololgy antibes. Wageningen, netherlands. 133 p. [ Links ]
Sikora, RA; Fernández, E. 2005. Nematode parasites of vegetables. In Luc, M; Sikora, RA; Bridge, J. eds. Plant parasitic nematodes in subtropical and tropical agriculture. 2 ed. Wallingford, UK, CABI Publishing. p. 319-392. [ Links ]
Shurtleff, MC; Averre III, CW. 2000. Diagnosing plant diseases caused by nematodes. The american Phytophatological Society. St. Paul, Minnesota, USA. p. 187. [ Links ]
Wallace, HR. 1971. The influence of the density of nematode populations on plants. Nematologica 17:154-166. [ Links ]
Zaki, A; Siddiqui, ZA; Iqbal, A; Mahmood, I. 2001. Effects of Pseudomonas fluorescens and fertilizers on the reproduction of Meloidogyne incognita and growth of tomato. Applied Soil Ecology 16:179-185. [ Links ]
*Correspondencia a:
2Wilber Salazar-Antón: Laboratorio de Fitopatología. Departamento de agroecología. Universidad nacional autónoma de Nicaragua-León. Nicaragua. Apartado postal n° 64. Teléfono:(505) 311-1779 / Fax:(505) 311-1780. wilber.salazar@ct.unanleon.edu.ni
3Tomás de Jesús Guzmán-Hernández: Instituto Tecnológico de Costa Rica, sede San Carlos. Apartado postal 159-7050, Florencia, Alajuela, Costa Rica. Tel: (506) 2475-5310. tjguzman@itcr.ac.cr
Recibido: 12 de junio, 2012. Aceptado: 28 de octubre, 2013


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}