











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
The genus Lepanthes Sw., a highly diverse genus of Neotropical orchids, with more than 1100 species, and new species being frequently discovered in Ecuador, is divided into two subgenera: Lepanthes and Marsipanthes (Baquero 2018, Baquero et al. 2018, Baquero et al. 2019, Karremans 2016, Luer & Thoerle 2011, Luer 1996, Thoerle & Hirtz 2015, Tobar et al. 2018, Zambrano & Solano 2019).
The genus Lepanthes has plants with ramicauls enclosed by lepanthiform sheaths, flowers with transversely expanded petals, fleshy or membranaceous, with two or three lobes, and the lip two or three lobed (Luer 1996). Most plants of Lepanthes subgenus Lepanthes have a complex lip with a body connecting a pair of blades which normally embrace the column, and a very small structure at the base of the lip called the appendix (Luer 1996, Blanco & Barboza 2005). In 1986, the subgenus Marsipanthes was proposed by Luer to accommodate three atypically saccate-flowered species (with the sepals forming a pouch). In 1996, in his monograph of the genus, Luer added four more species to subgenus Marsipanthes and presented a key to its species and sections (Luer 1986, 1996). All the species in the subgenus have more or less saccate flowers with fleshy instead of membranaceous petals, which are born from a peduncle lying on top of the leaf, with the successively flowered raceme reaching the tip or beyond (Luer 1996). In 2003, another species of Lepanthes is added to the Marsipanthes subgenus and described as L. niesseniae Luer. This is the only species of Marsipanthes in which the peduncle and flower raceme do not reach the tip of the leaf and the flowers are not saccate (Luer & Thoerle 2012).
Recently, in a poorly explored forest from north-west Ecuador protected by EcoMinga Foundation, a new and spectacular species of Lepanthes was discovered. The species belongs to subgenus Marsipanthes and shares, together with L. niesseniae¸ a peduncle and flower raceme which are shorter than the leaf and with very large plants for the subgenus. The new species is described here.
Taxonomic treatment
Lepanthes tulcanensis Baquero & M.F.Monteros, sp. nov. (Fig.1-4).
TYPE: Ecuador. Carchi: Reserva Dracula, 2043 m, 15 September 2018, Marco Monteros MFM201 (holotype: QCNE).
Diagnosis: Lepanthes tulcanensis is most similar to L. niesseniae, sharing the elliptical ovate leaves suffused with purple, the inflorescence with the flower shorter than the length of the leaves, and the shallower flowers compared to the rest of the species in the subgenus. Lepanthes tulcanensis differs from L. niesseniae in the upper lobe of the petals sub-rectangular to oblong, convex, slightly concave towards the central vein, covered with a velvety texture, and the lower lobe narrowly triangular, with the apex hispid vs. the upper lobe of the petals oblong and bosselated, and the lower lobe curved and rounded at the apex, the lip velvety with the appendix triangular and acute vs. the bossellated and verrucose lip with a narrowly oblong appendix in L. niesseniae.
Plant epiphytic, sympodial, caespitose herb up to 40 cm tall. Roots ca. 0.8 mm in diameter. Ramicauls slender, arcuate to horizontal, elongated, thin, 25-35 cm long, enclosed by long-acuminate, coarse, apiculate at the margins, loose lepanthiform sheaths, 1.8-2.5 long. Leaf green suffused with purple, thinly coriaceous, conduplicate and reticulate, ovate, the base slightly cuneate, the apex 2.5 mm, long-acuminate, 10-13 × 5.5-7.0 cm. Inflorescence congested, 1 to several, successively, many-flowered racemes up to 30-50 mm long, born on top of the leaf by a slender peduncle 20-40 mm long; floral bracts oblique, acuminate, 17-20 mm long; pedicels 5 mm long. Ovary costate, 5 mm long. Flower with sepals translucent yellow suffused with orange, with a dark red blotch at the dorsal sepal and three red blotches towards the center of each lateral sepal and petals yellow, yellowish-green, suffused with brown towards the margins of the upper lobe, column red, and lip yellow suffused with yellowish-green, sanguine red at the margins of the lobes, anther cap rose. Sepals carinate along the veins on the abaxial surface. Dorsal sepal broader and longer than the lateral sepals, broadly ovate, shallowly concave, 7-veined, minutely ciliate at the margin, caudate, 9.3 × 8.5 mm, including a 3.2 mm long tail, connate to the lateral sepals for 2 mm. Lateral sepals broadly ovate, 4-veined (each lateral sepal), oblique, minutely ciliate at the margins, caudate, connate 4.5 mm into a shallow cup, 9.5 × 11.0 mm, including a 3 mm long tail, each individual sepal free for ca. 5 mm. Petals transversely bilobed, velvety, the upper lobe sub-rectangular to oblong, convex, slightly concave towards the midline and apex of the petal, rounded and hispid at the apex, 1.7 × 0.8 mm long, the lower lobe narrowly triangular, slightly convex, obtuse and hispid at the apex, 1.5 × 0.6 mm long, the upper lobe rounded and the lower lobe straight where the lobes meet at the midline forming an acute angle between them. Lip velvety, transversely bilobed, with the lobes broadly cuneate, auriculate, oblique, flanking the column, forming a narrow body, connate to the base of the column, the frontal part of each lobe hispid towards the anther, with a triangular, acute, bristled appendix with an echinaceous and globose structure at the sinus, 1.7 × 2.0 mm. Column subterete, 1.3 mm long, the anther apical and stigma ventral, the viscidium bent upwards, the apex of the rostellum elongate, acute. Anther cap cucullate, obovate, 0.4 mm long. Pollinia 2, yellow, pyriform, attached to a detachable viscidium, 0.4 mm long. Capsule not observed.
Habitat and ecology:
Lepanthes tulcanensis is so far endemic to a relatively small area in northwestern Ecuador, close to the border with Colombia. A population of three plants from a first locality were found growing in an extremely moist cloud-forest at an unexplored area within the Dracula Reserve at around 2000 m in elevation. One of the plants was collected and a division of the plant was used for the holotype. The rest of the plant was taken to the Quito Botanical Garden for ex situ culture. A few months later, another robust plant was found growing in similar conditions 11 km from the first group of plants. At both localities, the plants of L. tulcanensis were found growing low (2 m) off the ground on wide trunks where high moisture and dappled light was constant.

A. Lepanthes tulcanensis (leaf and flower view, in situ).
B. Lepanthes tulcanesis (flower frontal view).
C. Lepanthes niesseniae (flower frontal view).
D. Lepanthes quadricornis (flower frontal view).
E. Lepanthes ribes (flower frontal view).
F. Lepanthes lucifer (flower frontal view).
Photographs by Marco F. Monteros (A, B and E from the holotype MFM 201, QCNE), and Luis E. Baquero (C, D and F).
Figure 1 Comparison of the most similar species and other species of the Marsipanthes subgenus to Lepanthes tulcanensis Baquero & M.F.Monteros.
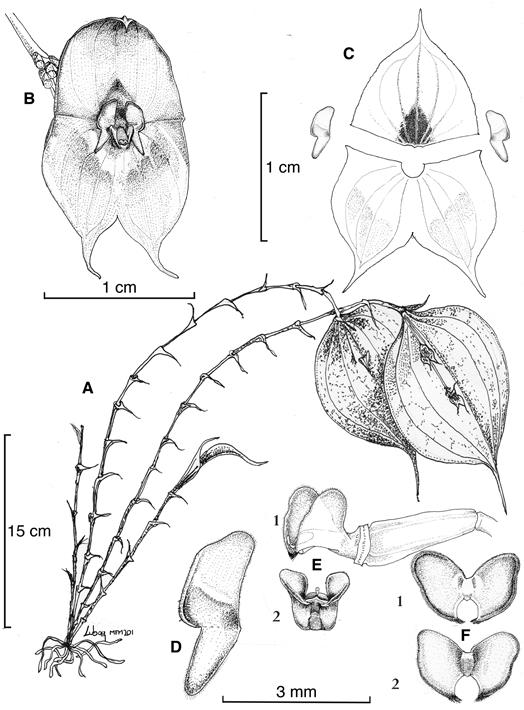
A. Habit.
B. Flower.
C. Dissected perianth.
D. Petal.
E1. Lateral view of lip and ovary.
E2. Frontal view and lip column.
F1. Lip in expanded position, adaxial view.
F2. Lip in expanded position, abaxial view.
Drawn by Luis E. Baquero from the plant that served as the holotype (MFM 201, QCNE).
Figure 2 Illustration of Lepanthes tulcanensis Baquero & M.F.Monteros.

Paratype
Ecuador. Carchi: Reserva Dracula, 2043 m, 21 October 2018, Marco Monteros MFM202 (paratype: QCNE-Spirit).
Phenology
This species has always been observed blooming at its habitat, in different months. A plant is cultured at the Quito Botanical Garden but it seems to need higher moisture for the leaves to develop in optimal condition, and no flowers have appeared since the plant was taken to Quito.
Lepanthes tulcanensis is a member of subgenus Marsipanthes considering the dorsal sepal with seven veins and lateral sepals with more than four veins-eight in total-, thick and fleshy petals, the bilobed lip without blades, the shallowly cupped flowers and the inflorescence with a long peduncle and congested raceme which rests at the adaxial side of the leaf. This species is, perhaps, the most atypical species of the subgenus Marsipanthes since it has much larger plants (close to 40 cm, including the leaf) and bigger leaves (with a long acuminate apex) than any other member of the subgenus. From a distance, L. tulcanensis resembles L. rhodopylla Schltr. due to the large plants with dark, reticulated, acuminate leaves. Nevertheless, the flowers are completely different not only from L. rhodopylla but from any other species in the genus, based on the inflorescence which is born at the adaxial side of the leaf, the dorsal sepal with 5 veins, and 8-veined synsepal, the bilobed lip without blades, the shallowly cupped flowers and the fleshy petals (Fig. 1 - - 4).
Conservation status
Two healthy populations have been found over the years near the location where it was initially discovered in Carchi province in north- west Ecuador. Nevertheless, a few plants have perished since the first population was discovered in September of 2018. These plants apparently died due to the harsh summer of 2019. Another threat for the populations of this species and the species itself (as far as we know about it) is that the area where all the populations of this species are known from is now affected by mining activity. We suggest to place it under the IUCN category “Critically Endangered - CR”, under criteria B1, considering an extent of occurrence of less than 100 square km with only two known locations (a) and a continued decline observed (b) since the species is strongly affected by mining activity and, it might perish in the next years if mining activity is not controlled.

A. Petals and lip, frontal view.
B. Petals and lip, – view.
C. Lip and sinus, frontal view.
D. Apex of the lip and the sinus, Frontal view.
E. Hispid hairs at the tip of the lobes from the lip.
Photographs by Marco F. Monteros (A-B), and Luis E. Baquero (C-E from paratype MFM 202, QCNE).
Figure 4 Details of Lepanthes tulcanensis Baquero & M.F.Monteros.

