










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkMedicina Legal de Costa Rica
On-line version ISSN 2215-5287Print version ISSN 1409-0015
Med. leg. Costa Rica vol.27 n.2 Heredia Sep. 2010
Revisión Bibliográfica
Actualización de la Fisiología Gástrica
David Rodríguez Palomo *
Andrea Alfaro Benavides +
* Especialista en Anatomía, Departamento de Anatomía, Universidad de Costa Rica, daropa@gmail.com
+ Médico general, Universidad de Costa Rica
Resumen
La fisiología gástrica constituye una compleja y dinámica interacción entre estructuras anatómicas, sus secreciones, el ambiente circundante y los factores exógenos aportados por el individuo. Este trabajo pretende hacer una breve revisión actualizada sobre la fisiología de la secreción y la motilidad gástricas. Se expondrá la función de las principales células de la mucosa gástrica, sus secreciones y aquellas sustancias, hormonas y neurotransmisores que las estimulan, modulan o inhiben, haciendo especial énfasis en los mecanismos de producción del acido gástrico. Además, se tratará específicamente el tema de la motilidad gástrica con idéntica metodología.
Palabras clave
Ghrelina, neurotensina.
Abstract
Gastric physiology constitutes a complex and dynamic interaction between anatomic structures, their secretions, the environment around and the exogenous factors provided by the individual. This work pretends to offer a brief actualized literature review about the physiology of the gastric secretions and gastric motility. We will expose the function of the main gastric mucosa cells, its secretions and the substances, hormones and neurotransmitters that enhance, modify and inhibit them, making special emphasis in the gastric acid secretion. Moreover, the subject of gastric motility, specifically, will be explained with identical methodology.
Key words
Ghrelina, neurotensin.
Introducción
El estómago de los vertebrados realiza una variedad de funciones que incluyen el almacén de alimentos, la exposición de los alimentos al ácido que secreta, proveer una barrera que prevenga el paso de microorganismos al intestino y mantener una mucosa protectora contra agresiones endógenas y exógenas(1).
El sistema nervioso entérico (ENS) y diferentes formas de percepción sensorial inician y sostienen el proceso coordinado de motilidad gastrointestinal, circulación, absorción, secreción exocrina y endocrina e inclusive la saciedad, siendo así sus principales blancos las células del músculo liso, las células de la mucosa secretora, aquellas de la microvasculatura y las células inmunomoduladoras e inflamatorias(2,3,4).
Secreciones gástricas y su control
Células y productos asociados a la generación de ácido
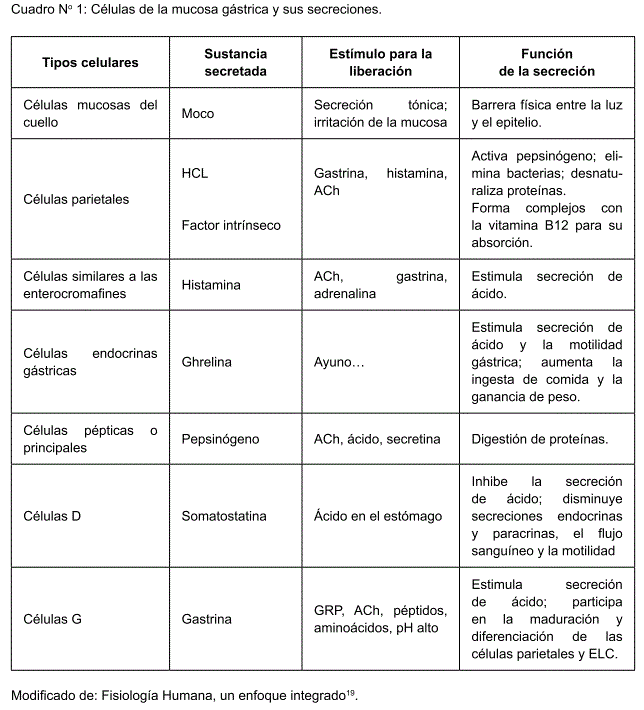
El estómago es rico en células productoras de péptidos hormonales, a saber, las células similares a las enterocromafines (ELC) que secretan histamina, las células D productoras de somatostatina, las células similares a A que producen grelina y obestatina, las células D1/P cuyos productos son desconocidos, las células enterocromafines secretoras de serotonina y las células G encargadas de la producción de gastrina(5).
Células similares a las enterocromafines (ELC): se localizan en la mucosa oxíntica en proximidad directa con las células parietales6,7. Son capaces de autoreplicarse bajo la influencia de la gastrina(7).
Histamina: es el principal estimulador paracrino de la secreción de ácido(7). La histamina es sintetizada en la mucosa gástrica mediante la histidina decarboxilasa (HDC) en las células similares a las enterocromafines (ELC)(6,7,8,9) así como los mastocitos5 y liberada solamente de las células ELC en respuesta a la gastrina, que actúa en los receptores CCK2 de éstas(5,6,7,8). Estimula los receptores H2 de las células parietales para la secreción gástrica(5,6,7,8,9).
Ghrelina: es un péptido liberador de la hormona de crecimiento, que ejerce múltiples acciones en la fisiología gastrointestinal, como el aumentar la ingesta de comida y la ganancia de peso, estimular la secreción de ácido y la motilidad gástrica(9,10,11). Se encuentra en las células endocrinas gástricas y el hipotálamo. Su liberación ocurre en períodos de ayuno y se cree que ejerce su acción al inducir la liberación de histamina mediante activación vagal nerviosa, lo que conduce a un aumento en la secreción de ácido por las células parietales. Su acción puede ser abolida con atropina o vagotomía cervical bilateral(9).
Células D: están presentes en la mucosa antral y oxíntica, secretan somatostatina(5,6,8,12).
Somatostatina: en el tracto digestivo, es producida por células D endocrinas y paracrinas y nervios entéricos. Su efecto es siempre inhibitorio, disminuyendo las secreciones endocrinas y paracrinas, el flujo sanguíneo y la motilidad(6,7,8,12,13). Su vida media es de menos de 3 minutos(12).
Aún cuando tiene cierto efecto sobre las células parietales, su mayor efecto lo ejerce inhibiendo la liberación de histamina y gastrina. Su secreción es estimulada por el acido gástrico y la gastrina(6,7,8,13). La activación colinérgica de la secreción gástrica promovida por la distensión del estómago, se debe en parte a la supresión de la liberación de somatostatina. Su secreción también se ve afectada por estimulo neural, suprimida por activación colinérgica e incrementada por PIV(13).
Células G: se hallan en la región antral del estómago y liberan gastrina(6,14).
Gastrina: secretada principalmente por las células G del antro gástrico(6,15), también se produce en el intestino delgado y páncreas. Es el producto de un único gen en el cromosoma 17, siendo su precursor la preprograstrina. Es liberada en respuesta a proteínas, péptidos y aminoácidos de la dieta(15).
Es el mayor regulador hormonal de la secreción ácida; estimula la secreción de ácido al unirse al receptor CCK2 de las células parietales y las células ELC(6,7,9,14,15). Su liberación es claramente influenciada por el pH gástrico, un pH alto estimula fuertemente su secreción, mientras un pH muy ácido la inhibe. Otras dos hormonas, el péptido liberador de gastrina y la somatostatina, estimulan e inhiben su liberación, respectivamente(6,7,14,15).
También desempeña una función crítica en la proliferación, organización y fisiología de la mucosa gástrica al participar en la maduración y diferenciación terminal de éstas las células parietales y las células ELC, principales responsables de la secreción de ácido. Asimismo, influye en la migración de las mismas a lo largo del axis de la glándula gástrica(14).
Células parietales: tienen a su cargo la producción de HCL (ácido clorhídrico) y pepsinógeno. Expresan receptores H2, M3, CCK2 Y sst2, los cuales regulan la producción del ácido(5,15,16).
La función de las células parietales no está regulada solamente por hormonas (gastrina, CCK) y mensajeros paracrinos (histamina, somatostatina) sino también por neurotransmisores de las neuronas entéricas, a saber, ACh, catecolaminas y neuropéptidos como el péptido pituitario activador de la adenilato ciclasa (PACAP), el péptido intestinal vasoactivo (VIP) y la galanina(5,7). La principal, la Ach, expresa 5 subtipos de receptores, M1 en las células líder y mucosas de la superficie, M2 y M4 en las células D, M3 en las células parietales y M5 en fibras nerviosas entéricas posganglionares. De todos estos estímulos para la producción de ácido, el principal es la histamina vía las células ELC(5,15).
Prostaglandinas: son factores autocrinos que inhiben la secreción acida estimulada por histamina y la liberación de histamina estimulada por gastrina. Son generadas en el epitelio y lámina propia gástricos, siendo sus principales productores los macrófagos y las células endoteliales capilares(6,7).
TGF-alfa y PYY: el factor de crecimiento transformador alfa es un factor autocrino presente en las células parietales que inhibe la secreción ácida gástrica. El péptido YY es liberado postprandialmente de células ileales y colónicas e inhibe las fases cefálica y gástrica, además se une a receptores de las células ELC e inhibe la liberación de histamina estimulada por gastrina(6).
Fases y regulación de la secreción de ácido
El ácido facilita la digestión de proteínas y la absorción de calcio, hierro y vitamina B12, y además ejerce una acción protectora contra microorganismos exógenos(16).
Fases: La secreción de ácido ha sido clásicamente dividida en tres fases interrelacionadas, una cefálica, otra gástrica y la intestinal. En la fase cefálica, la simple imaginación o percepción visual de la comida es capaz de aumentar el estímulo vagal e iniciar la actividad gastrointestinal, así como el oler, probar o tragar. La fase gástrica es producto del efecto químico de los alimentos y la distensión gástrica, donde la gastrina parece ser su principal mediador. La última, la intestinal, es responsable solo de una pequeña proporción de la secreción de ácido en respuesta a la comida, y sus mediadores aún están en controversia(3,6,7,16).
Con respecto al papel de los distintos alimentos en la producción de ácido, las proteínas y los aminoácidos estimulan la secreción de ácido, mientras los carbohidratos y las grasas lo inhiben4,6,7. El efecto de los carbohidratos requiere de la exposición intestinal a los mismos y su mecanismo de acción es incierto. La grasa estimula la liberación de CCK, un potente inhibidor de la secreción ácida(6). Los L-aminoácidos y no los D-aminoácidos pueden inclusive modular la secreción gástrica en ausencia de los intermediarios hormonales clásicos como la gastrina e histamina, esto por ser potentes activadores alostéricos del receptor sensador de calcio (CaSR) expresado en las células parietales y las células G(17).
La secreción ácida gástrica está regulada mediante la interacción de señales endocrinas, paracrinas y neurocrinas por al menos tres vías mensajeras principales: (a) gastrina-histamina, (b) CCK-somatostatina y (c) neural, mediante la ACh y neuropéptidos. También existen comunicaciones entre los receptores H2 y M3, y CCK2 y sst2 de las células parietales, probablemente a través del traslape de segundos mensajeros como el cAMP y la fosfolipasa C. Esta comunicación es indispensable, como lo demuestra el hecho de que en ausencia del receptor M3 en ratones mutilados, las células parietales no alcanzan su capacidad secretora máxima pese al contacto con estimuladores vagales y la integridad de la vía gastrina-histamina (lo mismo no sucede con los receptores H2 y sst2)(5,6,7,16).
Interacción de las vías: (a) La gastrina es liberada de las células G y actúa en los receptores CCK2 de las células ELC, estimulando la producción de segundos mensajeros como el fosfatidil inositol y el calcio, activando la pkC y aumentando la expresión de ARNm de la histidin carboxilasa (HDC), acelerando la liberación y síntesis de histamina, que actuará en los receptores H2 de las células parietales y estimulará la secreción de ácido. (b) La CCK circulante actúa en los receptores CCK1 de las células D, quienes liberan la somatostatina que inhibirá las células G, las ELC y las parietales, al actuar en el receptor sst2, inhibiendo la vía gastrina- histamina(5,7,15). En ratones doblemente bloqueados para la producción de gastrina y CCK, se observó que las células parietales seguían siendo capaces de producir ácido en respuesta al estímulo vagal7. (c) La ACh actúa en los receptores M3 de las células parietales estimulando la secreción de acido y, en M2 y M4 de las células D para inhibirla(5,7,15). Además la adrenalina y NA actúan en receptores β2 de las células ELC movilizando histamina, mismo resultado que se logra mediante el PACAP (receptor PAC1) y el VIP (receptor VAPAC2). La galanina inhibe la vía de la gastrina-histamina, también en las células ELC (receptor Gal1)(7).
Cuando la histamina se une a su receptor H2 en la membrana basolateral de las células parietales, la resultante cascada de eventos incluye un aumento del calcio citoplasmático y elevados niveles de AMPc, que disparan el eflujo de iones cloruro e hidrogenión de la superficie apical de la célula mediante un canal de conductancia de cloruro y la H-K-ATPasa recién traslocada(6,16,17). En su estado de reposo, las células parietales están llenas de vesículas secretoras que coalescen ante la estimulación para fusionarse con los canalículos que drenan hacia el lumen apical resultando en la secreción de HCl. El cese de la secreción de ácido está asociado a la reinternalización de la bomba de protones, mediado por la cola citoplasmática de la unidad beta de la bomba6,7,16.
La formación de ácido incluye los siguientes pasos: (a) el ión cloruro es activamente transportado del citosol hacia el lumen del canalículo y lo opuesto sucede con los iones de sodio que son transportados activamente fuera de los canalículos hacia el citoplasma, creando un potencial negativo que conduce a la difusión de iones potasio positivamente cargados y un número menor de iones sodio del citoplasma al canalículo. En resumen, KCl principalmente y NaCl entran al canalículo. (b) El agua se disocia en iones hidrógeno e hidroxil en el citoplasma parietal, los iones hidrogeno son activamente secretados hacia el canalículo en intercambio por iones potasio mediante la H-K-ATPasa. Los iones de sodio son activamente reabsorbidos por otra bomba de manera que la mayoría del sodio y potasio difundidos hacia el canalículo son reabsorbidos dentro del citoplasma. El HCl es entonces secretado por el extremo abierto del canalículo hacia el lumen glandular. (c) El agua pasa hacia el canalículo mediante osmosis debido a los iones ya excretados hacia el canalículo. Así la secreción final tiene contiene agua, HCl (150-160mEq/L), KCl (15mEq/L) y una pequeña concentración de NaCl. (d) Por último, el dióxido de carbono, ya sea formado durante el metabolismo celular o proveniente de la sangre, se combina bajo la influencia de la anhidrasa carbónica con los iones hidroxil para formar iones bicarbonato, los cuales difundirán hacia el liquido extracelular en intercambio por iones cloruro6,7,8,16. Clásicamente se ha dicho que en los adultos mayores, una reducción en la función de las células parietales condiciona la disminución en la secreción de ácido y la elevación del pH gástrico, sin embargo, otros estudios han demostrado que esto es propio de pacientes con sintomatología gastrointestinal, pues en adultos mayores con morbilidades varias sin síntomas gástricos, hasta un 90% han sido capaces de acidificar el contenido gástrico aún en su estado basal(18).
Secreción de moco
Una constantemente renovada capa de moco cubre el tracto gastrointestinal y constituye una barrera entre el contenido luminal y la mucosa. El moco es secretado por las células epiteliales y sus constituyentes principales son grandes glicoproteínas, mucinas y agua(6,8,12). En el estómago se han identificado dos tipos distintos de mucinas, la MUC5AC secretada por las células mucosas superficiales y la MUC6, secretada por las células del cuello, así como las siguientes mucinas transmembrana: MUC1, MUC4, MUC16, cuyo rol parece estar ligado a señalización de transducción y fenómenos de adhesión(12).
De igual manera, el moco está dispuesto en dos capas, una externa aguada y otra interna firmemente adherida al epitelio (su remoción lo dañaría). La primera se encarga de la unión con agentes nocivos y nitritos y de la liberación constante de NO; ésta se mezcla con los alimentos y se desprende. La segunda desempeña un papel más importante para la protección de la mucosa del ácido corrosivo, pues mantiene un pH neutral (el del pH yuxtamucosal), retarda la difusión retrograda de iones hidrógeno y mantiene el bicarbonato secretado por el epitelio. La MUC5AC es el componente principal de ambas capas, sin embargo, el estimulo para el engrosamiento de ambas capas es distinto, ambas responden al estímulo de la PGE2, mientras sólo la capa interna responde a la donación luminal de NO; las razones aun no han sido claramente esclarecidas(12).
Otras secreciones
Pepsinógeno: Es secretada por las células pépticas y mucosas de las glándulas gástricas. Existen distintos tipos de pepsinógenos sin distinción de su actividad (hasta ahora) y todos son inicialmente inactivos, pero al contacto con el acido clorhídrico se activan formando la pepsina. Esta es una enzima proteolítica muy activa a un pH óptimo ente 1.8 y 3.5, pero completamente inactiva por encima de pH 5(16). La Ach y péptidos de la familia gastrina-CCK estimulan la secreción de pepsinógeno(6).
La pepsina se encarga de digerir los productos proteicos de la dieta, principalmente aminoácidos, potentes estimuladores de la liberación de gastrina y secreción de ácido(4,6,16).
Factor intrínseco: El factor intrínseco se produce en las células parietales junto con la secreción de HCL y es esencial para la absorción de la vitamina B12(1,16).
La cobalamina es cofactor y coenzima para muchas reacciones bioquímicas, incluyendo la síntesis de ADN, la síntesis de metionina a partir de la homocisteína y la conversión de propionil en succinil CoA del metilmalonato. El factor intrínseco se une a la cobalamina para permitir su absorción en la mucosa ileal mediante el receptor de cubilina de los enterocitos y permitir su transporte con la transcobalamina(1,6,8,16). La deficiencia del factor intrínseco, debida a gastrectomía total o anemia perniciosa (enfermedad autoinmune caracterizada por destrucción de la mucosa gástrica, especialmente fúndica) junto con secreción deficiente de HCl o gastrina y secreciones biliares y pancreáticas, suman el 75-80% de las causas de deficiencia de cobalamina en adultos mayores(1).
Motilidad y vaciamiento gástricos
Las principales funciones de la motilidad gástrica son la acomodación y almacenamiento de la comida ingerida, la trituración de partículas sólidas junto son su mezcla para una máxima exposición a las enzimas digestivas y la liberación de los mismos en una forma armónica hacia el duodeno8,16,19. Además el esfínter esofágico inferior y el pilórico regulan el tránsito a través de ellos y evitan el reflujo. Más aún, ya se conoce del papel de esta mecánica en la generación de saciedad(8,16).
A lo largo del tracto digestivo, los patrones de actividad motora difieren según correspondan al periodo de llenado o posterior a la ingesta. En el primer periodo, la actividad motora está altamente organizada en secuencias cíclicas contráctiles conocidas como complejo motor migratorio (MMC)(8,19). El MMC consiste de tres fases distintas consecutivas que migran a lo largo del intestino delgado. La fase I inicia con un período de quiescencia motora, la cual es seguida de una fase de aparentes contracciones erráticas e irregulares (fase II) y culmina con contracciones fásicas ininterrumpidas (fase III). Cada ciclo tarda entre 1 y 2 horas aproximadamente. En el estómago, los patrones del MMC tienden a comenzar y terminar de forma simultánea en todos los sitios, en vez de ser un proceso de propagación, como sucede en el intestino delgado(8). Además, los patrones de motilidad del estómago son región específicas; el fondo y el cardias (regiones marcapaso) generan contracciones tónicas, que se mantienen durante minutos u horas, mientras el cuerpo distal y el antro exhiben actividad motora fásica, con ciclos de contracción-relajación que solo duran algunos segundos(19,20).
Las contracciones fásicas se asocian a ciclos espontáneos de despolarización y repolarización conocidos como potenciales de onda lenta debido a su frecuencia19,20 (3 ondas/min para el antro gástrico, 12 ondas/min en el duodeno, 8 ondas/min en el íleo y 6-10 ondas/min en el colon)(19) respecto a las células marcapaso cardiacas (60-90 ondas/ min)(20). Cuando un potencial de onda alcanza el umbral, los canales de Ca+ voltaje dependientes se abren y la célula dispara uno o más potenciales de acción generando la contracción del musculo liso. Más potenciales de acción se disparan y mayor es la fuerza y duración de las contracciones cuanto mayor es la duración de la onda lenta. Estas ondas lentas se originan en células de musculo liso modificadas, llamadas células de Cajal(19,20), ubicadas en los bordes mientérico y submucoso de las fibras circulares(8,19).
Estómago proximal
Como se menciono antes, el estomago proximal sufre contracciones y relajaciones lentas, así durante la deglución, el fondo se relaja unos 15 minutos luego de la ingestión de alimentos para luego retornar a su tono basal (mediada vagalmente)20), a la que sigue una prolongada acomodación, neuralmente mediada por mecanorreceptores de la pared (el estómago tiene una capacidad de 1-2 litros sin generar distensión(8,20). Durante la intensa actividad gástrica contráctil, el esfínter esofágico inferior conserva un tono basal aumentado y exhibe contracciones fásicas superimpuestas para prevenir el reflujo, pues ondas de gran amplitud de un ciclo por minuto, son llevadas a cabo. Sin embargo, una verdadera actividad rítmica ocurre sólo en el antro distal en la fase III(8). El tracto gastrointestinal puede responder a diferentes estímulos sensoriales gracias a su quimio, termo y mecano sensibilidad21, de ahí que toda la dinámica contráctil varíe según los mismos.
Factores que influyen en el vaciamiento gástrico: además de la edad y el género, el volumen, la osmolaridad y la densidad calórica juegan un papel clave en el vaciamiento gástrico3,16,20. En general líquidos inertes se vacían rápido con un promedio entre 8 a 18 min. Un bolo de 300cc de sustancia salina será evacuado el doble de rápido que una carga de 150cc. Si en promedio se liberan aproximadamente 200kcal/h hacia el duodeno, líquidos con alto contenido calórico se vacían más lentamente que unas pocas calorías por unidad de volumen. Un aumento en la osmolaridad disminuye la respuesta a la contractibilidad en el intestino delgado. Las características de los nutrientes por sí mismas también regulan el vaciamiento, así vemos como el aminoácido L-triptófano (precursor de la 5-HT) retrasa el vaciado y el efecto de los triglicéridos en la motilidad es dependiente de la longitud de la cadena de ácidos grasos pues varían en su habilidad para liberar CCK. La acidez gástrica es igualmente importante, una disminución de la acidez retarda el vaciamiento de líquidos y sólidos. Hasta la temperatura es importante pues el frío inhibe el vaciado(16,20). Con la resección quirúrgica del fondo o una fundoplicatura, la presión intra gástrica aumenta y el vaciamiento se acelera. Si el atrio es resecado, la fase inicial de vaciado de líquidos también se acelera, sugiriendo que tanto el estómago proximal como el distal juegan un rol en el vaciamiento de los líquidos, al igual que el duodeno(20).
Agentes neurohumorales relajan el estómago proximal, entre ellos la CCK, secretina, PIV, gastrina, somatostatina, dopamina, glucagón, péptido insulinotrófico dependiente de glucagon (GIP) y bobesina(3,4,20). La motilina, involucrada en la regulación interdigestiva y postprandial mediante la estimulación del MMC(19,22), y la TRH aumentan la presión en el fondo(3,4,20).
El SNC juega un papel no menos importante, el estrés mental prolonga la periodicidad del MMC, el hambre aumenta la actividad motora fásica, el miedo y la depresión reducen las contracciones gástricas así como el frío y la isquemia, entre otros4. En general, las alteraciones motoras más comunes debido al estrés son el retraso en el vaciamiento gástrico y la aceleracióndel tránsito colónico(23). Los mediadores de la interacción cerebral-gástrica han sido evaluados intensamente, entre ellos los péptidos cerebrales de acción central, la TRH (hormona liberadora de Tirotropina) y el CRF (factor liberador de corticotropina) principalmente(4,23). La infusión intraventricular de TRH induce una rápida y prolongada respuesta contráctil, acelerando el vaciamiento gástrico; su efecto es abolido por vagotomía y la aplicación de atropina, y respecto a su localización podemos decir que gran parte de la TRH medular total se encuentra en el núcleo motor dorsal del vago y sus receptores se hallan en neuronas gástricas vagales preganglionares. La inyección de CRF en el líquido cerebroespinal inhibe la motilidad gastrointestinal, disminuyendo el vaciamiento gástrico; la Atresina, un derivado de la CRF con una actividad intrínseca muy baja y gran afinidad por los receptores CRF2 β (receptores de ubicación periférica), previene estas alteraciones propias del estrés23. Otros mediadores como la CCK, opiáceos, bombesina, taquikinas, somatostatina, Factor atrial natriurético, GABA y calcitonina retrasan el tránsito gástrico(4,20).
Estómago distal
Las contracciones rítmicas sincronizadas del estómago distal son controladas por señales eléctricas generadas en un marcapaso en la curvatura mayor y siempre están asociadas con las ondas suaves (aunque éstas persisten en ausencia de la actividad contráctil activa). La motilidad antral de llenado es cíclica y es llevada a cabo mediante el MMC en un promedio de 100min. El esfínter pilórico, durante la fase III del MMC se mantiene abierto y el contenido gástricos pueden ser evacuado hacia el duodeno.
Bajo condiciones de llenado, el píloro exhibe una compleja respuesta motora con prolongados períodos de cierre durante el mezclado y la retropulsión duodenal(20).
Las vías neurohumorales también afectan la actividad distal, las contracciones antrales son estimuladas por la ACh, CCK, bobesina y motilina (4,20,21); mientras la secretina, somatostatina, glucagon, GIP, TRH, neurotensina y PGE2 la inhiben y el PIV y el NO la disminuyen(4,20). El NO, PIV, PGE1, galanina y 5-HT son mediadores inhibitorios del esfínter pilórico; la ACh, SP y CCK son mediadores excitatorios. La distensión, estimulación química y osmolaridad del duodeno activan vías excitatorias ascendentes hacia la región antro-pilórica y así enlentecen el vaciamiento gástrico(20).
A modo de resumen, podemos agrupar algunas sustancias neurohumorales según su efecto dominante in vivo sobre la contractibilidad del musculo liso gastrointestinal de la siguiente manera. Con efecto estimulante: ACh, adenosina, bobesina, CCK, GRP (polipéptido liberador de gastrina), histamina, serotonina motilina, Neuroquinina A, opioides, PGE2, TRH. Inhibidores: CGRP (péptido regulador del gen de calcitonina), GABA, Galanina, Glucagon, NYP (neuropéptido Y), Neurotensina, NO, PACAP, PHI (péptido histidina isoleucina), PYY, Secretina, Somatostatina y VIP(20).
Respecto a la 5-Hydroxitriptamina o mejor conocida como 5-HT o Serotonina, sabemos que es liberada de las células enterocromafines(20,24), está presente en neuronas entéricas(20,21) y regula la función gastrointestinal de manera excitatoria e inhibitoria según su localización anatómica y las especies evaluadas en los estudios, además del subtipo de receptor (5-HT1 al 5-HT7) presente(20,24) y el tipo de agonista y/ o antagonista empleado en el estudio(21,24).
Propiamente en el estómago, los receptores 5-HT4 están presentes en el fondo, cuerpo y antro, y sus agonistas, tal como la Cisaprida, Metoclopramida, estimulan la actividad motora evaluada en estado de ayuno(20,21,24). La Cisaprida además facilita la acomodación gástrica en el periodo postprandial(21). La Buspirona y el Sumatriptan, agonistas 5-HT1, activan neuronas inhibitorias y suprimen la fase III gástrica, retrasan el vaciamiento, aumentan la relajación antral y fúndica y disminuyen la motilidad antral postprandial(20,21). El receptor 5-HT3 también tiene cierta actividad en el vaciamiento gástrico(20).
La ghrelina induce el MMC(10,11) y acelera el vaciamiento gástrico(9,10,11) funciones propias de la motilina(22) con quien comparte una gran similitud estructural aunque muy poca afinidad con los receptores de ésta pese a que la congruencia entre los receptores de uno y otro péptido varía entre un 44% y 87%, dejando amplias dudas respecto a una posible reacción cruzada que no se ha logrado documentar(10,22). Sus receptores están presentes en neuronas mientéricas, vagales y centrales, por ejemplo, aferentes vagales y en el ganglio nodoso (vías central y periférica), y su efecto in vivo puede ser bloqueado mediante vagotomía(10,11).
Una importante diferencia entre ambos péptidos lo constituye la concentración plasmática necesaria para ejercer un efecto apreciable en los estudios, pues se necesita hasta una relación de 6:1 de ghrelina vs. motilina para inducir el MMC(10). Al respecto cabe destacar que sus niveles plasmáticos tiene un patrón similar al de la leptina, con un pico nocturno durante el ayuno y mínimas contracciones matutinas tras el desayuno; su efecto se ha documentado solamente en periodos de ayuno(11).
Defensas gástricas
Los principales portales de ingreso de patógenos son la piel, el epitelio gastrointestinal, el respiratorio y el tracto urogenital. El epitelio gástrico no es una barrera estática, existe una comunicación compleja y dinámica entre los patógenos y el mismo. Dentro de los factores nocivos endógenos están el HCl, el pepsinógeno, la pepsina y sales biliares; extrínsecos tenemos medicamentos, alcohol y bacterias(6,25).
El sistema de defensa de la mucosa se puede dividir en tres niveles: preepitelial, epitelial y subepitelial. La primera línea de defensa es una capa de moco y bicarbonato que actúa como barrera fisicoquímica e impide la difusión de iones y moléculas como la pepsina. La superficie epitelial brinda su defensa mediante la producción de moco, los transportadores iónicos que mantienen el pH intracelular, la producción de bicarbonato y uniones estrechas intracelulares. Si estas células fallan, un proceso de migración regulado por los factores de crecimiento epidermal (EGF), transformador (TGF) y fibroblástico (FGF) restituirá la región dañada; cuando se regeneran las células participan el EGF y TGF-a y en la angiogénesis el FGF y el factor de crecimiento vascular endotelial (VEGF). El nivel subepitelial lo constituye un sistema microvascular en la capa submucosa, que suministra bicarbonato, micronutrientes y oxígeno, y además elimina productos tóxicos metabólicos. Las prostaglandinas también tienen un papel crucial como parte del sistema de defensa y reparación(6).
Recibido para publicación: 24 de enero 2010 Aceptado: 15 de abril de 2010
Referencias
1. Andrés, E., Loukili, N., Noel, E., Kaltenbach, G., Abdelgheni, M., Perrin, A., Noblet, M., Maloise, F., Schilienger, J.P., Blicklé, J. (2004). Vitamin B12 (cobalamin) deficiency in elderly patients. CMAJ, 171, 251-259 [ Links ]
2. Goyal, R., Hirano I. (1996). The Enteric Nervous System, Review Article. NEJM, 334, 1106-1115 [ Links ]
3. Hofer, D., Asan, E., Drenckhahn, D. (1999). Chemosensory Perception in the Gut. News Physiol Sci, 14, 18-23 [ Links ]
4. Raybould, H. (1998). Does Your Gut Taste? Sensory Transduction in the Gastrointestinal Tract. News Physiol Sci, 13, 275-280 [ Links ]
5. Chen, D., Aihara, T., Zhao, C., Hakason, R., Okabe, S. (2006). Differentiation of the Gastric Mucosa. I Role of histamine in control of function and integrity of oxyntic mucosa: understanding gastric physiology through disruption of targeted genes. AJPGastrointest Liver Physol, 291, 539-544 [ Links ]
6. Kasper, D., Fauci, A., Longo, D., Braunwald, E., Hauser, S., Jameson, J. (2005). Harrison, Principios de Medicina Interna. (16º ed.). México: McGrawHill. [ Links ]
7. Physiology of gastric acid secretion. (2009, May). www.uptodate.com Soll, A. [ Links ]
8. Feldman, M., Friedman, L., Lawrence, B. (2006). Sleisenger and Fordtran’s Gastrointestinal Liver Disease. ( 8º ed.). USA: Saunders [ Links ]
9. Yakabi, K., Kawashima, J., Kato, S. (2008). Ghrelin and gastric acid secretion. World J Gastroenterol, 14, 6334-6338 [ Links ]
10. Peeters, T. (2003). Central and peripheral mechanisms by which ghrelin regulates gut motility. Journal of physiology and pharmacology, 4, 95-103 [ Links ]
11. Tack, J., Depoortere, I., Bisschops, R., Delporte, C., Coulie, B., Meulemans, A., Janssens, J. (2006). Influence of ghrelin on interdigestive gastrointestinal motility in humans. Gut, 55, 327-333 [ Links ]
12. Phillison, M., Johansson, M., Henriksnas, J., Petersson, J., Gendler, S., Sandler, S., Persson, E., Hansson, G., Holm, L. (2008). The gastric mucus layers: constituents and regulation of accumulation. Am J Physiol Gastrointest Liver Physiol, 295, 806-812 [ Links ]
13. Liddle, R. (2009, May). Physiology of somatostatin and its analogues www.uptodate.com [ Links ]
14. Jain, R., Samuelson, L. (2006). Differentiation of the Gastric Mucosa. II Role of gastrin in gastric epithelial cell proliferation and maturation. Am J Physiol Gastrointest Liver Physiol, 291, 762-765 [ Links ]
15. Liddle, R. (2009, May). Physiology of gastrin www.uptodate.com [ Links ]
16. Guyton, A., Hall, J. (2006). Textbook of Medical Phisiology. (11º ed.). USA: Elsevier Saunders [ Links ]
17. Busque, S., Kerstetter, J., Geibel, J., Insogna, K. (2005). L-Type amino acids stimulate gastric acid secretion by activation of the calciumsensing receptor in parietal cells. Am J Physiol Gastrointes Liver Physiol, 289, 664-669 [ Links ]
18. Hurwitz ,A., Brady, D., Schaal, E. (1997). Gastric Acidity in Older Adults. JAMA, 278, 659-662 [ Links ]
19. Silverthorn, D. (2008). Fisiología Humana, Un enfoque integrado. (4º ed.). Buenos Aires, Argentina: Editorial Médica Panamericana [ Links ]
20. Hansen, M. (2003). Neurohumoral Control of Gastrointestinal Motility. Physiol Res, 52, 1-30 [ Links ]
21. Tack, J., Sarnelli, G. (2002). Serotonergic modulation of visceral sensation: upper gastrointestinal tract. Gut, 51, 77-80 [ Links ]
22. Kamerling, I., Van Haarst, A., Burggraaf, J., Schoemaker, R., Kam, M., Heinzerling, H., Cohen, A., Masclee, A. (2003). Effects of a nonpeptide motilin receptor antagonist on proximal gastric motor function. Br J Clin Pharmacol, 57, 393-401 [ Links ]
23. Beglinger, C., Degen, L. (2002). Role of thyrotrophin releasing hormone and corticotrophin releasing factor in stress related alterations of gastrointestinal motor function. Gut, 51, 45-49 [ Links ]
24. Kim, H. (2009). 5-Hydroxytryptamine4 receptor agonists and colonic motility. J Smooth Muscle Res, 45, 25-29 [ Links ]
25. Kunzelmann, K., McMorran, B. (2004). First Encounter: How Pathogens Compromise Epithelial Transport. Int Union Physiol Sci, 19, 240-244 [ Links ]

