











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
Los lepidópteros son uno de los grupos de organismos más grandes que existen, conocidos comúnmente como mariposas y polillas. Son importantes objeto de estudio debido a que muchas especies son plagas de importancia económica en la agricultura. Se estima que este orden posee aproximadamente 157 424 especies incluyendo 22 especies fósiles clasificadas en 45 superfamilias, 134 familias y 15 562 géneros, siendo el segundo grupo más diverso de animales que existen, después del Coleóptera (1) por lo que muchas de éstas todavía no han sido estudiadas. En Costa Rica habitan más de 12 000 especies diferentes de lepidópteros (2).
Los lepidópteros presentan una metamorfosis completa, con alas membranosas cubiertas de escamas (2). La mayoría de los lepidópteros en su estado larval se alimentan de las plantas, mientras que en su estado adulto se alimentan del néctar de las flores, siendo muy importantes en los ecosistemas debido a su función como herbívoros, al ser uno de los grupos de plagas más dañinos en la agricultura; así también como polinizadores, debido a sus hábitos de alimentación en su estado adulto y como presa de otros insectos (3).
Noctuidae contiene alrededor de 42 400 especies y es la superfamilia más grande dentro de lepidóptera (4). Se caracteriza porque sus ejemplares poseen hábitos nocturnos y son atraídos por la luz, la mayoría son polífagos y se alimentan de hojas, no obstante, algunas otras especies atacan órganos como los frutos o los tallos (5). (6) determinó que más de 6000 especies de lepidóptera son de importancia económica como plagas de cultivos agrícolas, muchas de las cuales se ubican dentro de la superfamilia Noctuidae.
La familia Erebidae (anteriormente denominada Arctiidae) está dentro de la superfamilia Noctuidae, el cual incluye pocas especies plagas de cultivos, los cuales producen daños localizados y con una periodicidad baja. Las larvas de Erebidae son muy peludas, cilíndricas y gruesas; mientras que los adultos tienen un tamaño mediano a grande, con colores mayoritariamente negros o blancos con franjas de color o manchas, los cuales presentan un cuerpo de delgado a robusto y alas enteras donde las posteriores son notablemente más pequeñas que las anteriores (2).
Estigmene albida es un lepidóptero perteneciente a la familia Erebidae, el cual se encuentra distribuido en América. Posee un hábitat que se ubica principalmente en Estados Unidos, donde se encuentra en lugares como Idaho, Montana, Arizona, Nuevo México, el oeste de Dakota del Sur y el sureste de California, así como gran parte de México (7). El Servicio de Información sobre Biodiversidad Global (GBIF, por sus siglas en inglés), localiza a este insecto principalmente en Estados Unidos, sin embargo, menciona a Costa Rica como el segundo país con más reportes de aparición (8). Sin embargo, la información disponible para E. albida es escasa a nivel mundial.
Es por esta razón, que este estudio realizó una comparación de los resultados obtenidos de E. albida con la especie Estigmene acrea, uno de los ejemplares más importantes de esta familia y que pertenece al mismo género que el lepidóptero en estudio. Se han realizado varios reportes de E. acrea como insecto plaga de importancia económica atacando más de 60 especies de plantas diferentes (9), siendo los arvenses de hoja ancha los hospederos más comunes (10) como Amaranthus sp. y Malva sp. (11), sin embargo, esta plaga puede expandir su rango de acción y atacar a cultivos como el maíz, el algodón, la soya, la remolacha azucarera (10), el frijol, el tabaco y diferentes hortalizas (11).
El higo (Ficus carica) es un cultivo mediterráneo de importancia económica debido a su fruto y calidad nutricional. En Costa Rica el higo se cultiva principalmente en la zona Norte de Cartago (12), en donde hay reportes del gusano cogollero del maíz (Spodoptera frugiperda) atacando este cultivo (13), sin embargo, la mayoría de plagas de importancia en este cultivo no están presentes en Costa Rica, e incluso, algunas de ellas, a nivel fitosanitario son catalogadas como plagas cuarentenarias para este cultivo, como la cochinilla rosada (Maconellicoccus hirsutus) y la cochinilla de la higuera (Ceroplastes rusci) (14), lo que demuestra que existe poca información referente a las plagas que pueden atacar a este cultivo en el trópico y en Costa Rica.
Este artículo brinda uno de los primeros reportes de Estigmene albida como insecto atacando cultivos agrícolas de interés comercial y el primer reporte de este insecto atacando el cultivo del higo (Ficus carica) en Costa Rica. Es por esta razón, que el objetivo de este estudio fue determinar el ciclo de vida y comparar la fertilidad de Estigmene albida bajo condiciones controladas de laboratorio aisladas en el cultivo del higo (Ficus carica), así como la identificación de un parasitoide asociado a éste, como potencial agente biocontrolador.
Materiales y Métodos
Colecta del material
Hojas con larvas de insectos fueron colectadas de una plantación de higo (Ficus carica) ubicada en el Campo Frutícola del Centro de Investigación en Biotecnología (CIB) ubicado en Cartago, en las coordenadas 09° 51’ 09.1’’ (±5) latitud norte y 083° 54’ 25.4’’ (±5) longitud oeste el cual se encuentra a 1405 msnm. Las hojas fueron transportadas al Laboratorio de Biocontrol del CIB perteneciente al Instituto Tecnológico de Costa Rica. Cada larva presente en las hojas fue individualizada en contenedores de plástico con un volumen de 60 ml bajo condiciones ambientales controladas por medio de una cámara climática, con una temperatura de 24 ± 0,1 °C, una humedad relativa del 70 ± 0,5 % y un fotoperiodo de 16 horas luz y 8 horas oscuridad.
Descripción del ciclo de vida e identificación del lepidóptero
Cada larva individualizada fue alimentada mediante un segmento aproximado de 1 cm2 con la dieta artificial BIO-MIX H-89 modificada (15) y bajo las condiciones ambientales controladas descritas anteriormente. Cada dos días el alimento fue reemplazado y los envases con las larvas fueron cambiados por envases nuevos con el fin de limpiar los excrementos generados, hasta completar su desarrollo larval.
Posteriormente, cuando las larvas se transformaron en pupas, éstas fueron trasladadas a envases nuevos bajo las mismas condiciones ambientales controladas, en ausencia de alimento hasta que el adulto emergiera. Diez individuos adultos fueron colocados en bolsas de papel de 4000 cm3 con el fin de favorecer la copulación de los mismos. A cada bolsa se le colocó una mota de algodón de aproximadamente 40 ml de volumen impregnado con una solución de agua y miel en una proporción 1:1, como reemplazo del néctar de las flores. El algodón fue remplazado cada 4 días de las bolsas de papel.
A partir de las 48 h, las masas de huevos obtenidas en las bolsas de pared fueron recuperadas e individualizadas en recipientes de plástico de 60 ml, repitiendo esta actividad hasta la muerte de los adultos. Las masas de huevos se mantuvieron en los recipientes hasta su eclosión, momento en el cual se volvieron a individualizar, repitiendo el proceso de crianza de las larvas nuevamente.
Se determinó la duración de cada etapa de desarrollo para cada individuo, desde su fase de huevo hasta la adultez con el fin de establecer su ciclo de vida, detallando en los diferentes estadíos larvales. Por último, cuando el ciclo de vida estaba establecido, los insectos adultos fueron identificados por el taxónomo especialista en lepidópteros Ing. José Joaquín Montero Ramírez. Todos los resultados obtenidos fueron comparados con datos reportados para E. acrea.
Determinación de la fertilidad y eclosión
Luego de completar el ciclo de vida, un adulto macho y una hembra fueron colocados en una bolsa de papel (4000 cm3) bajo las mismas condiciones ambientales controladas, repitiendo el procedimiento hasta obtener 44 parejas individualizadas. Se registraron las siguientes variables para cada pareja:
1.N inicial
2.Porcentaje de hembras que ovipositan
3.Promedio de días para primera oviposición/hembra
4.Promedio de huevos/hembra
5.Promedio de larvas/hembra
6.Porcentaje de Hembras con Posturas Viables (HPV):
•Promedio de días para primera oviposición/HPV
•Promedio de huevos/HPV
•Promedio de larvas/HPV
•Porcentaje promedio de viabilidad de huevos
•Promedio de días para eclosión/HPV
Estas mismas variables se evaluaron en seis hembras individualizadas para estudiar la incidencia de partenogénesis en la especie. Además, se compararon algunas de las variables anteriores según tipo de reproducción (sexual y partenogénesis) mediante pruebas inferenciales, según los datos cumplieran con los supuestos estadísticos de normalidad y homocedasticidad. La normalidad de los datos se evaluó mediante las pruebas de Anderson-Darling, Ryan-Joiner o Kolmogorov-Smirnov; mientras que la homocedasticidad se evaluó con la prueba de Levene. Todos los análisis se realizaron asumiendo un valor α = 0,050 mediante el programa Minitab® (16).
Además, se aplicó la correlación lineal de Spearman para evaluar la relación de dependencia entre la cantidad de huevos por pareja, los días para la primera oviposición y la cantidad de larvas eclosionadas para las hembras en apareamiento que produjeron huevos. Posteriormente, se analizó la dependencia entre la cantidad de larvas y el tiempo de eclosión para los casos en que se registró eclosión producto de HPV. Todos los análisis se realizaron asumiendo un valor de α = 0,050 mediante el programa Minitab® (16). Todos los resultados obtenidos fueron comparados con datos reportados para E. acrea.
Aislamiento e identificación de parasitoide
De la colecta de insectos que se realizó desde el Campo Frutícola, tres larvas presentaron síntomas de parasitismo por otro insecto, debido a su baja movilidad, falta de alimentación e hinchazón. Las larvas fueron manipuladas de la misma forma que las demás, sin embargo, fueron colocadas en envases plásticos de 500 ml y monitoreadas diariamente. Al día siguiente de la muerte de las larvas, se observó la generación de un capullo alrededor de la misma el cual contenía de 3 a 4 pupas, esto ocurrió bajo las mismas condiciones ambientales controladas. Al emerger los adultos, estos fueron fueron llevados para su identificación por el taxónomo especialista en dípteros Dr. Manuel Zumbado Arrieta.
Resultados y discusión
Descripción del ciclo de vida e identificación del lepidóptero
El lepidóptero fue identificado como Estigmene albida, una polilla de la superfamilia Noctuidae, familia Erebidae. El ciclo de vida de E. albida se estableció en 57 días totales bajo condiciones controladas de laboratorio (Figura 1). Para E. acrea se ha establecido que el ciclo de vida dura, en promedio, entre 35 a 42 días en condiciones ambientales (17), por lo que E. albida presenta un ciclo de vida 15 días mayor en condiciones controladas de laboratorio que el reportado para E. acrea, sin embargo, el mismo autor también reporta que en estado larval puede llegar a durar hasta 45 días, razón por la cual, se estima que el rango del ciclo de vida puede ser ampliado, dependiendo de las condiciones ambientales y del régimen de alimentación en el que se encuentren los insectos.
E. albida presentó huevos esféricos ovipositados en grupos que, en promedio, eran de 376 huevos por hembra, los cuales recién puestos eran de color amarillo, pero luego cambiaban a un color blanco-grisáceo, los cuales eclosionaban luego de 5 días. E. acrea presenta huevos esféricos que miden aproximadamente 0,6 mm, los cuales inicialmente son amarillos, pero luego cambian a un color grisáceo. Las hembras producen aproximadamente entre 400 a 1000 huevos, los cuales eclosionan después de 4 a 5 días (17). En general, el tiempo de eclosión de los huevos en ambas especies es similar, sin embargo, bajo condiciones de laboratorio, el promedio de huevos puesto por cada hembra en E. albida es inferior al reportado para E. acrea, el cual puede deberse a las condiciones limitadas de espacio en las que se encuentran los adultos en las bolsas de papel o a diferencias propias de cada especie.
E. albida presentó 5 estadíos larvales (figura 2). El primer estadío se caracterizó por larvas de color amarillo pálido, con pelos corporales finos de color negro. Al iniciar el segundo estadío larval, las larvas aumentaron la cantidad de pelos corporales y estos variaban de color marrón, café y negro, además presentaron una franja longitudinal de color amarillo o blanco. El tercer instar se caracterizó por un oscurecimiento de la larva y de los pelos corporales, evidenciándose todavía la franja longitudinal. Para el cuarto estadío larval, los pelos corporales son de color negro en la parte superior y marrón en la parte inferior, manteniendo este mismo patrón en el quinto estadío, solo que la larva es predominantemente negra y de mayor tamaño, durando 26 días en total en la fase larval. En general, en todos los estadíos, la larva se alimentaba vorazmente del alimento artificial, aumentando su consumo conforme incrementaba de tamaño.
Las larvas de E. acrea presentan de 5 a 7 instares. Al emerger, las larvas miden aproximadamente 2 mm, son de color amarrillo pálido hasta carmelita oscuro o marrón y son gregarias hasta alcanzar una longitud de 10 mm, además presentan largos pelos o setas. Al empezar el segundo instar, se observan franjas longitudinales generalmente de color marrón, castaño, amarillo y blanco; los pelos corporales se vuelven más negros, alcanzando una longitud de 15 mm, incrementan su movilidad y desarrollan mandíbulas fuertes. En el tercer instar, las larvas se oscurecen y alcanzan tamaños de hasta 30 mm. Esta apariencia se mantiene en el cuarto y quinto instar, alcanzando tamaños de hasta 45 y 55 mm respectivamente, aumentando su movilidad y voracidad. Se han dado reportes que el estado larval de E. acrea puede durar entre 20 hasta los 45 días (17) (18). En general, los estadíos larvales de E. acrea y E. albida son similares entre sí, tanto en su descripción como en la duración de la fase.
Al empezar la fase de pupa, E. albida formó un capullo de seda fino, los cuales se podían ubicar en la tapa o en la base de los envases plásticos, sin embargo, después de 8 días, el capullo se iba abriendo y quedaba únicamente la pupa desnuda, la cual es una pupa obtecta de color café oscuro, durando en total 16 días en esta fase. En cuanto a E. acrea, la pupa es de color marrón oscuro y se encuentran generalmente en el suelo en un capullo formado por pelos sedosos y los pelos corporales de la larva. Esta etapa dura entre 12 a 14 días (17), alcanzando una longitud de 32 mm (18), por lo que se evidencia un aumento del tiempo que E. albida requiere en la fase pupal, la cual puede deberse a las condiciones ambientales controladas a las que estaba o por las características propias de esta especie. Además, el capullo formado por E. albida, al encontrarse en condiciones controladas tiende a desaparecer conforme avance la fase pupal, característica que no pasa en el ambiente para E. acrea.
Por último, los adultos de E. albida presentaron una coloración mayoritariamente blanca, con pocas manchas irregulares en las alas o en los bordes de ésta, presentando algunas una ausencia total de las mismas. Los machos eran más pequeños que las hembras y sus alas traseras eran de color ligeramente crema, característica no observada en las hembras. Los segmentos abdominales son de un color amarillo con manchas negras dorsalmente. Los adultos produjeron masas de huevos después de, en promedio, 48 h desde que se colocaban juntos, duración similar a lo reportado para E. acrea.
Los adultos de E. acrea son polillas que miden entre 3,5 a 4,5 cm de envergadura. Son predominantemente blancas con alas con pequeñas e irregulares manchas negras. Las alas traseras son amarillas en los machos y blancas en las hembras, presentando un dimorfismo sexual. Los segmentos abdominales son mayoritariamente amarillos y llevan una serie de manchas negras dorsalmente en los pleuritos y en los esternitos. Un día después de la emergencia del adulto ocurre la copulación, y luego de esta, 24 h después ocurre la deposición de huevos. Las hembras pueden vivir de 4 a 5 días y pueden poner más de un grupo de huevos (17) (18).

Figura 1. Ciclo de vida de Estigmene albida bajo condiciones ambientales controladas.

Determinación de la fertilidad y eclosión
En el cuadro 1 se presenta el resumen de las variables en estudio relacionadas con la fertilidad y la eclosión en la especie E. albida.
Cuadro 1. Resumen de las variables asociadas con fertilidad y eclosión en E. albida.
| Variable | Tipo de reproducción | Valor p | |
| Sexual | Partenogénesis | ||
| 1. n inicial = hembras en evaluación | 44 | 6 | --- |
| 2. % de hembras que ovipositaron | 65,9 (29 de 44) | 83,3 (5 de 6) | --- |
| 3. Promedio de días para primera oviposición/hembra | 3,2 ± 2,1 2,0 (mediana) | 4,6 ± 3,8 (5 mediana) | p>0,0501 |
| 4. Promedio de huevos/hembra | 375,4 ± 129,4 | 324,2 ± 103,8 | p>0,0502 |
| 5. Promedio de larvas/hembra | 38,3 ± 109,0 0,0 (mediana) | 0,0 0,0 (mediana) | p>0,0503 |
| 6. % de Hembras con Posturas Viables (HPV) | 17,2 (5 de 29) | 0,0 (0 de 6) | --- |
| 6.1 Promedio de días para primera oviposición/HPV | 2,0 ± 0.0 | --- | --- |
| 6.2 Promedio de huevos/HPV | 377,4 ± 100,2 | --- | --- |
| 6.3 Promedio de larvas/HPV | 222,4 ± 179,0 | --- | --- |
| 6.4 % promedio de viabilidad de huevos | 59,0 | --- | --- |
| 6.5 Promedio de días para eclosión/HPV | 6,0 ± 2,2 | --- | --- |
1 Prueba t-student para 2 muestras independientes (medias) y Prueba Mann Whitney U (medianas).
2 Prueba t-student para 2 muestras independientes (medias).
3 Prueba t-student para una muestra (media) y Prueba de Wilcoxon de una muestra (mediana).
Las medianas se calcularon y analizaron para aquellas variables que no cumplieron algún supuesto estadístico.
A partir de los resultados del cuadro 1 se puede observar que las hembras partenogénicas ovipositaron en mayor proporción (83,5%) que las hembras del grupo de apareamiento (65,9%). Sin embargo, para el caso de las variables analizadas estadísticamente entre los dos tipos de reproducción: el promedio de días para la primera oviposición/hembra (variable 2), el promedio de huevos/hembra (variable 3) y el promedio de larvas/hembra (variable 4); no se encontraron diferencias significativas entre sus valores promedio ni entre las medianas, según la prueba correspondiente. Lo anterior muestra evidencia de que la cantidad de huevos y el tiempo promedio en que una hembra adulta de E. albida oviposita por primera vez, no dependen de la cópula previa en esta especie. Diferencias entre los dos tipos de reproducción analizadas en diversos invertebrados han señalado que la reproducción sexual predomina ante la escasez de recursos o cuando se requieren múltiples genotipos para la explotación más eficiente de los recursos; mientras que la reproducción sin fecundación se favorece cuando los recursos están más disponibles según las estaciones del año, generalmente cuando se requiere más cantidad de individuos de un solo genotipo para explotar recursos específicos (19).
Investigaciones realizadas en Estigmene acrea sobre los factores que influyen en la reproducción han determinado que el tipo de dieta tiene influencia directa sobre la cantidad de oviposición y el tiempo de postura, así como sobre la sobrevivencia de las hembras (20), las cuales ovipositan mayor cantidad de huevos de forma temprana cuando se alimentan de material vegetal en comparación con una dieta sin material fresco, y viven en promedio dos días más (9 días). En esa investigación encontraron que la cantidad de huevos promedio por hembra se encontraba entre 548 para la reproducción sexual y 429 para la partenogénica, siendo estadísticamente mayor cuando se alimenta de material vegetal fresco, así como un tiempo para la primera oviposición que varía entre 3,30 y 4,66 días siguiendo esa misma relación. En nuestro caso, el pie de cría se alimentó con dieta formulada y el promedio registrado de 376 (sexual) - 325 (partenogénica) huevos por hembra y 3,2 (sexual) - 4,6 (partenogénica) días para la primera oviposición, resultados que se asemejan al reportado para E. acrea, a pesar de que en este estudio se utilizó una dieta modificada y no material fresco.
Por otro lado, el apareamiento demostró ser el tipo de reproducción efectiva para producir huevos viables que eclosionen en larvas, representado por un 17,2 % de hembras que lograron descendencia; mientras que ninguna hembra partenogénica produjo larvas. Según el modelo propuesto por (19), la reproducción sexual predomina en invertebrados cuando responde a estrategias de supervivencia que requieren múltiples genotipos para la explotación de recursos. Dentro de las primeras evidencias sobre la fertilidad de especies relacionadas, (21) demostraron que el porcentaje de huevos viables en la especie E. acrea es variable entre las diferentes generaciones, siendo un 48,0 ± 14,0 % para la F1, donde los factores como la dieta, la transferencia de adultos a nuevos criaderos y los pequeños periodos sin alimento pueden afectar la producción de huevos viables.
En otra investigación, también se asoció el éxito en la reproducción con la alimentación de los machos, los cuales mejoran la fertilidad cuando se alimentan de material vegetal fresco por la influencia de alcaloides pirrolizidínicos de las hojas sobre el desarrollo de la coremata (22). La variación de la temperatura también tiene una influencia importante en la fertilidad de E. acrea, según la investigación realizada por (23), temperaturas de 25°C alcanzan un 66% de eclosión, sin embargo, al incrementarse la temperatura a 35-40°C y las horas de exposición, la variación de dicho porcentaje es considerable (74-0% de eclosión). También, la variación de la temperatura combinada con diferentes regímenes de humedad relativa (10-60%) provocan alteraciones en el porcentaje de eclosión de los huevos en E. acrea, ya que conforme las condiciones se vuelven estresantes se disminuye la eclosión exitosa (24). Además, (25) describieron también la sensibilidad de E. acrea al incremento de la temperatura y cómo este factor afecta la cantidad y duración de los estadíos. Sin embargo, nuestro experimento controló estos factores para evitar presión adicional sobre la eclosión de las larvas y encontramos que el porcentaje de eclosión de huevos viables es menor en E. albida comparado con lo reportado para E. acrea.
En cuanto al promedio de larvas por hembra (variable 5) registrado solo para la reproducción sexual, su valor alcanzó 38,3 individuos pero presentó una variación importante de ± 109,0 individuos, lo cual provocó que no existiera diferencia significativa (p > 0,050) al comparar esta serie de datos con 0,0 de eclosión de larvas provenientes de hembras partenogénicas; sin embargo, esta condición de 0,0 % de huevos viables en las hembras partenogénicas se debe verificar con un estudio posterior utilizando mayor cantidad de individuos para definir la incidencia del fenómeno de partenogénesis en E. albida, ya que este dato no está actualmente disponible en investigaciones previas.
Las variables analizadas sobre las HPV sugieren un tiempo promedio de eclosión de 6,0 ± 2,2 días posterior a la postura. El 59 % de sus huevos fueron viables y produjeron 223 larvas en promedio por hembra en su primer ciclo de reproducción. No obstante, la desviación estándar calculada fue también alta para estas variables asociadas con la fertilidad.
Por otro lado, el análisis de Anderson Darling encontró que algunas series de datos para la matriz de correlaciones no presentaban distribución normal (p < 0,050) y se seleccionó entonces la prueba de Spearman para determinar la relación lineal entre las variables en estudio.
De las 44 parejas de insectos que se monitorearon se obtuvo postura de huevos en solo 29 de los casos (representa el 65,9 %). A partir del análisis de estas 29 parejas se encontró que existe correlación lineal significativa (p < 0,050) solo para las variables Larvas y Primera oviposición, donde eclosionan menor número de larvas conforme la oviposición de los huevos ocurra luego de los primeros dos días de la reunión de la pareja. El coeficiente rho obtenido -0,411 indica la magnitud y la dirección de esta relación (figura 3a). Para las variables de Huevos y Primera oviposición los coeficientes obtenidos fueron positivos, sin embargo, no representaron una relación lineal significativa entre ellas (figura 3b y 3c). Se obtuvo también relación no significativa para Huevos y Larvas, en contraste con otras investigaciones en lepidópteros donde generalmente estas variables se han encontrado relacionadas (13).
De las 29 parejas que ovipositaron, solo en 5 de los casos se obtuvo eclosión de larvas (representa el 17,2 %). Estos datos se analizaron también para la correlación lineal de Spearman encontrando un coeficiente negativo, pero no significativo (figura 3d).
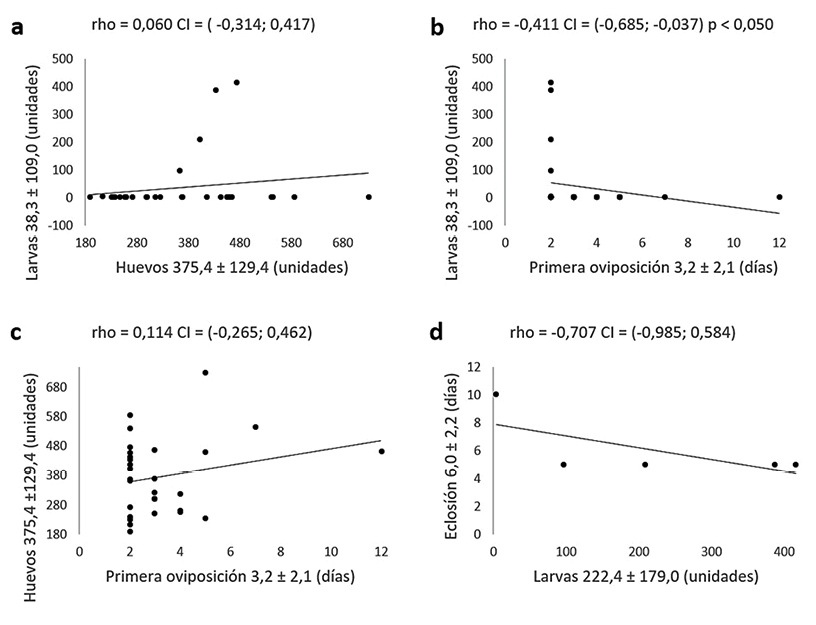
Figura 3 Matriz de correlación lineal de Spearman, Intervalo del 95 % de confianza para el coeficiente rho y significancia de la prueba (α = 0,050). Hembras en reproducción sexual. a, b, c. Relación para parejas que presentaron oviposición (n=29). d. Relación para larvas eclosionadas (n=5).
Figura 3. Matriz de correlación lineal de Spearman, Intervalo del 95 % de confianza para el coeficiente rho y significancia de la prueba (α = 0,050). Hembras en reproducción sexual. a, b, c. Relación para parejas que presentaron oviposición (n=29). d. Relación para larvas eclosionadas (n=5).
Aislamiento e identificación del parasitoide.
El control biológico viene a ser una alternativa para la reducción en el uso de productos químicos en los cultivos, la reducción de costos y el desarrollo de una agricultura más sostenible. Dentro del control biológico el uso de depredadores y parasitoides juega un papel importante, ya que estos mantienen su acción reguladora en el tiempo, contrario a los productos químicos, evidenciando el gran potencial que tienen para controlar las principales plagas, enfermedades y arvenses que existen (26). Los parasitoides son los organismos más utilizados en el control biológico (26), lo que evidencia que la búsqueda e identificación de nuevos microorganismos biocontroladores autóctonos o de ya existentes con nuevos hospederos es beneficioso y puede ser usado, en el futuro, como una fuente de control de una potencial plaga en los cultivos.
De las larvas colectadas inicialmente, tres presentaron indicios de parasitismo. Al dar el seguimiento de las mismas, en cada larva salieron de 3 a 4 pupas coarctatas de las cuales emergió un díptera (figura 4a). El adulto fue identificado como Carcelia sp. una mosca de la familia Tachinidae (figura 4b) (27). Esta familia es de las más grandes de este orden, con aproximadamente 8 200 especies (27), los cuales son enemigos naturales importantes de plagas de insectos, especialmente de larvas de lepidópteros, por lo que son usados frecuentemente en el control biológico (28). Se ha reportado que el taquínido Carcelia reclinata es un parasitoide polífago de E. acrea y otros lepidópteros defoliadores (28) (29). Asimismo, (30) reportaron que el taquínido Gymnocarcelia ricinorum presenta un 35% de parasitismo natural contra E. acrea en el cultivo de la fresa.
Por otro lado, (31) reporta que todos los taquínidos son endoparasíticos, mayoritariamente de larvas de lepidópteros, los cuales poseen tres estadíos larvales, atacan larvas o pupas, pueden ser solitarios o gregarios, dependiendo de la especie y que la fase pupal puede ocurrir tanto dentro como fuera del hospedador. Además, clasifica a los taquínidos, como koinobiontes conformadores, ya que, al parasitar al hospedero, este no muere, y más bien se beneficia que el hospedero se mantenga con vida, mostrando una integración fisiológica hospedante-parasitoide el cual requiere de las hormonas del hospedero para sincronizar su desarrollo. En este caso, Carcelia sp se comportó como un endoparasitoide, gregario, koinobionte, el cual forma su pupa dentro de un capullo afuera de la larva de E. albida.

Figura 4. Carcelia sp. como parasitoide de E. albida. a- Pupas obtenidas de larvas parasitadas. b- Adulto
Figura 4. Carcelia sp. como parasitoide de E. albida. a- Pupas obtenidas de larvas parasitadas. b- Adulto
Conclusiones
Estigmene albida es un lepidóptero que pertenece a la familia Erebidae, superfamilia Noctuidae, el cual presenta muy poca información reportada en literatura. Se determinó que su ciclo de vida bajo condiciones ambientales controladas en laboratorio y bajo una dieta artificial fue de 57 días, reportando cinco estadíos larvales muy similares a los que se reportan para E. acrea, pero presentando una fase pupal y de adultez con una duración mayor. Además, se logró determinar que, aunque las hembras partenogénicas ovipositaron en mayor proporción que las hembras que copularon, ninguno de los huevos que produjeron fueron viables, caso contrario a las hembras sexuales, de las cuales el 17.2 % produjeron descendencia ovipositando en promedio 376 huevos por hembra, de los cuales el 59 % eclosionó en larvas viables. Además, se logró identificar a Carcelia sp. como un taquínido parasitoide para E. albida, el cual tiene potencial como posible agente biocontrolador para este lepidóptero.
