











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
In Costa Rica, the tropical dry forest (TDF) life zone (1) is mainly found in the Tempisque River’s lower basin, in the Guanacaste province (2). Climatically, this zone is characterized by high temperatures and the marked rainfall seasonality, represented by 4 to 6 months of drought (2) (3) with an average annual rainfall of less than 2000 mm (2). Taking these conditions into account, the TDF presents greater bioclimatic variation throughout the year than the country’s wet and rainy forests (4)
Such environmental variation makes it necessary to the plants and animals that inhabit these areas to develop special morphological, physiological and behavioral adaptations. In this sense, forests that develop in this life zone are mainly deciduous during the driest months, synchronizing flower and fruit production with water availability (3). In the same way, animals must adapt to the forest conditions throughout the year, especially with water scarcity and food resource fluctuation (5)
Regarding animals, bats stand out as an interesting case of study. Bats are a diverse and abundant group of mammals, with more than 1300 species in the world (6). As a group, bats have great ability adapting into different environments (7). This is reflected by the variety of trophic guilds being developed, such as frugivorous, nectarivorous, carnivorous, insectivorous, fishing and even hematophagous bats. Bat’s trophic diversity is not only interesting in ecological terms. The variety of trophic guilds turns bats into important ecosystem services providers. Their role as seed dispersers and plant pollinators as well as pest insect controllers, has been highlighted (8)
Taking into account the TDF environmental variability throughout the year and bat adaptability capacity, our objective was to analyze the assemblage and guild composition of bats related to the rainfall pattern of an agricultural TDF landscape of Costa Rica.
Materials and methods
The study took place in the Laguna Mata Redonda Wildlife Refuge (LMRWR), located in the Tempisque River’s lower basin in the Guanacaste province, Costa Rica, between 10.77° 19.015’ latitude and 85° 24.971’ longitude. The area is 30 meters above sea level. The LMRWR is a 400 ha protected area, mainly composed by a wetland, secondary forest fragments and a wooded savanna. Its dry season is characterized by 5 to 6 months (between December and May) of drought. During the year its average temperature is 33.75 °C and the average precipitation is 143.36 ± 130.80 mm per month (9).
Bat sampling was carried out between March and August of 2012, accomplishing 17 nights, distributed as: six nights during dry season (March), five nights during the dry-to-rainy-transition season (May) and six nights during rainy season (July and August). Animal capture was carried out using four 12m x 2.5m mist nets with 36mm of mesh. Nets were active between 18:00 h to 22:00 h, due to greatest activity reported for phyllostomids (10), for an overall of 340 net/hours. Nets were moved every night, distributed among the main natural elements of the landscape: small secondary forest patches, wooded savanna and wetland borders. Given site accessibility, only the eastern margin of the wetland was sampled.
Each animal captured was identified up to species level according to Timm et al. (11)and LaVal et al. (12) and classified by guild as in LaVal et al. (12) and Gonzalez-Salazar et al. (13). All captured specimens were released as soon as possible and no animals were collected. All the research permits requested by the National Conservation Areas System (SINAC) of Costa Rica were accomplished.
Assemblage composition between seasons was analyzed using a similarity percentage analysis (SIMPER) (PAST software v.1.90). SIMPER breaks down the contribution of each species individuals to the similarity or dissimilarity observed between the samples, allowing us to identify the most important species in the observed dissimilarity pattern (14). Precipitation data used was produced with the online data system GIOVANNI (https://giovanni.gsfc.nasa.gov/giovanni/), developed and maintained by the Goddard Data and Information Service of the NASA Earth Sciences Laboratory (15).
Results
A total of 163 individuals of 11 species were captured throughout the sampled period. During dry season, Glossophaga soricina was the most frequently captured species, while in the rainy season was Artibeus jamaicensis (table 1). During the rainy season, more species richness (eight species) was found, as opposed to the dry season (five species). According to SIMPER analysis, the dissimilarity of the assemblage between dry and rainy season is 70.54%; being A. jamaicensis and G. soricina the species that contribute the most with the dissimilarity between seasons (table 1).
Table 1 Guilds and captures of bat species in the LMRWR. (+ = mainly; NR= not registered; n=individuals, %: relative percentage of total captures).
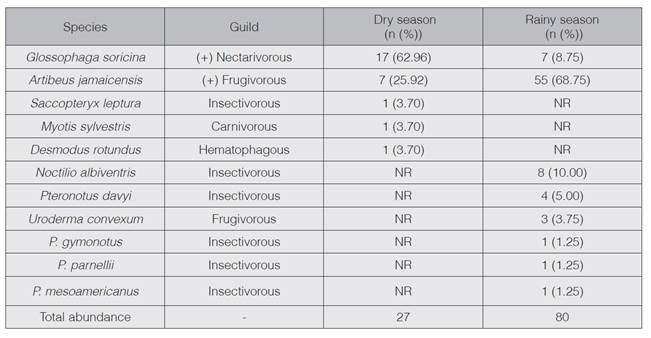
When analyzing the catches according to guilds, distinctive patterns are observed throughout the sampled period (figure 1). During the driest months reported in 2012 (from January to March) and during the dry-rainy transition period (April to May), the majority of catches correspond mainly to nectarivorous species. However, as precipitation increases the representation of this guild decreases. On the other hand, frugivorous captures present an inverse pattern, since during the driest months its presence is lower compared to the wetter months. A similar trend to that presented by frugivorous is observed within the insectivore guild, where catches during dry season are lower compared to the months of greater precipitation. In the case of the hematophagous (represented only by Desmodus rotundus) catches were accomplished only during 2 months (March and May). Given the minimal capture of carnivores (only one individual of Myotis sylvestris) a temporary trend for this guild cannot be described.
Discussion
The bat assemblage in the LMRWR varies distinctively throughout the year both in richness and abundance of species as in guild composition. In terms of richness and abundance, the assemblage is considerably dissimilar between seasons, however a similar pattern prevails. During both seasons, one species dominates the assemblage (G. soricina during dry season and A. jamaicensis during rainy season) with several other species being less abundant (even rare). This pattern may indicate that food availability, as we further emphasize when analyzing the assemblage in terms of guilds, maybe is one of the most important factors in the conformation of the assemblage between seasons.
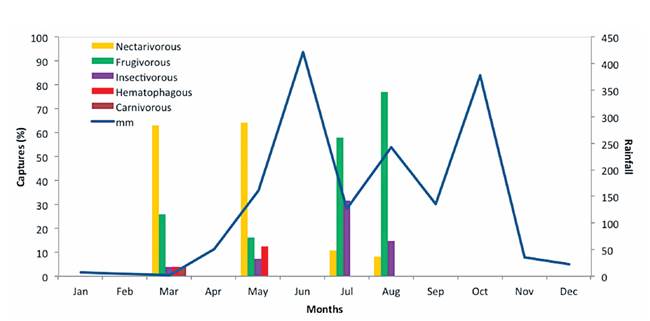
Figure 1 Rain pattern (mm) in the Laguna Mata Redonda Wildlife Refuge during 2012 and its relation with bat guild captures (%) in four months.
Speaking in terms of guilds, frugivorous and nectarivorous presences seem to be associated with on-site plant phenology which is closely related with the precipitation pattern. In the Guanacaste TDF there are general distinctive flowering and fruiting periods (16): the first one during the early dry season (when the greater nectarivorous presence happens) and a second one during the late rainy season; while fleshy fruit production and maturation occurs during the mid-late rainy season (when greater frugivorous presence arises).
In the case of the insectivore guild, its presence seems to be associated with an increase in the wetland size due to higher precipitation. During the rainiest months, the wetland reaches its highest water level, turning into an insect reproduction site, offering greater food resources for insectivorous bats. It has been detailed how insectivorous species tend to respond with greater intensity to insect concentration than to physical habitat conditions, as demonstrated for the insectivorous species Pteronotus parnellii (mesoamericanus) (17).
The activity of hematophagous bats seems to be associated with at-site livestock enclosure dynamics. During late dry and early rainy seasons, cattle are usually grouped on barns or paddocks where water and food is farmer-supplied due to the scarcity of natural pastures. Once rains stabilize, allowing pastures to grow and the wetland to fill up, cattle is allowed to graze freely. This dynamic seems to be exploited by D. rotundus, as noted by the fact that its activity is concentrated in forest patches near enclosures and paddocks; while during the free gracing period D. rotundus must pursuit cattle farther away, thus diminishing its presence in the area.
Our observations for frugivorous, nectarivorous and insectivorous species agree with Stoner & Timm (5). In Palo Verde National Park, over three years of sampling, these authors observed how the capture of different species of bats varies between seasons, associating it with food resources availability. Palo Verde is an area located less than 10 km away from the LMRWR, belonging to the same life zone, so we expect that the climatic and phenological conditions to be similar with those on our study area. Also our observations support the fact that TDF mammals can be classified as generalists (which change their diet over time in order to use available resources) or seasonal migrants (which occasionally use TDF and move to other forest types depending on the availability of resources) (5). Our results provide evidence to consider that the LMRWR bat assemblage is conformed of generalist seasonal migrants
Conclusion
Despite our low sampling effort and the fact that it was limited to four months, our results highlight the variance in a “simple” bat assemblage during the year, probably due to local movement of species related to food availability. This condition highlights the importance of natural elements remaining in agricultural landscapes (such as forest fragments and seasonal wetlands) as sources of temporary resources. Given general low protection and connectivity between Central- American TDF areas (18), biodiversity maintenance will depend not only on protected wild areas, but on conservation capacity at the landscape level of different natural elements that are still preserved in human-impacted areas.
Acknowledgments
To the Área de Conservación Arenal Tempisque (ACT) and Juan Bravo (CEMEDE-UNA) for promoting the research. To the LMRWR park rangers Jacinto and Martin for their field assistance. To Gustavo Ruiz, Nataly Obando, Irene Mata and Irene Guzmán for their field assistance. To Lisa Alfaro for translation review.

