











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
En Costa Rica, el chile dulce (Capsicum annuum) es la sexta hortaliza más cultivada, con un área de cultivo aproximada de 500 ha, una producción de 8533 t, un consumo per cápita anual de 3,5 kg (SEPSA 2018) y un rendimiento de 8,2 a 43 t.ha-1 en cultivo a campo abierto (MAG 2007) y hasta 128 t.ha-1 en invernadero (Soto-Bravo et al. 2020). En el 2014 se exportó aproximadamente 1,3 millones de kg a Estados Unidos (Mora et al. 2018).
El tomate (Solanum lycopersicum L.) es el cultivo que más se siembra en Costa Rica, con el 90% a campo abierto y un 10% en ambiente protegido. En el 2015, su cultivo generó un ingreso de $37 millones, con un consumo per cápita de 17,3 kg (López 2017). En el 2018, el área cultivada se estimó en 1100 ha con una producción total de 57 238 t (INEC 2020).
Para la producción de ambos cultivos, se requiere de emplear una gran cantidad de recurso hídrico. Pero debido al agotamiento y contaminación de este recurso indispensable para la producción, junto al cambio climático y la creciente demanda de alimentos debido al acelerado crecimiento demográfico, se ha generado todo un reto que enfrentará la agricultura en el presente siglo, asegurar su sostenibilidad económica, social y ambiental. Paralelamente, se proyecta un incremento sustancial en la demanda de agua dulce para riego de cultivos en general, la cual representa el 1,5% del total del recurso hídrico disponible en el planeta, y donde el sector agrícola es el principal demandante con un 69% (FAO 2017).
De acuerdo con lo planteado anteriormente, es evidente la necesidad de mejorar la eficiencia de uso del agua (EUA) que representa la relación que existe entre la producción de biomasa (kg) y el consumo de agua (m3) del cultivo. Para mejorar la EUA, una alternativa es la producción hidropónica en invernadero (Fernández y Camacho 2005, Fernández 2006). En dicho sistema de cultivo es posible mejorar la EUA al regular la evapotranspiración del cultivo (ETc) a través de la reducción de sus componentes de evaporación y transpiración foliar. En sistemas hidropónicos, habitualmente el sustrato está cubierto de plástico, por lo que la evaporación de agua es despreciable, disminuyendo así la ETc, ya que se limita únicamente al componente de transpiración.
Para disminuir la transpiración del cultivo se utilizan riegos deficitarios (RD), un método de irrigación en el cual se suministra un volumen de agua inferior al requerimiento (Chai et al. 2016) sin causar estrés fisiológico que pueda afectar el rendimiento del cultivo.
Para utilizar la técnica de RD con el fin de incrementar la EUA, es necesario determinar previamente los valores óptimos o umbrales de agotamiento de humedad (UAH) en el sustrato (Nardella et al. 2012, Mattar et al. 2020). Los UAH se generan a partir de ensayos con tratamientos de RD decrecientes, con el objetivo de disminuir la ETc manteniendo o incrementando el rendimiento del cultivo, lo que mejora así la EUA.
El manejo de la irrigación con UAH permite un adecuado balance entre el área foliar (fuente) y los frutos (sumidero), que en términos prácticos se conoce balance vegetativo-generativo en cultivos de fruto. Para esto es importante caracterizar los patrones morfofisiológicos y productivos del cultivo, para ser utilizados como indicadores del balance fuente/sumidero que permite obtener altos rendimientos. Dichos patrones dependen de la genética, del clima y del manejo del riego y la nutrición (Qian et al. 2012, Li et al. 2017), ya que influyen directamente en la distribución de asimilados y consecuentemente en el balance fuente/sumidero (Iqbal et al. 2015).
Diferentes estudios en distintos países han demostrado que el RD contribuye a mejorar la EUA en los cultivos. En Almería, España, en estudios realizados por Fernández y Camacho (2005) en cultivos bajo invernadero, obtuvieron una EUA de 37 kg.m-3, mientras que, en México, Flores et al. (2007) reportaron valores de EUA de 35 kg.m-3. Recientemente, en España, Valcárcel et al. (2019) demostraron que al reducir un 50% la ETc durante el periodo reproductivo del cultivo de tomate, generó un ahorro del recurso hídrico de un 13% sin afectar la productividad, lo cual mejoró la EUA. En Países Bajos, se han obtenido EUA de hasta 66 kg.m-3 en invernaderos con el uso de calefacción e inyección de CO2 (Pardossi et al. 2016).
Sin embargo, el manejo de la irrigación en cultivos hortícolas con RD basado en UAH bajo condiciones del trópico ha sido poco estudiado y la literatura es escasa, principalmente en cultivos hidropónicos bajo invernadero. Al respecto, Pacheco (2018) en un estudio en el cultivo de tomate en fibra de coco bajo invernadero, en Alajuela, Costa Rica, demostró que el control de riego a la demanda del cultivo con un 30% de agotamiento hídrico, mostró el mejor comportamiento en el balance entre crecimiento vegetativo y generativo reflejado en un mayor rendimiento.
Por otra parte, para irrigar con precisión es determinante conocer las propiedades físico-hídricas del sustrato, y de esta forma determinar los UAH óptimos en sistemas de cultivo hidropónico según el cultivar, el manejo agronómico y las condiciones agroclimáticas. El objetivo final del manejo con RD con valores UAH, es reducir la ETc y aumentar o mantener los rendimientos para mejorar la EUA. Dicho manejo consecuentemente se manifiesta en un adecuado equilibrio entre el crecimiento vegetativo y generativo reflejado en una balanceada arquitectura del dosel vegetal sin excesivo crecimiento vegetativo o un comportamiento muy generativo (Soto et al. 2020). El objetivo del presente estudio fue evaluar el efecto de 3 niveles de agotamiento del contenido de humedad volumétrica en un sustrato de fibra de coco, sobre el comportamiento vegetativo-generativo de los cultivos de tomate y chile dulce hidropónicos en invernadero.
Materiales y métodos
El estudio se realizó entre agosto 2020 y enero de 2021 en la Estación Experimental Agrícola Fabio Baudrit Moreno, ubicada en San José de Alajuela, Costa Rica (10º 01’ N y 84º 16’ W, 840 msnm), con valores promedios de precipitación anual, temperatura y humedad relativa de 1940 mm, 22°C y 78%, respectivamente.
Se utilizó un invernadero multitúnel de hierro galvanizado, con cubierta de polietileno transparente (200 µm) y malla anti-insectos (50 mesh) en las paredes, formado por 4 módulos de 9,75 m de ancho y 50 m de largo, altura de 6 m al centro del túnel y 4 m a la canoa, orientado en dirección este-oeste. La ventilación fue pasiva en paredes laterales combinada con apertura automatizada de ventanas cenitales según la velocidad del viento e intensidad de lluvia.
Durante el experimento, dentro del invernadero se monitoreó la radiación solar global (Figura 1), (Piranómetro LI-COR Modelo LI-200SA), la temperatura (Figura 2) y la humedad del aire (Figura 3), (VAISALA Modelo HMP-35C); todos estos datos fueron almacenados en un "data logger" (Campbell Scientific. Mod. CR1000).
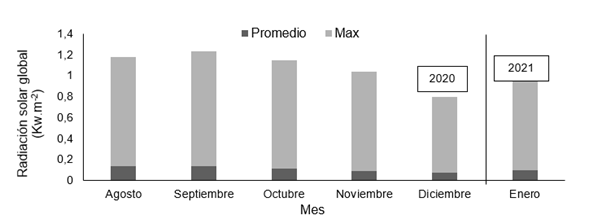
Figura 1. Valores promedio mensual de radiación solar global (Kw.m-2) media y máxima en el interior del invernadero durante el desarrollo de la investigación
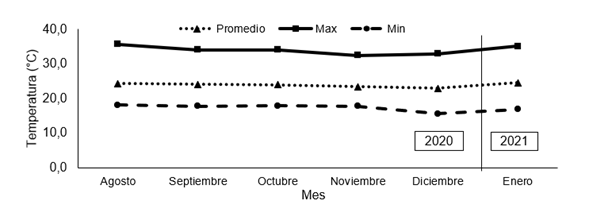
Figura 2. Valores promedio mensual de temperatura (°C) media, máxima y mínima en el interior del invernadero durante el desarrollo de la investigación.

Figura 3. Valores promedio mensual de humedad relativa (%) media, máxima y mínima en el interior del invernadero durante el desarrollo de la investigación.
Se utilizó un sistema de cultivo hidropónico abierto sin recuperación de drenaje, con un sustrato de fibra de coco en tablas comprimidas (100 cm x 14 cm x 13 cm) con cobertura plástica color blanco y un volumen aproximado 18,2 L. Las características físicas del sustrato (Tabla 1) se determinaron con la norma europea UNE-EN-13041 (AENOR 2007).
Tabla 1 Características físicas de la fibra de coco utilizada en el experimento como medio de cultivo.
| Características físicas | Fibra de coco | % Óptimo* |
| Partículas > 4 mm (%) | 54,25 | -- |
| Partículas entre 1 y 4 mm (%) | 39,00 | -- |
| Partículas < 1 mm (%) | 6,75 | -- |
| Densidad aparente (g.cm-3) | 0,096 | -- |
| Densidad real (g.cm-3) | 1,468 | -- |
| Porosidad total (%) | 93,47 | > 85% |
| Partículas sólidas (%) | 6,53 | < 15% |
*Según Ansorena 1994.
En el cultivo de chile dulce se utilizó el híbrido “Dulcitico” de crecimiento indeterminado, inicio de cosecha entre 75 y 90 días después de siembra, frutos de forma cónica color verde-rojizo, alto grado brix y un rendimiento de hasta 12,8 kg.m2 (Soto-Bravo et al. 2020). En tomate se utilizó el híbrido “Gladiador”, el cual presenta crecimiento indeterminado vigoroso, inicio de cosecha de 70 a 100 DDT, con frutos de larga vida postcosecha, multilocular, de buen cierre pistilar y con peso medio de 180-240 g y resistencia a Verticilum y Fusarium razas 1 y 2.
En ambos cultivos, la siembra se realizó por trasplante el 20 de agosto de 2020, con plántulas de 35 días después de la siembra de edad en chile dulce y 28 días en tomate, con 5 a 6 hojas verdaderas, producidas en turba (peat moss) en bandejas plásticas de 98 celdas. Se sembraron 3 plantas por tabla de fibra de coco con distancias de 33,3 cm entre plantas y de 1,5 m entre hileras para una densidad de 2 plantas por metro cuadrado.
En prácticas culturales de chile dulce se realizaron deshijas por debajo de la primera bifurcación de la planta. El amarre en chile dulce se realizó con cuerdas de nylon que sujetaban 2 tallos principales desde la primera bifurcación que lo guiaban hasta un cable acerado ubicado a una altura de 2,5 m. Todas las ramas emergentes se sujetaron con hilos de nylon a anillos colocados en las 2 cuerdas principales. En el cultivo de tomate se realizaron deshijas de los brotes laterales con el fin de formar la planta a un solo tallo, el cual fue conducido con cuerdas de nylon hasta los cables acerados del sistema de tutorado. Para el manejo de plagas y enfermedades en ambos cultivos se realizaron podas sanitarias de hojas senescentes, tallos y frutos dañados, y se realizó un monitoreo exploratorio semanal para determinar la presencia de plagas y la necesidad de aplicaciones fitosanitarias de productos biológicos, preventivos o erradicantes.
Para ambos cultivos, se aplicaron 3 tratamientos de agotamiento hídrico con una gradiente de disminución constante de la humedad volumétrica (θ) en la fibra de coco. Para ello se determinó la curva de retención de humedad volumétrica (θ) del sustrato (Figura 4) según la norma UNE-EN-13041 (AENOR 2007). Una vez definido el nivel de agotamiento hídrico, se estimó el volumen de riego para cada tratamiento con base en el porcentaje de agotamiento establecido, el volumen de sustrato (18,2 L) y un porcentaje de drenaje (15%) previamente establecido (Soto et al. 2020).
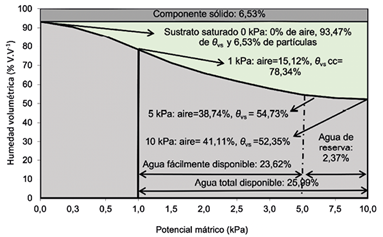
Figura 4 Contenido de partículas sólidas y curva de desorción de agua y capacidad de aireación a diferente potencial mátrico; y contenidos de agua fácilmente disponible, agua de reserva y agua total disponible en el sustrato de fibra de coco. Laboratorio de hortalizas, Estación Experimental Agrícola, Fabio Baudrit Moreno. Universidad de Costa Rica. Alajuela, Costa Rica. 2020.
En la Tabla 2 se resumen las características de los tratamientos de agotamiento hídrico utilizados en los cultivos de tomate (TT) y chile dulce (TC).
Tabla 2 Tratamientos de agotamiento utilizados en los cultivos de tomate (TT) y chile dulce (TC) en fibra de coco, expresados en porcentaje de agotamiento, potencial mátrico, porcentaje de humedad volumétrica y litros de agua agotados del ATD en cada tabla de 18,2 L de sustrato.
| Tratamiento | Porcentaje de agotamiento | Potencial mátrico (kPa) | Humedad volumétrica (%) | Litros de agotamiento |
| TT11 y TC11 | 11 | 1,70 | 69,90 | 1,54 |
| TT22 y TC22 | 22 | 2,50 | 61,45 | 3,07 |
| TT32 y TC32 | 32 | 7,50 | 53,01 | 4,61 |
El agua se suministró mediante un sistema de riego a la demanda del cultivo, que monitoreaba, en tiempo real, los volúmenes de agua aplicada (caudalímetro ARAD, Mod. SF 15) y de drenaje (pluviómetro Mod. ECRN-100, ECHO Rain) y el contenido de humedad volumétrica del sustrato con un sensor capacitivo (Decagon Devices) tipo TDR (Time domain reflectometry). Los datos recolectados se almacenaron en un “datalogger” (Decagon Devices ProCheck).
El área experimental de cada cultivo estaba compuesta por 12 hileras separadas a 1,5 m, con 17 sacos de fibra de coco/hilera. En cada saco se colocaron 3 plantas espaciadas a 0,33 m para una densidad de 2 plantas por m-2 y un total de 612 plantas por cultivo. En ambos cultivos, se utilizó un diseño experimental en bloques completos al azar, donde cada tratamiento de agotamiento hídrico tenía 3 repeticiones para un total de 9 unidades experimentales/cultivo. Cada unidad experimental estaba formada por 15 sacos de fibra de coco distribuidas en 3 hileras de 5 sacos cada una. En cada saco se sembraron 3 plantas, para un total 45 plantas por unidad experimental.
La solución nutritiva utilizada correspondió a las concentraciones de nutrientes validadas para los cultivos de tomate y chile dulce por Duarte (2019) y Monge (2020), respectivamente (Tabla 3). La conductividad eléctrica estimada que incluyó los aportes del agua de riego (CE: 0,14 dS.m-1) fue de 1,72 dS.m-1.
Tabla 3 Concentración de nutrientes en la solución nutritiva utilizada para la producción de chile dulce y tomate en fibra de coco bajo invernadero.
| Nutriente (mg.L-1) | CE | |||||||||||
| Ca | K | Mg | N | S | P | Fe | Cu | Mn | Zn | B | Mo | (dS.m-1) |
| 160 | 203 | 45 | 140 | 65 | 35,4 | 1 | 0,3 | 0,8 | 0,3 | 0,5 | 0,1 | 1,58 |
Variables respuesta
Variables morfológicas: se evaluó quincenalmente el ancho (AH) y longitud de hoja (LH) y el diámetro basal del tallo (DT); y mensualmente el número de hojas (NH), la longitud de tallo (LT) y el número de nudos (NN). El área foliar (AF), la producción de materia seca (MS) se evaluaron 3 veces a lo largo del ciclo. Dichas variables se evaluaron en 3 plantas por unidad experimental para un total de 9 plantas por tratamiento.
A. El AH se midió con cinta métrica en la parte media de la hoja.
B. La LH se calculó desde la base hasta la punta del foliolo.
C. El DT se midió con un caliper digital en la base del tallo a 2 cm por encima del nivel del sustrato.
D. El NH por planta se cuantificó mediante un conteo manual por planta.
E. La LT se midió con una cinta métrica, desde la base del tallo hasta el ápice del eje más largo.
F. El número de nudos (NN) se determinó mediante conteo manual por planta de todos los nudos formados al momento de la evaluación.
G. El AF se cuantificó al remover manualmente las hojas de cada planta y utilizar un medidor de área (LI-COR MODEL 3100).
H. La producción de MS se evaluó en 3 muestreos de 3 etapas fenológicas: 1-fase vegetativa, 2- inicio de floración y cuaje de frutos y 3- al final de ciclo de producción. En cada evaluación se recolectaron 2 plantas por repetición para un total de 6 por tratamiento, se cuantificó la masa fresca, se colocaron en bolsas de papel y se secaron en estufa a 70ºC hasta obtener peso constante.
Variables fisiológicas: se evaluó la conductancia estomática (gs), la evapotranspiración del cultivo (ETc) y la eficiencia de uso de agua (EUA). La gs (mmol.m-2.s-1) se midió con un porómetro (Decagon, Mod. SC-1) cada 15 días en 3 plantas por repetición y un total de 9 plantas por tratamiento. La ETc (L.m-2) se determinó semanalmente mediante el método de balance de agua, a partir de los volúmenes (L.m-2) de riego (R) y de drenaje (D) y la diferencia entre el contenido de humedad volumétrica del sustrato (∆θ= θ i- θ f) al inicio (θ i) y final (θ f) de cada semana, según ecuación descrita por Soto (2018).
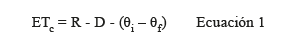
Finalmente, la eficiencia de uso de agua (EUA) se determinó como el cociente entre rendimiento de fruto fresco (kg.m-2) y la ETc (m3.m-2), tanto para el rendimiento comercial como para el rendimiento total (Salazar et al. 2014).
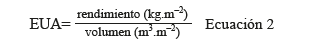
Variables productivas: durante 14 semanas del periodo de cosecha de ambos cultivos, se evaluó semanalmente el rendimiento de frutos en 12 plantas por repetición y 36 por tratamiento. Se evaluó el Rc con la clasificación de los frutos en 3 categorías de calidad según su peso, en primera (F1), segunda (F2) y tercera (F3). En chile dulce se clasificaron los frutos en 3 calidades: F1 con pesos entre 107 y 185 g sin deformaciones ni manchas, F2 con pesos entre 81 y 132 g y F3 con masas de 46-80 g con deformaciones y manchas leves según Monge (2020). En tomate se clasificaron los frutos en F1, F2 y F3 con diámetros aproximados a 8 cm, 6 cm y 4 cm, respectivamente.
Finalmente, para ambos cultivos se contabilizaron los frutos de desecho (Fd) con daños físicos y mecánicos, para estimar el rendimiento total de frutos (Rt) como la suma de Rc más desecho.
Análisis estadístico. El análisis de los datos se realizó mediante el programa estadístico R y con uso de las librerías: agricolae, tidyverse, readxl, stringr, DT, car, carData y ggplot2 (Möhring et al. 2021). En primera instancia, se procedió a realizar un análisis de normalidad (Kolmogorov-Smirnov) y homocedasticidad (Bartlett) para las variables teóricamente normales. Cuando se cumplían los supuestos anteriores, se realizó un análisis de varianza y si se detectaron efectos significativos, se realizó una comparación múltiple de medias (p<0,05) según Di Rienzo et al. (2002) para contrastar las variaciones entre los tratamientos. Seguidamente se realizaron contrastes todos contra todos al ser solo 3 tratamientos. Cabe resaltar que los mismos no son ortogonales por lo tanto se utilizó la corrección de Bonferroni como α/k, donde k fue el número de comparaciones a realizar.
Resultados y discusión
Comportamiento morfológico. En tomate, los tratamientos de agotamiento hídrico afectaron (p<0,05) el AH, el DT, el AF y la producción de MS, mientras que no hubo efectos estadísticamente significativos (p<0,05) sobre LH, NH, LT y NN (Tabla 4).
El promedio de AH en el cultivo de tomate mostró una tendencia a incrementar conforme se redujo el estrés hídrico, donde el TT11 fue similar a TT22 y superior a TT32, sin diferencias entre TT22 y TT32 (Tabla 4). El patrón en el comportamiento del AH, aun cuando no hubo diferencias entre tratamientos en la LH, fue consecuente con un incremento en el AF conforme descendió el estrés hídrico. El TT11 obtuvo un 12,5% y un 26% más de área foliar con respecto a con TT22 y TT32 (Tabla 4).
Tabla 4 Valores promedio de ancho de hoja, área foliar, materia seca, diámetro del tallo, número de hojas, longitud de la hoja, número de nudos y longitud del tallo en plantas de tomate sometidas a 3 tratamientos de agotamiento hídrico de 11% (TT11), 22% (TT22) y 32% (TT32) del contenido de humedad volumétrica del sustrato.
| Variable respuesta | Tomate | ||
| TT11 | TT22 | TT32 | |
| Ancho de hoja (cm) | 44,28 a | 42,11 ab | 40,25 bc |
| Área foliar (cm2) | 11212 a | 9800 b | 8219 c |
| Materia seca (g) | 222 a | 185 b | 157 bc |
| Diámetro de tallo (mm) | 41,95 a | 38,79 bc | 37,46 c |
| Número de hojas | 79 a | 79 a | 79 a |
| Longitud de hoja (cm) | 49,66 a | 46,05 a | 44,71 a |
| Número de nudos | 78 a | 78 a | 78 a |
| Longitud de tallo (LT) | 576,6 a | 577,2 a | 575,6 a |
Letras diferentes en la misma fila indican diferencias significativas entre tratamientos (p˂0,05).
Consecuentemente, a menor agotamiento hídrico y mayor AF en T11, la producción de MS fue superior, probablemente debido a una mayor capacidad fotosintética soportado por una mayor fuente de AF. En MS, el TT11 produjo un 16% más de MS respecto a TT22 y 30% más con respecto a TT32; donde estos 2 últimos no mostraron diferencias entre sí (Tabla 4). Finalmente, el DT de plantas de tomate, fue menor conforme incrementó el agotamiento hídrico, donde el TT11 fue en promedio un 7,5% y un 10,7% superior a TT22 y TT32, respectivamente, sin diferencias entre estos últimos (Tabla 4).
En el cultivo de chile dulce, todas las variables morfológicas fueron estadísticamente diferentes entre tratamientos. Las variables relacionadas con el tamaño de la fuente de la planta, tales como el AH, LH y NH, fueron estadísticamente superiores conforme disminuyó el estrés hídrico. Consecuentemente, el AF y la producción de MS fue superior en el TC11 respeto a TC22 y TC32. De forma similar al cultivo del tomate; el AH fue la variable que mantuvo un patrón consecuente con el comportamiento del AF, con una tendencia al incremento estadísticamente significativa, conforme se redujo el nivel de estrés hídrico (Tabla 5).
Tabla 5 Valores promedio de ancho de hoja, área foliar, materia seca producida, diámetro del tallo, número de hojas, longitud de la hoja, número de nudos y longitud del tallo en plantas de chile dulce sometidas a 3 tratamientos de agotamiento hídrico de 11% (TC11), 22% (TC22) y 32% (TC32) del contenido de humedad volumétrica del sustrato.
| Variable respuesta | Chile dulce | ||
| TC11 | TC22 | TC32 | |
| Ancho de hoja (cm) | 11,44 a | 10,39 b | 9,64 c |
| Longitud de hoja (cm) | 21,39 a | 20,84 b | 20,13 b |
| Número de hojas | 1017 a | 844 b | 765 bc |
| Área foliar (cm2) | 11212 a | 9800 b | 8219 c |
| Materia seca (g) | 222 a | 185 b | 157 bc |
| Número de nudos | 907 a | 794 b | 725 bc |
| Longitud de tallo (cm) | 310,3 a | 286,1 b | 276,2 b |
| Diámetro de tallo (mm) | 34,80 a | 32,29 b | 29,58 b |
Letras diferentes en la misma fila indican diferencias significativas entre tratamientos (p˂0,05).
Similar al cultivo de tomate, el DT de plantas de chile dulce fue menor conforme disminuyó el agotamiento hídrico (Figura 3C), donde TC11 fue superior un 7% que TC22 y 15 % más que TC32, estos 2 últimos fueron iguales entre ellos (Tabla 5).
En ambos cultivos, las plantas con el menor agotamiento hídrico (TT11 y TC11) produjeron mayor biomasa aérea (MS) que los tratamientos con mayor agotamiento hídrico. Esto estuvo relacionado principalmente con el AH que repercutió positivamente en mayor tamaño de fuente reflejado en una superior AF. En respuesta a un mayor nivel de agotamiento hídrico, los tratamientos con 22 y 32% redujeron el AH, LH y el NH, variables que determinan el tamaño del AF foliar, la cual está directamente relacionada con la transpiración del cultivo y con la producción de biomasa (MS).
El anterior comportamiento ha sido evidenciado por estudios realizados en chile dulce por González et al. (2007) y Quesada (2015). El primero demostró que el déficit hídrico durante la etapa productiva redujo el AF y consecuentemente la producción de biomasa, mientras que Quesada (2015) en el cultivo bajo invernadero observó una tendencia a disminuir el AF conforme se aumentaba el agotamiento, lo que consecuentemente redujo la producción de MS.
En tomate, se ha demostrado que el estrés hídrico inhibe el crecimiento y limita la expansión foliar (García et al. 2004), debido a que el proceso de expansión celular es mucho más sensible a dicho estrés que el proceso de división celular (Florido y Bao 2014). Además, el déficit de agua reduce el crecimiento de la raíz y consecuentemente el crecimiento de la parte aérea (Potters et al. 2007), lo que afectó dicha relación.
Los resultados obtenidos en este estudio demostraron que el estrés hídrico en ambos cultivos inhibió el crecimiento vegetativo limitado por la expansión foliar (AH, LH, AF), lo que afecta consecuentemente el balance vegetativo-generativo del cultivo. Estas modificaciones en la arquitectura del dosel vegetal responden principalmente al manejo del riego y a su efecto sobre la relación entre los componentes vegetativo aéreo y el sistema radical (Potters et al. 2007). Esto debido a que las plantas requieren un sistema radicular suficientemente desarrollado para abastecer la demanda hídrica de los órganos aéreos (Peleg et al. 2011). Las raíces crecen adecuadamente cuando la humedad es óptima y uniforme en todo el volumen del sustrato, lo que favorece un buen volumen radicular.
Un manejo del riego con alto nivel de agotamiento hídrico implica riegos poco frecuentes que afectan una distribución uniforme de la humedad en el perfil del sustrato, lo que consecuentemente limita el crecimiento radical y afecta la relación entre el dosel vegetal y sistema radicular de la planta (Potters et al. 2007), como respuesta adaptativa a una menor capacidad radicular para abastecer la demanda transpiratoria. Al respecto, Bar-Tal et al. (1995) demostraron que en chile hidropónico bajo invernadero al reducir el volumen del contenedor se limitó el crecimiento radical y consecuentemente el AF.
En el presente estudio, las plantas de ambos cultivos modificaron el AF al variar la forma de la hoja, en términos de AH en tomate y el AH, LH y NH en chile dulce, en respuesta al nivel de estrés hídrico, como una forma de evitar la pérdida excesiva de agua, ya que el AF está estrechamente relacionada con la tasa de transpiración del cultivo (Taiz y Zeiger 2003, Qiu et al. 2013, Di Benedetto y Tognetti 2016).
El DT es una variable que está directamente relacionada con el manejo del riego de los cultivos, ya que su contracción y expansión es una respuesta morfofisiológica al estado hídrico de las plantas (Kumar et al. 2017). En el presente estudio, el DT de las plantas de chile dulce y tomate con mayor agotamiento hídrico de 22 y 32%, fueron en promedio un 10% más delgados que el tratamiento de 11%. Al respecto, Kumar et al. (2017) demostraron que el DT en plantas de tomate disminuyó conforme se redujo el aporte hídrico, mientras que Taiz y Zeiger (2003) demostraron que la variación en el diámetro basal del tallo es un indicador de estrés hídrico en el cultivo de tomate.
Ante situaciones de estrés hídrico la planta modifica el DT, que está relacionado con el estado hídrico del cultivo. Además, modifica el NH, el AH y la LH que determinan su AF, la cual está directamente relacionada con la transpiración del cultivo (García et al. 2004, Dobermann 2007). Al respecto, Pacheco (2018) no obtuvo efecto ante 3 niveles de agotamiento hídrico (10, 20 y 30%) sobre el NH y LT en plantas de tomate hidropónico en invernadero, probablemente debido a que el nivel de estrés hídrico no fue suficiente para afectar los procesos fisiológicos de la planta.
En el cultivo de chile dulce en invernadero, estudios realizados por Gallardo et al. (2004), González et al. (2007) y Jara (2018), encontraron que el DT se redujo conforme aumenta el nivel de estrés hídrico, lo cual evidenció así que dicho parámetro es un indicador del estrés hídrico.
Conductancia estomática (gs). Las plantas regulan la gs mediante señales que involucran algunas fitohormonas, principalmente ABA, como mecanismo para mitigar el estrés hídrico y evitar la deshidratación.
En la presente investigación, en ambos cultivos, los tratamientos con 11% de agotamiento hídrico, obtuvieron los mayores valores de gs, con una tendencia decreciente conforme aumentó el porcentaje de agotamiento hídrico (Figura 5). Asociado a la reducción del AF, el déficit hídrico promovió una reducción del potencial hídrico de las hojas, lo cual actúa como una señal hidráulica-hormonal que induce al cierre parcial de estomas (Khapte et al. 2019), manifestándose en una reducción de la gs, tal como como se evidenció en el presente estudio.
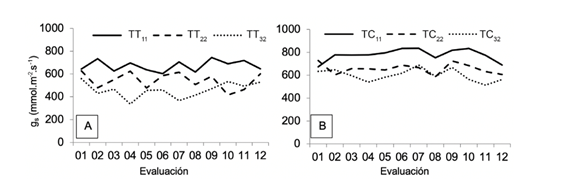
Figura 5 Valores promedio de conductancia estomática (mmol.m-2.s-1) en hoja, en tratamientos de 3 porcentajes de agotamiento hídrico de 11, 22 y 32% del contenido de humedad volumétrica del sustrato, en plantas de (A) tomate (TT11, TT22, TT32) y (B) de chile dulce (TC11, TC22, TC32 ).
En el cultivo de tomate, a lo largo del periodo, la gs fluctuó entre los 400 y 800 mmol.m-2.s-1 (Figura 4A), con diferencias significativas entre todos los tratamientos. El tratamiento TT11 obtuvo los mayores valores de gs, con una tendencia creciente conforme disminuyó el porcentaje de agotamiento hídrico, con valores promedios durante el ciclo de cultivo de 677,54 mmol.m-2.s-1, 543,74 mmol.m-2.s-1 y 463,67 mmol.m-2.s-1, para los tratamientos TT11, TT22 y TT32, respectivamente.
En el caso del cultivo de chile dulce la gs fluctuó entre 400 a 900 mmol.m-2.s-1 (Figura 4B), con diferencias significativas entre todos los tratamientos. El TC11 presentó los mayores valores de gs durante el ciclo de cultivo con 778,24 mmol.m-2.s-1, seguido de TC22 con 681,59 mmol.m-2.s-1 y finalmente TC32 con 611,35 mmol.m-2.s-1. En ambos casos las plantas sometidas al estrés hídrico regularon la apertura estomática principalmente debido a la acumulación de ABA, esta acumulación de fitohormonas ha sido ampliamente estudiada y está estrechamente ligada a una reducción en la gs, la cual se puede utilizar como una medida indirecta, ya que es el mecanismo principal para mitigar la pérdida de agua por evapotranspiración. En el cultivo de tomate valores de gs de 800 mmol.m-2.s-1 son un indicativo de que no se está dando una adecuada circulación del agua y fotosíntesis (Davies et al. 2000, Campos et al. 2009, Giuliani et al. 2018).
La regulación en la pérdida de agua mediante el cierre parcial de estomas genera reducciones desde 33 hasta 44% en la gs en cultivares de tomate bajo déficit hídrico (Savic et al. 2009). En este estudio, en tomate se determinó una disminución en la gs de hasta un 31% en el tratamiento TT32 respecto a TT11. En otro estudio realizado por Nemeskéri et al. (2019), al aplicar riego deficitario obtuvieron valores de gs (893,4 mmol.m-2.s-1) inferiores a un cultivo de tomate bien irrigado (950 mmol.m-2.s-1).
En el cultivo de chile dulce, Ismail et al. (2002) encontraron una reducción de hasta 160 mmol.m-2.s-1 en las plantas con mayor nivel de estrés hídrico. Lo cual es coincidente con los resultados obtenidos en la presente investigación, ya que la gs se redujo de TC32 a TC11 en 146 mmol.m-2.s-1 en los picos más altos y de 128 mmol.m-2.s-1 en los más bajos, debido probablemente a elevadas concentraciones de ABA. En otro estudio, Aladenola y Madramootoo (2014), obtuvieron valores de gs similares a los obtenidos por Ismail et al. (2002), con reducciones desde 15% hasta 25% en los tratamientos más agotados.
Evapotranspiración de los cultivos (ETc) de tomate y chile dulce. En ambos cultivos, la ETc promedio total (L.m-2) no fue estadísticamente diferente (p<0,05) entre tratamientos (Figura 6). No obstante, se evidenció una tendencia a incrementar la ETc conforme acrecentaba el agotamiento hídrico.
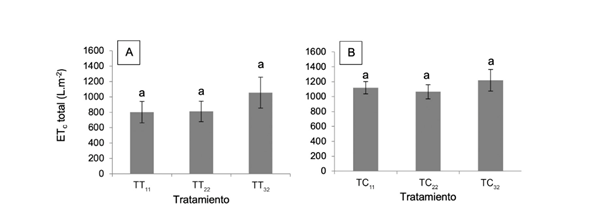
Figura 6 Valores promedio de evapotranspiración del cultivo (ETc) de (A) tomate (TT) y (B) chile dulce (TC) sometidos a 3 tratamientos de agotamiento hídrico de 11% (TT11, TC11), 22% (TT22, TC22) y 32% (TT32, TC32) del contenido de humedad volumétrica en el sustrato. Letras diferentes en la misma columna indican diferencias significativas entre tratamientos (p˂0,05).
Esto probablemente fue debido a que, según el tratamiento hídrico, varió el manejo del riego. Al final del día el volumen total de riego aportado (L.m-2) fue similar entre tratamientos, aunque con volúmenes y frecuencias de riego diferentes, ya que conforme incrementó el nivel de agotamiento, aumentó el volumen (cuánto) y la frecuencia de riegos (cuándo). Al respecto, Rodríguez et al. (2014) demostraron que el manejo del riego influye directamente en el comportamiento morfológico y productivo del cultivo.
Comportamiento productivo
En ambos cultivos, no hubo efecto (p>0,05) de los tratamientos sobre el rendimiento de frutos total y de desecho. Sin embargo, hubo diferencias estadísticamente significativas (p<0,05) sobre el rendimiento comercial, debido a las diferencias en la partición de frutos según calidad (Tabla 6).
Tabla 6 Rendimiento total, comercial y por calidad de frutos por planta, en los cultivos de tomate (TT) y (B) chile dulce (TC) sometidos a 3 tratamientos de agotamiento hídrico de 11% (TT11, TC11), 22% (TT22, TC22) y 32% (TT32, TC32) del contenido de humedad volumétrica en el sustrato.
| Componente rendimiento | Tomate | Chile dulce | |||||
| TT11 | TT22 | TT32 | TC11 | TC22 | TC32 | ||
| Primera (I) (kg.pl-1) | 1,24 a | 0,92 b | 0,92 b | 1,63 a | 1,51 ab | 1,30 bc | |
| Segunda (II) (kg.pl-1) | 4,83 a | 4,70 ab | 3,90 bc | 1,71 a | 1,53 ab | 1,31 bc | |
| Tercera (III) (kg.pl-1) | 1,81 a | 1,80 a | 1,79 a | 1,60 a | 1,54 ab | 1,49 bc | |
| Desecho (kg.pl-1) | 0,40 a | 0,59 a | 0,89 a | 0,70 a | 0,88 a | 0,99 a | |
| Comercial (kg.pl-1) | 7,88 a | 7,42 ab | 6,61 bc | 4,94 a | 4,58 ab | 4,10 c | |
| Total (kg.pl-1) | 8,28 a | 8,01 a | 7,50 a | 5,64 a | 5,46 a | 5,09 a | |
Letras diferentes en la misma fila indican diferencias significativas entre tratamientos (p˂0,05).
En el cultivo de tomate se encontraron diferencias en cuanto a la producción de frutos de I y II calidad, no así en las calidades III y desecho. El tratamiento TT11 obtuvo el mayor rendimiento en calidad I, con 320 g más por planta que los tratamientos TT22 y TT32, estos últimos fueron iguales entre sí (Tabla 6). Dicha diferencia equivale a 6,4 t.ha-1 más de producción de frutos de I calidad.
En producción de II calidad, el TT11 obtuvo 930 g más por planta con respecto a TT32 (Tabla 6), no así con respecto a TT22, pues estos fueron iguales. En t.ha-1 dicha diferencia fue equivalente a 18,6 t.ha-1 más de frutos de II calidad. Los tratamientos TT22 y TT32 fueron estadísticamente iguales. En rendimiento comercial (I, II y III), el TT11 obtuvo 1270 g más que TT32, equivalente a una diferencia de 25,4 t.ha-1.
En el cultivo de chile, el rendimiento total de frutos no fue estadísticamente diferente entre tratamientos. En rendimiento de frutos de I, II y III calidad, el TC11 fue estadísticamente superior a TC32 y similar a TC22. Los tratamientos TC22 y TC32 fueron estadísticamente iguales en todas las calidades de fruta. El TC11 obtuvo un rendimiento de frutos de 330 g de I, 400 g de II y 110 g de III por planta, superior a TC32 (Tabla 5), equivalente a un aumento de 16,8 t.ha-1.
Eficiencia de uso de agua (EUA)
En la presente investigación, tanto en tomate como en chile dulce, los tratamientos con 11 y 22% de agotamiento hídrico obtuvieron una EUA similar, y superior a los tratamientos con un agotamiento de 32%. En el cultivo de tomate, el TT11 obtuvo un rendimiento 47,4% superior a TT32; mientras que, en chile dulce, el TC11 obtuvo un rendimiento 57,5% superior a TC32 (Figura 7).
Figura 7 Eficiencia en el uso de agua con base en rendimiento comercial de los cultivos de tomate (A) y de chile dulce (B), sometidos a 3 tratamientos de agotamiento hídrico de 11% (T11), 22% (T22) y 32% (T32) del contenido de humedad volumétrica del sustrato. Letras diferentes indican diferencias significativas entre tratamientos (p˂0,05). Letras diferentes en la misma columna indican diferencias significativas entre tratamientos (p˂0,05).
Considerando que la EUA (kg.m-3) es la relación entre el rendimiento de frutos (kg.m-2) y la ETc del cultivo (m3.m-2), esta mayor EUA fue producto de un mayor rendimiento comercial en las plantas con 11 y 22% de agotamiento respecto a 32%, ya que la ETc no difirió entre tratamientos. A mayor agotamiento hídrico, la planta tiene que ejercer mayor succión (kPa) para extraer el agua del sustrato, lo que implica un mayor gasto energético a expensas del rendimiento. Consecuentemente, se generan cambios morfofisiológicos para adaptarse a la condición de agotamiento hídrico, que según su intensidad puede manifestarse en aumentos o disminuciones de la productividad (Rodríguez et al. 2014, Soto 2018).
Lo anterior se debe a que, en ambos cultivos, un mayor nivel de agotamiento hídrico tuvo como consecuencia un desbalance entre los componentes vegetativos y reproductivos (fuente/sumidero), lo que afectó el suministro de foto-asimilados para el buen llenado de frutos. Las plantas como respuesta a la condición de estrés adoptaron un comportamiento más reproductivo con el fin de preservar la especie (Soto 2018). Además, como respuesta al estrés hídrico, la planta aborta órganos reproductivos tales como flores y frutos recién cuajados, sosteniendo solo los que puede llenar (Ferrara et al. 2011, Tanaskovik et al. 2017). Por el contrario, los tratamientos con menor agotamiento hídrico de 11 y 22%, evidenciaron un mejor balance entre crecimiento vegetativo y generativo, que se tradujo en un mayor rendimiento comercial de frutos de primera y segunda calidad.
El contenido óptimo de humedad en el sustrato conocido como umbral de riego (UR), depende principalmente del clima, del cultivo y del tipo de sustrato, entre otros factores. La aplicación de estrategias de riego deficitario puede tener diferentes efectos sobre el cultivo. Por tanto, una apropiada estrategia de riego se refiere a la utilización de un valor de UR que permita incrementar la EUA (Kirda et al. 2004), así como reducir la ETc y/o incrementar el rendimiento del cultivo (Ćosić et al. 2015). En la presente investigación, los tratamientos TT11 y TC11 permitieron mejorar la EUA en rendimiento comercial, ya que se obtuvo un incremento del rendimiento en frutos de I y II calidad, aun cuando la ETc fue similar entre tratamientos. Coincidentemente, en un estudio realizado en Jordania en tomate, con el uso de riegos deficitarios se obtuvo un aumento de la EUA al reducir la ETc, a pesar de una reducción en el rendimiento comercial (Mahadeen et al. 2011). De forma similar en España, la EUA incrementó, ya que se obtuvo un apreciable ahorro de 28% en agua con una reducción de 16,4% en el rendimiento comercial (Lahoz et al. 2016). En otros estudios, Khapte et al. (2019) reportaron que, a pesar de las reducciones en rendimiento debido a un descenso en el número y tamaño de frutos, el déficit hídrico mejoró la calidad de frutos en firmeza, contenidos de licopeno, sólidos solubles, ácido ascórbico y la acidez, como consecuencia de una redistribución de los foto-asimilados en menos sumideros.
Los efectos de una estrategia de riego deficitario también varían según la etapa fenológica del cultivo. En estudios realizados con RD en el cultivo de tomate durante la etapa de floración, se reportaron valores de EUA de 19,2 kg.m-3 (Nangare et al. 2016) y de 17,45 kg.m-3 (Patanè et al. 2011) producto de un incremento en el rendimiento sin reducciones en el consumo de agua. Dichas EUA fueron apreciablemente inferiores a los valores obtenidos con 11 y 22% de agotamiento hídrico en el cultivo de tomate (kg.m-3) del presente estudio.
En Jordania se desarrolló un ensayo con agotamientos similares a los de esta investigación, en donde se utilizaron diferentes niveles de RD clásico durante todo el periodo productivo del cultivo de tomate. Los autores reportaron una reducción en el rendimiento, sin embargo, aumentó la EUA con 17 kg.m-3 y 23,9 kg.m-3 en 2 diferentes cultivares (Mahadeen et al. 2011). Dichos valores de eficiencia fueron similares a los obtenidos en esta investigación, donde se obtuvo una máxima EUA de 37,32 kg.m-3 en el tratamiento TT11, luego una de 33,49 kg.m-3 en el TT22 y finalmente una de 25,22 kg.m-3 en el TT32.
En el cultivo de chile dulce, de forma similar al tomate, el rendimiento disminuyó conforme aumentó el nivel de agotamiento producto de modificaciones morfofisiológicas y reproductivas, que afectaron la EUA. Al respecto, Karam et al. (2009) reportó que reducciones del requerimiento hídrico de 80, 60 y 40% redujo el rendimiento en 11,3; 12,2 y 38,2%, respectivamente. Finalmente, Aladenola y Madramootoo (2014), en un estudio con tratamientos de RD clásicos durante la etapa productiva, demostraron que conforme aumentaba el nivel de agotamiento disminuía el rendimiento comercial. Así los valores de EUA fluctuaron desde 2,9 kg.m-3 hasta 31,6 kg.m-3.
Conclusiones
En chile y tomate el nivel de agotamiento hídrico no afectó el rendimiento total de frutos.
Los tratamientos con 11 y 22% de agotamiento hídrico incrementaron el rendimiento comercial debido a una mayor producción de frutos de primera y segunda calidad y menos desecho, lo cual se reflejó en una mayor eficiencia de uso de agua.
El incremento del rendimiento comercial está relacionado con un mejor balance entre crecimiento vegetativo y reproductivo debido a una mejor distribución de asimilados para el llenado de frutos de mayor calidad.

