











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
Uno de los grandes desafíos del nuevo milenio para la humanidad es la mitigación y adaptación al cambio climático. De acuerdo con el avance de las investigaciones referentes a la emisión de gases de efecto invernadero, se recomienda tomar las medidas necesarias para reducir los impactos a escala regional y global.
Los bosques son los ecosistemas terrestres más diversos y cumplen funciones ambientales y ecológicas, sociales y económicas. Los mismos ofrecen beneficios directos e indirectos al entorno, uno de los principales es el servicio de regulación que garantiza la provisión de agua y el equilibrio de la temperatura. Los servicios ecosistémicos de regulación del clima, de la calidad del agua y la calidad del aire son otros valiosos aportes para la sociedad. Los ecosistemas, en general, mantienen flujos de materia y energía entre sí y con otros ecosistemas; tales flujos afectan directamente la temperatura y las precipitaciones, y en la medida en que existe mayor evapotranspiración aumenta la precipitación a escala local. Igualmente, los ecosistemas y los agroecosistemas son sumideros de CO2, un gas de efecto invernadero que en altas concentraciones afecta la temperatura (Corredor et al. 2012).
Los bosques de galería o ribera, también conocidos como sotos, poseen una vegetación tan frondosa que cubre por entero un río, su nombre proviene del hecho de que su vegetación cubre al río con una especie de túnel, como en la galería de una mina. Estos bosques son formaciones de árboles, arbustos y especies herbáceas que se desarrollan en los márgenes de los ríos, extendiéndose a los riachuelos que los alimentan e inclusive a algunas zonas de drenajes de las aguas de escurrimiento, en redes continuas de vegetación natural de gran importancia ecológica, ya que controlan la erosión de los márgenes, juegan un importante rol en el ciclo del agua y la regulación ambiental, pero también se comportan como corredores de biodiversidad, que comunican a diferentes ecosistemas (Stevenson y Rodríguez 2008).
Cuellar y Salazar (2016) sostienen que la capacidad de capturar carbono en el suelo a una profundidad de 50 cm, en los sistemas evaluados está en función de: textura y estructura, pH, nutrientes presentes, tipo de agregado, precipitación que interviene en el crecimiento de la vegetación arbórea y en la descomposición de la hojarasca, temperatura y profundidad del suelo.
De acuerdo con la Evaluación de los Ecosistemas del Milenio, los servicios ecosistémicos (SE) se pueden definir como todos aquellos beneficios que la sociedad obtiene de los ecosistemas; un concepto cada vez más aplicado a la conservación del ambiente, el bienestar humano y la implicación de las intervenciones antropogénicas en el medio natural (MEA 2005).
Se considera que los bosques tropicales del mundo, debido a su amplia distribución, elevada diversidad y contribución a funciones clave del planeta como la regulación climática e hidrológica, proveen servicios ecosistémicos críticos (Balvanera 2012).
La regulación de la calidad del agua también proviene de ecosistemas que proveen una gran variedad de funciones hidrológicas importantes para el bienestar humano. Esas funciones se convierten en bienes y servicios ecosistémicos cuando son valoradas en términos del bienestar y el desarrollo de la sociedad. Los servicios hidrológicos incluyen la regulación de caudales para mitigar inundaciones, la recarga de acuíferos que mantienen caudales durante la época seca, la purificación del agua y el control de la erosión (Corredor et al. 2012).
Los servicios de regulación están relacionados con la capacidad de los ecosistemas para regular procesos ecológicos esenciales y sostener sistemas vitales a través de ciclos biogeoquímicos (Camacho Valdez y Ruiz Luna 2012).
Esta investigación apoya el desarrollo de políticas sociales y ambientales que garanticen la puesta en marcha de proyectos ecosistémicos sostenibles. De conformidad con la Ley N° 4241/10 la propuesta de estudio abarca la importancia de los ecosistemas naturales con miras al desarrollo sustentable (INFONA, PNUD, FMAM 2019).
Al respecto, el bosque de galería del Campus de la Universidad Nacional de Asunción representa un potencial ecosistémico importante para varios pobladores de las zonas aledañas. Dicho ecosistema forestal de relevancia proporciona bienes y servicios de almacenamiento de carbono, regulación del microclima local y provisión de agua para la conservación ecológica de la sub cuenca del arroyo San Lorenzo, Paraguay.
Según lo anterior, el objetivo de esta investigación fue analizar el servicio ecosistémico de regulación relacionado con el carbono almacenado y el aprovicionamiento de agua y nutrientes del bosque de galería del Arroyo San Lorenzo, Paraguay.
Materiales y métodos
Localización. El trabajo de investigación se llevó a cabo en el Campus de la Universidad Nacional de Asunción, ubicada en la ciudad de San Lorenzo a 11 km de Asunción, capital de la República del Paraguay, entre las coordenadas 25°20'18'' Lat. Sur y 57°31'04'' Lat. Oeste, a 125 m sobre el nivel del mar; en el denominado bosque de galería del Campus de la Universidad Nacional de Asunción.
El bosque de galería del Campus de la Universidad Nacional de Asunción presenta una superficie aproximada de 5,3 ha (384 m x 140 m) (Benítez 2018).
Variables estudiadas. Fueron instaladas 4 parcelas (750m2) cada una en forma aleatoria en 4 puntos más representativos del lugar de estudio, considerándose el tipo de vegetación y cobertura de suelo. Para evitar el efecto de borde, todas las parcelas se ubicaron a una distancia igual o mayor a 10 m del borde.
Las variables estudiadas fueron: la composición florística, las propiedades físicas y químicas del suelo, el carbono acumulado en el estrato arbóreo aéreo, radicular y suelo y la velocidad de infiltración media del agua.
Composición florística. El inventario de composición florística se realizó con el registro de los siguientes datos en planilla: diámetro a altura de pecho igual o mayor a 10 cm, reportes de nombre científico, familia, altura total y de fuste, estado sanitario.
Propiedades físicas y químicas del suelo. Para el muestreo de suelo se extrajeron 8 muestras (4 submuestras a una profundidad de 0 a 10 cm y 4 de 10 a 30 cm) de forma aleatoria en diferentes sectores del bosque de galería, en las mismas parcelas en donde se realizó el inventario forestal, por medio de una pala de punta. Luego se procedió a la homogenización y un secado preliminar para obtener 2 muestras compuestas por sector. Para la determinación de la densidad aparente se tomaron 4 muestras aleatorias por parcela con cilindros metálicos de 100 cm3 a una profundidad de 0 a 10 cm y de 10 a 30 cm, con un total de 32 cilindros con las muestras de suelo correspondientes.
Para la interpretación de los resultados obtenidos se compararon los valores de los nutrientes en el suelo con los parámetros tomados de referencia por el Laboratorio de Suelos y Ordenamiento Territorial de la Facultad de Ciencias Agrarias de la Universidad Nacional de Asunción, los mismos se exponen en las Tablas 1 y 2.
Tabla 1 Interpretación del contenido de nitrógeno total en porcentaje.
| Niveles de Nitrógeno | % N |
| Bajo | 0,06-0,10 |
| Medio | 0,11-0,20 |
| Alto | 0,21-0,30 |
| Muy Alto | 0,30- ˃0,30 |
Tabla 2 Determinación del contenido de materia orgánica y nutrientes del suelo.
| Nivel | Materia orgánica | P | Ca+2 | Mg+2 | K+ | Al+3+H+ |
| Bajo | < 1,2 | < 12 | < 2,51 | < 0,4 | < 0,12 | < 0,4 |
| Medio | 1,2 - 2,8 | 12 - 30 | 2,51 - 6,0 | 0,4 - 0,8 | 0,13 - 0,17 | 0,4 - 0,9 |
| Alto | > 2,8 | > 30 | > 6,0 | > 0,8 | > 0,17 | > 0,9 |
En la Tabla 3, se muestran los valores tenidos en cuenta para la determinación del pH del suelo.
Tabla 3 Determinación del pH del suelo.
| pH | |
| < 5,5 | Ácido |
| 5,6 - 6,4 | Ligeramente ácido |
| 6,5 - 7,4 | Neutro |
| > 7,4 | Alcalino |
Velocidad de infiltración media del suelo. Para el cálculo de la velocidad de infiltración media se realizaron 2 ensayos en diferentes sitios del bosque de galería mediante la instalación de cilindros (infiltrómetros de doble anillo). Se procedió al llenado del cilindro interior con carga hidráulica constante de 8 cm y medición de la altura del nivel cada 2, 7, 10, 17, y 32 minutos, al considerar que el descenso del agua fuera notorio entre cada medición, hasta que el descenso del nivel se presentara constante.
Se utilizó la siguiente ecuación, según Pizarro Tapia et al. (2008).
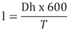
Donde
I = Velocidad de infiltración en (mm/h)
Dh = Diferencial de altura (mm)
t = Tiempo (min)
Carbono acumulado en el estrato arbóreo aéreo, radicular y suelo. Para estimar el carbono acumulado en el estrato arbóreo se midieron los árboles con DAP ≥10 cm con forcípula, altura de fuste y total con hipsómetro de cada individuo en cada parcela. Asimismo, para la determinación del carbono en suelo se procedió a la aplicación del Método de Walkley-Black, posteriormente se utilizaron las fórmulas enunciadas en la Tabla 4.
Tabla 4 Fórmulas empleadas para la estimación de carbono.
| Variables | Fórmulas | Descripción |
| Carbono aéreo total (tC.ha-1) | CAT= BT * 0,5 | CAT = Carbono aéreo total en toneladas de carbono (tC) BT = Biomasa total en tonelada (t) Factor de conversión = 0,5 (la materia seca contiene en promedio 50% de carbono almacenado, según la (IPCC 2006) |
| Carbono radicular (tC.ha-1) | CR= 0,24 * (CA) | CR = Carbono radicular en tonelada por hectárea CA = Carbono aéreo en tonelada por hectárea (t.ha-1) Relación media entre biomasa bajo/sobre el suelo de 0,24 para bosque (IPCC 2006) |
| Carbono total (tC.ha-1) | CT= CAT + CR | CT = Carbono aéreo total en toneladas de carbono (tC) CR= Carbono radicular en tonelada de carbono (tC) |
| Carbono equivalente (tC.ha-1) | CO2e= CT * 3,667 | CO2e = Dióxido de carbono extraído de la atmósfera CT = Carbono total 3,667, factor basado en la razón de las masas atómicas de carbono (12) y de oxígeno (16), utilizado por el IPCC en la guía para inventarios nacionales de gases de efecto invernadero |
| Área basal (m2.ha-1) | G= π x DAP2 40.000 | G = Área basal (m2) π = 3,1416 DAP = Diámetro a la altura del pecho (1,30 m) 40.000 (se utilizó este denominador para obtener el valor de área basal en m2) |
| Volumen de fuste (m3.ha-1) (fórmula de Huber) | V= G * f (*) * hF | V = Volumen m3 G = Área basal (m2) f (*) = Factor de forma de Hutchinson (0,775) y hF= Altura de fuste (m) hT= Altura total |
| Volumen total (m3.ha-1) (fórmula de Huber) | V= G * f (*) * hT | |
| Biomasa de fuste (t.ha-1) (Brown 1997) | Bf= (V * Pe)/1000 | Bf = Biomasa de fuste (t) V = Volumen de fuste (m3) Pe = Densidad aparente de la madera (Kg/m3) |
| Biomasa total (t.ha-1) (Brown 1997) | BT= BF * FEB | BT = Biomasa total en toneladas (t) BF = Biomasa del fuste en toneladas (t) FEB = Factor de expansión de biomasa FEB = (3,42) factor que se utiliza para todas las especies de árboles |
| Carbono orgánico en el suelo (%) Método de Walkley-Black (Tedescoet al. 1995) | % CO = % MO ÷ 1,72 | % CO = Carbono orgánico % MO = Materiaorgánica |
Resultados y discusión
Composición florística. En el bosque de galería del arroyo San Lorenzo, Paraguay, se registraron 8 familias, 12 especies y 141 individuos en una superficie de 0,3 ha, equivalente a 470 individuos.ha-1. Tal como se muestra en la Tabla 5, la familia más diversa y representativa fue la Meliaceae con 3 especies, seguida por la Myrthaceae y Moraceae con 2 especies.
Tabla 5 Composición florística del bosque de galería del arroyo San Lorenzo.
| Familia | Nombre científico | Nombre común |
| CECROPIACEAE | Cecropiapachystachya | Amba'y |
| EUPHORBIACEAE | Sapiumhaematospermum | Kurupika'y |
| FABACEAE | Inga uraguensis | Inga'i |
| MELIACEAE | Trichilia catigua | Katigua |
| MELIACEAE | Cedrelaodorata | Cedro |
| MELIACEAE | Guarea glabra | Cedrillo |
| MYRTACEAE | PsidiumguajavaL. | Guayaba |
| MYRTACEAE | Syzygiumsp. | Mbopinamichai |
| MORACEAE | Macluratinctoria | Tatajyva |
| MORACEAE | Ficus enormes | Guapo'y |
| RHAMNACEAE | Hoveniadulcis | Hovenia |
| SAPOTACEAE | Chrysophyllummarginatum | Pykasurembi'u |
Valores similares a los registrados en el presente trabajo fueron reportados por Aguirre (2017) en su investigación acerca de la estimación de carbono almacenado en el bosque de galería del arroyo San Lorenzo registró 11 familias, 16 especies y individuos a 487 individuos.ha-1.
Benítez (2018), registró mayor cantidad de individuos en su trabajo de investigación referente a la dinámica de carbono en el mismo bosque de galería donde se identificaron 11 familias, 14 especies y 620 individuos.ha-1.
Especies arbóreas representativas. Como se observa en el Figura 1, las especies de mayor representatividad registrada en el bosque galería son Sapiumhaematospermum (Kurupika'y) de la familia Euphorbiaceae con 55 individuos, y Syzyguimsp.(Mbopinamichái) de la familia Myrtaceae con 40 individuos. Estas 2 especies equivalen al 67% de los individuos arbóreos registrados. Estas 2 especies fueron reportadas como las de mayor abundancia por otros investigadores en la misma zona de estudio.
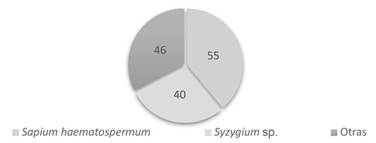
Figura 1 Especies de mayor representatividad del bosque de galería del Campus de la Universidad Nacional de Asunción, San Lorenzo.
Gómez (2013), en su investigación realizada en el bosque de galería del Campus de la Universidad Nacional de Asunción, San Lorenzo, encontró 28 individuos de la especie Sapiumhaematospermum (Kurupika'y) y 21 individuos de la especie Syzygiumsp. (Mbopinamichái). En tanto que Benítez (2018), en el mismo sitio registró 85 individuos de la especie Syzygiumsp.(Mbopinamichái) del total de 186 árboles reclutados.
Airaldi-Wood et al. (2018), en una investigación realizada en este mismo sitio para la determinación de la diversidad de quirópteros en el Campus, mencionaron que las especies Sapiumhaematospermumy Syzygiumsp.,del estrato alto del bosque se encuentran asociadas en el sitio junto con otras especies vegetales que proporcionaron a partir de sus frutos, el alimento para murciélagos frugívoros.
Volumen de fuste. El volumen de fuste de los árboles del bosque de galería en estudio fue de 48,5 m3.ha-1, con un rango de 29,3 a 70,4 m3.ha-1 por parcela.
Como se observa en la Figura 2, existe una diferencia significativa en descenso si se tomara en cuenta las citadas investigaciones de años anteriores. El descenso, presentado se debe al bajo número de individuos.ha-1 encontrado con respecto a los trabajos anteriores.

A esto se le suma la gran cantidad de individuos enfermos encontrados. Del total de 141 individuos arboreos en 0,3 hectáreas del bosque de galería del Campus, 91 estaban enfermos lo que equivale a un 64,50% y 50 se encontraban sanos lo que correspondió a 35,40%.
Propiedades físicas y químicas del suelo. El suelo del bosque de galería estudiado presentó textura arenosa, con tonos que van de marrón a marrón pálido. La densidad del suelo fue de 1287,5 kg/m3 y 1375 kg/m3 para las 2 profundidades, lo 1u3 según Fuentes Yague (1999) corresponde a suelos no compactados.
Aldana Lázaro (2017), sostuvo que el suelo de la especie Schinus molle contenía carbono en el rango de 0,64% a 0,72% y el suelo de la especie Ficus benjamina contiene carbono en un rango de 0,20% a 0,27%. Siendo el suelo de la especie Schinus molle la que almacenaba mayor cantidad de CO2 con un total de 43,20 t.ha-1, mientras que el suelo de la especie Ficus benjamina almacena 20,92 t.ha-1. Esto se debe a que existen mayor cantidad de individuos forestales pertenecientes a la especieSchinus molle en comparación a la especie Ficus benjamina.
Estos valores son próximos a los reportados por Andrade-Castañeda et al. (2016) quienes obtuvieron una densidad aparente ente 1,1 y 1,7 g/cm3.
El nitrógeno total encontrado en el suelo fue de 0,09% y 0,06% para 0 - 10 cm y 10 - 30 cmrespectivamente. La materia orgánica se encontraba en niveles bajos y medios, y el pH ácido. En cuanto a los nutrientes, el nivel de Fósforo (P) fue bajo, el contenido de Calcio (Ca+2) fue medio, el contenido de Magnesio (Mg+2) y Potasio (K+) fue alto, y por último el porcentaje de aluminio (Al+3+H+) fue bajo Tabla 6.
Tabla 6 Propiedades químicas del suelo del bosque de galería.
| Profundidad (cm) | PH | P (mg.kg-1) | Ca+2 (mg.k-1) | Mg+2 (mg.kg-1) | K+cmolc.Kg-1 | Al+3+H+ (mg.kg-1) |
| 0 - 10 | 5,28 | 2,18 | 3,08 | 1,14 | 0,34 | 0,21 |
| 10 - 30 | 5,38 | 2,18 | 3,03 | 1,33 | 0,40 | 0,22 |
Los datos coincidieron con lo mencionado por Aguirre (2017), quien en su investigación señaló que el suelo corresponde a la clase textural arenosa con tonalidad marrón, amarillo pálido a marrón negruzco entre otras. En cuanto al Nitrógeno (N) el porcentaje ha disminuido si se compara con los resultados obtenidos por Aguirre, quien obtuvo valores de 0,15% y 0,09% respectivamente.
Según Jandlet al. (2004), la fuente principal del Nitrógeno (N) en los suelos la constituye la mineralización de la materia orgánica. Según los resultados obtenidos a mayor profundidad disminuye la MO, y por tanto repercute en la relación Carbono/Nitrógeno (C/N).
Velocidad de infiltración media en el suelo. En el primer ensayo se obtuvo un promedio de 102 mm.h-1. El segundo ensayo se realizó a una distancia de 100 metros del primer ensayo, terreno abajo cercano al cauce, con un resultado de 30 mm.h-1. Se estima que la velocidad de infiltración media en el suelo del bosque de galería fue de 66 mm.h-1 según esos 2 resultados Figura 3.
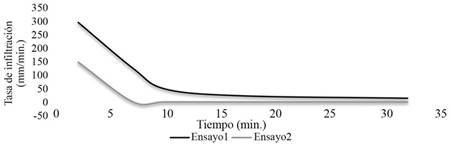
Los macroporos del suelo están colmados de agua sin posibilidad de circulación del aire. La casi nula presencia de oxígeno (O2) en los poros de los horizontes superficiales, permite las condiciones anaeróbicas donde existe una descomposición muy lenta de la materia orgánica. Esta condición afecta la disponibilidad de los nutrientes para las plantas y su desarrollo.
Thiers et al. (2014), se refirieron a esta condición de saturación al citar que el agua compite con el aire del suelo para ocupar el espacio poroso y, por lo tanto, controla la convección de gas y difusión a través del suelo, que afecta la actividad de los organismos y el potencial de oxidación/reducción en el suelo.
Van Lier (2017), afirmó que el valor de la conductividad hidráulica saturada, a su vez está determinada en gran medida por la presencia de macroporos y su continuidad. Por lo tanto, una pérdida de macroporosidad, como ocurre en la degradación del suelo por compactación, resulta en una conductividad hidráulica reducida saturada y por lo tanto con un aumento en el contenido de agua a capacidad de campo.
Carbono acumulado y CO2 equivalente en el bosque de galería. El carbono almacenado en el estrato arbóreo fue de 38,4 tC.ha-1, el error de muestreo ±1,64 tC.ha-1 con un nivel de confianza de 95%, el valor promedio de carbono acumulado en el suelo fue de 31,08 tC tC.ha-1,que totaliza 69,48 tC.ha-1de carbono acumulado en los 2 componentes del ecosistema en estudio.
Los resultados de Andrade-Castañeda et al. (2016) mencionan que el carbono almacenado en los bosques riparios de su área de estudio fue de entre 50,2 a 87,1 tC.ha-1, resultados muy altos en comparación con lo reportado por Batjes (1999) con los datos de FAO-IIASA en zonas agroecológicas y climáticas similares del trópico (36 a 38 tC.ha-1).
El contenido de carbono del mismo bosque según Benítez (2018), fue de 126,9 tC.ha-1. En la presente investigación se obtuvo un total de 69,48 tC.ha-1 lo que expone una diferencia de 57,41 tC.ha-1. Al considerar esa variación, se demuestra que hubo un descenso en la cantidad de carbono acumulado en el periodo (2018-2020) que podría atribuirse a la mortandad de árboles en la zonade estudios, al respecto, Lugo y Scatena (1996) sugieren que en la mayoría de los árboles de clases diamétricas y alturas superiores ocurre una senescencia producida por factores endógenos como la genética y la edad.
Así también Carey et al. (1994) afirman que los árboles mueren de pie producto de debilitamiento a causa de enfermedades o estrés ambiental, y en su caída llevan a otros individuos de clases diamétricas menores.
El dióxido de carbono equivalente extraído de la atmósfera arrojó un promedio de 254,81 tCO2e.ha-1,valor que ha disminuido según las investigaciones anteriores, y que también fue atribuible a la muerte de individuos arbóreos,tal como se puedever en la Figura 4.

Estos valores son superiores a los reportados por Suarez García (2017) quien determinó el almacenamiento de biomasa 9,46 t.ha-1 y carbono 4,73 t.ha-1 CO2,así como el potencial de captura de 30,10 t.ha-1 CO2.
Resultados similares fueron reportados por González Pantoja (2019) en donde el Cerro Nutibara almacenó 34 tC.ha-1 y 126 tC.ha-1 mientras que el Cerro Volador 6 tC.año-1 y 20 126 t CO2 e.
Los resultados de la presente investigación coincidieron con las afirmaciones de Torres (2017), quien indicó que el carbono almacenado en la biomasa aérea fue mayor que la del suelo.
Conclusiones
El bosque de galería ofrece servicios ecosistémicos de regulación como captación de carbono, constituyen la biomasa arbórea el mayor reservorio de carbono pese a la mortandad muchos individuos arbóreos lo que provoca la reducción en su capacidad de almacenamiento de carbono, aunque representan un gran potencial para la captura de CO2. Son una fuente de aprovisionamiento de agua para la conservación, además de constituirse como fuente de nutrientes del suelo que a excepción del fósforo, se mantuvieron en niveles medios y altos, en la sub cuenca del arroyo San Lorenzo, Paraguay.

