











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La pitahaya (Hylocereus spp.) es un cactus epífito con una creciente importancia comercial a nivel internacional, debido a su potencial como planta ornamental y para el consumo de tallos y frutos. La industrialización del cultivo ha favore- cido la creación de empleos, el aprovechamiento de áreas degradadas y un aumento en los ingresos de pequeños productores (Ortiz y Carrillo 2012, Valencia et al. 2013).
En las últimas décadas el interés por este cultivo se ha incrementado, ya que el fruto con- tiene pigmentos nitrogenados solubles en agua, llamados betalaínas. Estas presentan potencial como colorantes naturales, debido a que su ámbi- to de color que va del rojo-violeta al amarillo, se mantienen estables en un ámbito de pH más amplio (3-7) en comparación con otros colorantes naturales como las antocianinas (Stintzing et al. 2001, Stintzing et al. 2002, Strack et al. 2003, Wybraniec et al. 2001).
En relación con la expansión de la pita- haya en Costa Rica, esta ha tenido un incre- mento en los últimos años, sin embargo, aún se considera un cultivo minoritario, por lo que no se han realizado estudios detallados sobre las enfermedades que puede presentar. A nivel mundial se reporta que este cactus puede ser atacado en tallos y frutos por las bacterias Enterobacter cloacae, Erwinia carotovora y Xanthomonas campestris, y algunos hongos como Fusarium oxysporum, Fusicoccum sp., Curvularia lunata, Colletotrichum gloespo- rioides y Neoscytalidium dimidiatum (Valen- cia et al. 2004, Wright et al. 2007, Palmateer y Ploetz 2007, Masyahit et al. 2009, Masratul et al. 2013).
En plantaciones comerciales de este cactus en México, se reportó la presencia en los tallos de una pudrición suave. Se identificó una ente- robacteria como agente causal de dicho síntoma (Valencia et al. 2003).
Debido a la heterogeneidad de las especies del género Enterobacter y a la cercanía que pre- sentan en las secuencias de la región 16S, es com- pleja la identificación de las mismas. La mayoría de las especies que muestran una relación gené- tica por encima de un 60%, con E. cloacae, son agrupadas en el complejo Enterobacter cloacae (Hoffmann y Roggenkamp 2003).
Con base en lo anterior y ante la problemá- tica fitosanitaria que se presenta en este cultivo en el país, se planteó como objetivo de este trabajo el aislamiento, identificación y la verificación de los postulados de Koch de la bacteria que causan las pudriciones que afectan los tallos de la pitahaya en Costa Rica.
MATERIALES Y MÉTODOS
Recolección de muestras
La investigación se realizó en la finca “El Brillante”, la cual se dedica a la producción de pitahaya orgánica y se ubica en Miramar de Pun- tarenas (10° latitud norte y 84° longitud oeste), Costa Rica. La población en el área de estudio fue de 2666 plantas.ha-1, de las cuales se seleccionó 992 plantas distribuidas en 38 hileras para reali- zar esta investigación.
En la plantación se presentaron 2 tipos de pudrición con sintomatologías distintas, una se presentó en época seca y la otra en el perio- do lluvioso. De cada una estas se tomaron 25 muestras, cladodios completos con diferentes estados de desarrollo de la enfermedad; cada una se envolvió en papel periódico humedecido y fue colocada en una bolsa plástica. El material recolectado fue llevado al Laboratorio de Fito- patología del Centro de Investigación en Protec- ción de Cultivos (CIPROC) de la Universidad de Costa Rica (UCR).
Se evaluó la evolución de los síntomas pre- sentes en el campo durante un año. Para cada tipo de pudrición se marcaron 30 plantas a lo largo de la plantación con diferentes estados de desarrollo de los síntomas y se procedió a evaluar cambios visibles mediante fotografías generadas cada 15 días durante el periodo antes mencionado.
Caracterización de síntomas y aislamiento del agente causal
Para cada tipo de pudrición, se realizó la descripción de los síntomas presentes en las 25 muestras recolectadas, con base en las carac- terísticas visuales que cada una presentó, con ayuda de un estereoscopio. Los aspectos que se evaluaron fueron el color, tamaño y textura de lesiones, además, la consistencia y la tonalidad del tejido interno.
Se seleccionaron 15 cladodios sintomáti- cos de cada enfermedad, a partir de los cuales se llevó a cabo el aislamiento del agente causal mediante la metodología descrita por French y Hebert (1980) para hongos y bacterias. La desin- fección del material se realizó con etanol al 70% e hipoclorito de sodio al 1% durante 50 s y 20 s, respectivamente.
Los medios de cultivo utilizados fueron MacConkey y Agar-Triptona-Soya (ATS) (Atlas 2010). En cada uno de los síntomas, se rasgó el tejido de manera longitudinal y se extrajeron segmentos de la zona de avance en la parte interna del mismo (parte enferma y parte sana). Se triplicó cada aislamiento por cada medio utilizado, para un total de 45 repeticiones, por cada tipo de pudrición. Estos se incubaron en una cámara oscura a temperatura de 30°C, por un periodo de 3 días.
Una vez transcurrido este tiempo, se obtu- vo una colonia bacteriana por cada tipo de pudri- ción, visualmente distintas, que predominaron en todos los aislamientos realizados. Con ambas bacterias, se procedió a realizar la separación, con un asa desinfectada, de colonias individuales en medio ATS. Dichas colonias se incubaron bajo las mismas condiciones de temperatura y oscuri- dad, antes descritas, por un periodo de 24 horas. A partir de una colonia individual de cada bacteria, se realizó la purificación y multipli- cación de estas en medio ATS. Se contó con 3 repeticiones por bacteria y se incubaron a 30ºC
por 24 horas.
Con el material purificado, se procedió a realizar las siguientes pruebas bioquímicas y moleculares para la identificación:
Tinción de Gram
Se realizó la prueba de tinción de Gram con el objetivo de confirmar y determinar la morfología de las bacterias. Lo anterior se llevó a cabo mediante la metodología descrita por Schaad et al. (2001). Se observó la morfología de estas en el microscopio de luz, con un aumento de 100X
Prueba Triple Azúcar-Hierro (TSI)
Cada bacteria se inoculó por duplicado con aguja bacteriológica estéril en tubos de ensayo que contenían aproximadamente 10 ml de medio TSI y se incubaron por 24 horas a 30°C. Transcu- rrido este periodo, se evaluó el perfil de fermen- tación que presentaban, según el cambio de color por el indicador de pH (Rodríguez et al. 2005).
Prueba de las enzimas oxidasa y catalasa
Se determinó la presencia de la enzima citocromo C oxidasa en las bacterias en estudio. A partir de una suspensión de cada una de estas, se tomó una gota y se colocó en papel filtro que contenía el reactivo tetrametil-p-fenilendiamina. Se observó la velocidad con que ocurrió el viraje. En el caso de la prueba de la enzima catalasa, se colocó una gota de cada suspensión bac- teriana en un portaobjetos y se le agregó peróxido de hidrógeno 15% (H2O2). Se evaluó la liberación de oxígeno mediante la formación de burbujas (Rodríguez et al. 2005).
Identificación molecular
Extracción del ácido desoxirribonucleico (ADN)
Se realizó la identificación molecular en el Laboratorio de Técnicas Moleculares Aplicadas a la Fitoprotección del CIPROC, UCR, para lo cual se extrajo ADN a partir de 2 repeticiones de cada bacteria purificada, con 24 horas de crecimiento, mediante el uso del método CTAB (Murray y Thompson 1985). El ADN fue cuantificado con un espectofotómetro y se llevó a una concentra- ción final de 80 ng.μL-1
Reacción en Cadena de la Polimerasa (PCR) y secuenciación del ADN
La reacción en cadena de la poli- merasa (PCR) se realizó con los cebadores específicos: 16S, 27F (5'- AGAGTTTGATCMT- GGCTCAG-3') y 1492R (5'-TACGGYTACCTT- GTTACGACTT-3') descritos por Lane et al. (1985), que amplifican la secuencia total del ARN del gen ribosomal 16S.
Para cada repetición, la reacción se llevó a cabo en un volumen final de 25 μL, que consistió en 15,25 μL de agua desionizada ultrapura, 2,5 μL de buffer 10X (1X), 2,5 μL de dNTPs (1 mM), 1,25 μL de cada cebador (0,5 μM) y 0,25 μL de DreamTaq DNA Polymerase (1 U.μL-1). Se utili- zó un control negativo, el cual fue una reacción sin ADN. La reacción de amplificación se llevó a cabo mediante el siguiente perfil térmico: predes- naturalización inicial a 96ºC durante 2 minutos, seguido por 35 ciclos de desnaturalización a 96ºC durante un minuto, anillamiento a 56ºC por un minuto, extensión a 72ºC por 2 minutos, seguidos de una extensión final a 72ºC durante 10 minutos. El producto de PCR (aproximadamente de 1500 pares de bases), de cada repetición por bac- teria, para 16S se confirmó mediante electrofore- sis en gel de agarosa (0,8%), que se comparó con un marcador de peso molecular de 100 pb, Gene- Ruler. El producto de PCR se digirió con una endonucleasa llamada Exonuclease I y se llevó a una concentración de 50 ng.μL-1. Posteriormente, los 4 productos de PCR obtenidos se enviaron a secuenciar a la empresa Macrogen Inc., Corea del Sur. Se obtuvieron secuencias en ambas direccio- nes para cada muestra generada a partir de ciclos fluorescentes con un secuenciador.
Alineamiento de las secuencias y análisis filogenéticos
La calidad de las secuencias se confirmó con un alineamiento bidireccional y por compa- ración con los cromatogramas con el programa BioEdit (Hall 1999); además, se cortaron las terminaciones 5’ y 3’ de las cadenas para faci- litar el alineamiento. Se utilizó la herramienta informática Blast (Basic Local Alignment Search Tool del National Center for Biotechnology Infor- mation (NCBI), a partir de la cual se comparó la secuencia generada frente a las secuencias que se encuentran en la base de datos del banco de genes (GenBank).
El análisis filogenético se realizó con el software MEGA 6. Se llevaron a cabo las pruebas del Vecino más cercano y Máxima Verosimilidad, con 1500 repeticiones y accesiones de Enterobac- ter spp., provenientes del Genbank.
Prueba Vitek 2
Como respaldo, se llevó a cabo la iden- tificación de las bacterias mediante el sistema automatizado de identificación Vitek 2, en el Laboratorio de Bacteriología Médica de la Facul- tad de Microbiología de la UCR. Se realizaron 2 repeticiones por bacteria. El sistema Vitek 2 es una prueba bioquímica que utiliza una tarjeta con 64 pozos, cada uno de los cuales contiene un sustrato diferente con los que se miden las acti- vidades bioquímicas de las bacterias, tales como acidificación, alcalinización, hidrólisis enzimá- tica y crecimiento en presencia de sustancias inhibitorias (Pincus 2005).
Identificación de organismos secundarios
En época seca, sobre algunas de las lesio- nes viejas provocadas por la bacteria, se presen- taron estructuras redondeadas y oscurecidas, por lo que se realizó el aislamiento de tejido con estos signos en cajas Petri con medios de cultivo agar-papa-dextrosa (PDA) y V8 (Atlas 2010). Se contó con 2 repeticiones por medio de cultivo y se colocaron en oscuridad y a 25-30°C por 5 días. Se observaron los conidios de los organismos obtenidos, en el microscopio de luz, con un aumento de 100X. Asimismo, se visualizaron las estructuras oscurecidas con el estereoscopio.
Verificación de los postulados de Koch
A partir de cada una de las bacterias purificadas, se preparó una suspensión de apro- ximadamente 1,0x108 unidades formadoras de colonias por mililitro (UFC.ml-1) y se inocularon en tallos sanos de pitahaya. Para esto se tomaron 20 ml de agua estéril y se les agregó pequeñas cantidades de bacteria hasta lograr una turbidez similar a un tubo de McFarland con la concentra- ción mencionada.
La inoculación se llevó a cabo en el invernadero del Laboratorio de Fitopatología de la Universidad de Costa Rica en época seca (Febrero, 2014). En cladodios sanos y previamen- te desinfectados con etanol al 70% se inyectó las suspensiones preparadas. Para cada bacteria se inocularon 40 puntos distribuidos en 4 cladodios a los cuales se les colocó 0,5 ml de suspensión por cada punto de infección. Se contó con un testigo en cada cladodio que consistió en inyectar en la cara opuesta de los puntos de infección agua estéril con el mismo volumen y con la cantidad de puntos antes mencionados.
Se confirmaron los postulados de Koch gracias al reaislamiento de las bacterias inocula- das mediante el protocolo descrito por French y Hebert (1980) en los medios de cultivo antes men- cionados, con 3 repeticiones. La identificación del organismo obtenido se realizó por medio de las mismas pruebas bioquímicas implementadas anteriormente.
RESULTADOS Y DISCUSIÓN
En la plantación se presentaron síntomas diversos asociados al patógeno en estudio. Con- forme la enfermedad evolucionó en el tiempo, se dieron cambios en la coloración y morfología de las lesiones provocadas por la bacteria.
De acuerdo con las evaluaciones realiza- das, en el área de estudio se presentaron 2 tipos de pudriciones, una durante la época lluviosa y otra durante la época seca.
Época lluviosa
Durante la época lluviosa se presentan pudriciones amarillentas en diferentes estados de desarrollo que principalmente inician de los bordes y se extienden hacia el centro del clado- dio (Figura 1A, B y C). En algunos casos, las infecciones inician del centro del tallo a partir de lesiones provocadas por otros patógenos, como N. dimidiatum, o daños por insectos.

Fig. 1 Síntomas de pudrición presentes en los cladodios de pitahaya durante la época lluviosa. A. Síntoma inicial de la enfermedad. B y C. Avance de la pudrición en el tejido. D. Síntoma final de la pudrición.
En estados avanzados de la enfermedad se presentan 2 situaciones: la pudrición se limita a segmentos del cladodio o se da la pudrición por completo del mismo. En el primer caso el daño es detenido por la planta, debido a que aparente- mente esta tiene una alta respuesta hipersensible, la cual se puede generar por los altos contenidos de calcio que la planta es capaz de almacenar en forma de oxalato, tanto en los idioblastos como fuera de estos (Franceschi y Horner 1980, Webb 1999, Faheed et al. 2012). En el segundo caso, el tejido se rompe por el peso del material descompuesto, el cual al caer al suelo deja la epi- dermis unida al resto de la planta. Esta al secarse presenta una textura suave y papelosa de color blanquecino, lo que facilita su desprendimiento
y deja expuesto el haz vascular del tallo afectado (Figura 1D).
Un síntoma similar, provocado por Erwi- nia carotovora, se reportó en tallos de Hyloce- reus sp. Según OIRSA (2000), este patógeno se presenta en plantaciones comerciales de pitaha- ya en Nicaragua y provoca manchas amarillas que posteriormente avanzan como pudriciones acuosas y fétidas que destruyen por completo los cladodios. Cabe señalar que no se menciona la metodología utilizada para la identificación de este patógeno.
Época seca
Al final de la época lluviosa se presentó una serie de pudriciones de color marrón-negro.
Estas inician como puntos rojizos sin halo que posteriormente crecen y se tornan de color marrón. Los síntomas se desarrollan tanto en los bordes como en el centro del cladodio, con un crecimiento irregular que se extiende por todo el tallo (Figura 2A y 2B).

Fig. 2 Síntomas de pudrición presentes en los tallos de pitahaya durante la época seca. A. Síntoma inicial de la enferme- dad. B. Avance de la pudrición en el tejido. C. Síntoma final cuando la enfermedad afecta un segmento del tallo. D. Síntoma final cuando la enfermedad afecta todo el cladodio.
Los estados avanzados de la pudrición se presentaron de 2 maneras: si el daño se restringe a un segmento del cladodio, el tejido se contrae y adquiere una coloración blanquecina-grisácea con un halo ligeramente amarillento alrededor de la lesión (Figura 2C). En el caso de que la pudrición afecte completamente el cladodio, el tejido se con- trae hacia el haz vascular con una textura acartona- da y endurecida de color blanquecino (Figura 2D).
En algunos casos, se da la colonización de estructuras subglobosas de color negro sobre lesiones secas provocadas por la bacteria (Figura 3A). Estas corresponde a acérvulos de Colleto- trichum spp. (Figura 3B). También se pueden presentar conidios de Fusarium spp. (Figura 3C). En ambos casos, estos hongos actúan como organismos saprófitos del tejido afectado por E. hormaechei.

Fig. 3. Organismos saprófitos presentes en lesiones viejas provocadas por E. hormaechei, bajo condiciones de época seca. A. Acérvulos de Colletotrichum spp. B. Conidios de Colletotrichum spp. C. Conidios de Fusarium spp.
Los síntomas descritos concuerdan con lo reportado por Masyahit et al. (2009), que mencio- nan a Enterobacter cloacae como agente causal de las pudriciones acuosas en plantas de pitahaya en Malasia, las cuales inician como un ablandamiento del tejido, se tornan de color marrón y finalizan con la caída del tejido, sin que se afecte el haz vascular.
De acuerdo con Valencia et al. (2004), en plantaciones comerciales de fruta dragón en México, se desarrolla una pudrición blanda que algunas veces presenta un halo clorótico, y men- cionan una enterobacteria (sin identificar) asocia- da a estos síntomas
Aislamiento del agente causal
De cada tipo de pudrición se obtuvo una bacteria de color amarillento en medio de cultivo ATS y de color morado en MacConkey. Con base en lo anterior, se determinó que estas bacterias presentan la capacidad de fermentar lactosa
Pruebas Gram, Triple Azúcar-Hierro (TSI) y de las enzimas oxidasa y catalasa
Según los resultados de la tinción de Gram y de las pruebas bioquímicas, ambas bacterias son bacilos (con forma de bastón) Gram negativos, catalasa positivas y oxidasa negativas. Con la prue- ba con TSI se comprobó que las bacterias fermen- tan glucosa y sacarosa (A/A). Asimismo, debido a que hubo producción de gas, también presentan la capacidad de fermentar carbohidratos (G).
Lo anterior concuerda con las caracterís- ticas que presentan las bacterias que pertene- cen a la familia Enterobacteriaceae y al género Enterobacter. Estos agrupan microorganismos con forma de bastón (no pigmentados), aerobios, móviles capaces de fermentar glucosa a ácido con producción de gas o sin ella, son oxidasa nega- tivas y catalasa positivas (Brenner et al. 1986, Puerta y Mateos 2010)
Prueba Vitek 2
Según los resultados del Vitek 2, la especie de la bacteria corresponde a Enterobacter cloa- cae spp. cloacae, bionúmero 2627634553533010. D cloacae y E. hormachechei presentan un 63% de cercanía filogenética y difieren en que esta última muestra resultados negativos en las prue- bas con D-sorbitol, melibiosa, esculina, D-rafino- sa, α-metil-D-galactosida y adonitol. Se reportan cepas de E. hormachechei que dan positivo en la pruebas con D-sorbitol y melibiosa, lo que sugiere la existencia de biotipos de esta especie (O'Hara et al. 1989, Davin et al. 1997). Lo ante- rior podría explicar la diferencia obtenida en la identificación de la especie entre esta prueba y el análisis molecular
Identificación molecular
Con base en los resultados del Blast, 3 de las 4 secuencias obtenidas corresponden a un mismo organismo. De acuerdo con la región 16S y la comparación de complementariedad de secuencias de bases según el GenBank, con un total de 1352 nucleótidos analizados, esta bacteria presenta un 99,23% de homología con Enterobac- ter hormaechei (número de accesión JF690889.1) y 99,08% con E. asburiae (número de accesión JQ682630.1). De las secuencias provenientes de los aislamientos analizados, se introdujo una en el Genbank bajo el número de accesión KJ999997.
Cabe señalar que las 3 secuencias pro- venientes de H. costaricensis se unen en único clado, a pesar de algunas mutaciones que las diferencian entre los nucleótidos 373 al 395. Al comparar con otras secuencias provenientes del GenBank, las obtenidas en esta investigación presentan mayor cercanía con E. hormaechei y E. asburiae, y tienen cierta lejanía con la especie E. cloacae (Figura 4).
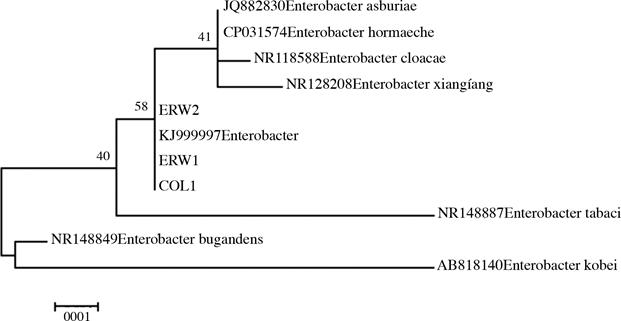
Fig. 4. Relación filogenética entre las secuencias de bacterias obtenidas de H. costaricensis con otras especies cercanas del género Enterobacter.
A pesar de lo anterior, E. hormaechei pertenece al complejo “Enterobacter cloacae”, en el cual se incluyen 6 especies: Enterobacter asburiae, Enterobacter cloacae, Enterobacter hormaechei, Enterobacter kobei, Enterobacter ludwigii y Enterobacter nimipressuralis. Estas pertenecen a linajes distintos dentro de un grupo genético común, debido a que presentan relacio- nes entre sus ADN muy estrechas, es decir, que existen regiones conservadas que se comparten a nivel de género e incluso entre diferentes especies. Fenotípicamente se pueden distinguir mediante pruebas bioquímicas, tales como Bio- log, Vitek, TSI, entre otros (O'Hara et al. 1989, Brenner et al. 1986, Hoffmann et al. 2005).
Las bacterias mencionadas comparten regiones conservadas en el genoma que las hace cercanas entre sí, pero presentan polimorfismos que permiten diferenciarlas. E. hormaechei se diferencia de otras enterobacterias porque pre- senta una fuerte actividad de la enzima glucosa deshidrogenasa. Esta oxida D-glucosa a D-glu- conato sin la presencia de pirroquinolina quinona (Grimont y Grimont 2006).
Cabe mencionar que la identificación molecular de la bacteria se llevó a cabo mediante el estudio del ARNr 16S, porque son fragmentos del genoma altamente repetidos que se han man- tenido bastante uniformes a lo largo de la evolu- ción. Además, proporcionan un sitio de iniciación adecuado para la elongación de los imprimadores y así, de forma más fácil, realizar la secuenciación (Lane et al. 1985, Neefs et al. 1993).
Las secuencias de ADN ribosómico (ADNr) de E. cloacae presentan variaciones en la región 16S que no permiten formar un grupo taxonómico homogéneo, pero a partir de estas se puede formar un árbol filogenético irregular en el que las cepas de Enterobacter cloacae se interre- lacionan fuertemente con otras especies de este género, como E. aerogenes y otras enterobacte- rias como E. coli. Esto refleja la heterogenicidad genética de las especies, lo que dificulta su identi- ficación sistemática (Tang et al. 1998, Hoffmann y Roggenkamp 2003).
Verificación de los postulados de Koch
Se obtuvo con un 100% de infección en cada uno de los cladodios inoculados con la bacteria obtenida (Enterobacter hormaechei). Esta al ser inoculada en tallos sanos, generó los mismos síntomas que se observaron durante la época seca en la plantación. En el testigo se dio la cicatrización de las heridas provocadas con la aguja a partir del octavo día después de iniciadas las inoculaciones, sin que se presen- taran síntomas.
Ocho días después de la inoculación (ddi) se observaron manchas amarillentas en los puntos donde se inyectó el patógeno (Figura 5A). La coloración de estas se intensificó y el centro de las mismas se tornó rojizo-marrón a los 12 ddi (Figura 5B). Ocho días después, el tejido infectado se hundió levemente y adquirió una coloración anaranjada con el borde rojizo (Figura 5C).

Fig. 5.Resultados de la inoculación de E. hormaechei en tallos sanos. A. Día 8. B. Día 12. C. Día 20. D. Día 24. E. Día 52; después de inoculación.
A los 24 ddi se dio la pudrición por com- pleto del tejido inoculado (Figura 5D). La fase final de los síntomas se manifestó a los 52 ddi como lesiones blanquecinas con un borde defi- nido color crema y con puntos necrosados en el centro de las mismas (conidios de Colletotrichum spp.) (Figura 5E).
Paralelo a lo anterior, en la cara inmedia- ta a los puntos de infección, se desarrolló una sintomatología similar a la mencionada, pero con mayor vigor. A los 8 días después de la inoculación se presentaron manchas irregulares color marrón (Figura 6A). Posteriormente, a las 20 ddi la pudrición tomó una tonalidad marrón- negro con un hundimiento en el mismo. El avance del oscurecimiento de la lesión ocurrió del centro hacia los bordes del síntoma descrito (Figura 6B).
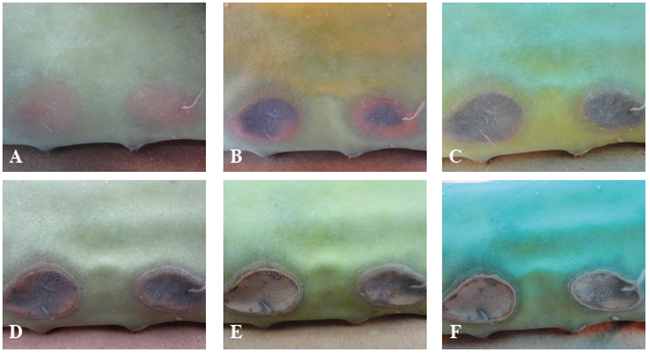
Fig. 6. Síntomas provocados por E. hormaechei inoculada en tallos sanos, en la cara opuesta de los puntos de inoculación. Día 8B. Día 20. C. Día 24. D. Día 36. E. Día 44. F. Día 52; después de la inoculación.
A los 24 ddi, se dio la pudrición por com- pleto del tejido infectado (Figura 6C). Doce días después, se presentó un aumento del hundimiento en las lesiones y el inicio de un cambio de color de marrón oscuro a marrón claro (Figura 6D), el cual se finalizó a los 44 ddi (Figura 6E).
Aproximadamente, 7 semanas después, se obtuvo como síntoma final lesiones blanquecinas irregulares blanquecinas con halo clorótico, sin la presencia de estructuras de organismos secun- darios (Figura 6F). De los reaislamientos de los síntomas descritos se obtuvo la misma bacteria inoculada.
Esta bacteria se reporta como un orga- nismo benéfico para las plantas, asociado al sistema radical de varios cultivos y relacionado con la promoción del crecimiento. A pesar de lo anterior y con base en observaciones de campo, se presume que la bacteria, además de aprovechar heridas para ingresar en los tejidos, puede ser endófita (se reporta como tal en otras especies vegetales) y se comporta como un organismo fitopatógeno cuando la planta se encuentra bajo condiciones de estrés (Kampfer et al. 2005, Ogbo y Okonkwo 2012, Ying et al. 2013, Egamberdieva et al. 2014).
Durante el desarrollo del estudio, se obser- vó que en la época seca las plantas presentaron síntomas de estrés lumínico y a la vez un incre- mento de la incidencia y la severidad de la enfer- medad causada por la bacteria. Esta situación se puede deber a que la pitahaya requiere de 30% a 60% de sombra, por lo que un un flujo de fotones para la fotosíntesis (FFF) mayor a 20 mol m-2 d-1 puede provocar fotoinhibición en especies como H. undatus. Lo anterior puede ocurrir en otras especies a diferentes FFF cercanos al valor mencionado, lo que se traduce en una menor tasa fotosintética y altera los recursos disponibles para la defensa de la planta (Raveh et al. 1998, Nobel y De la Barrera 2004).
Cabe señalar que la situación mencionada puede estar implícita en las diferencias de la sintomatología que se presenta en la época llu- viosa y seca. Es decir, cambios en las condiciones ambientales y en la capacidad de respuesta de la planta pueden generar modificaciones en los síntomas que este patógeno provoca (Arauz 2011, Retana 2015).
El primer reporte científico de pudriciones provocadas por una enterobacteria en tallos de pitahaya fue realizado por Masyahit y colabo- radores en el 2009, en plantaciones comercia- les de esta fruta en Malasia. Este es el primer reporte que se hace en el continente americano de E. hormaechei como patógeno de Hylocereus costaricensis.

