











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La ganadería bovina en Costa Rica inicia su desarrollo a partir de 1561, cuando ingresaron los primeros bovinos de origen europeo. En 1568, el gobernador de Costa Rica Perafán de Rivera ordenó la importación de 400 bovinos de Honduras y de ahí se fundaron los primeros hatos de cría en el país (Quirós 2006). En los siglos posteriores se dio la evolución de diferentes sistemas de producción en distintas regiones del país y el ingreso de diversos tipos raciales adecuados para diferentes fines: leche carne o doble propósito. Según el último censo ganadero se registró en Costa Rica una población bovina de 1 358 209 cabezas de las cuales un 70,3%, (955 854 cabezas) eran hembras y un 29,6% (402 355 cabezas) eran machos (CORFOGA 2000).
Los estudios de genética poblacional ayudan a cuantificar la diversidad genética presente en una población. Las relaciones genéticas entre diferentes subpoblaciones se pueden inferir a partir de los datos de las frecuencias de los alelos. Recientemente, los datos del ADN de microsatélites son ampliamente utilizados para los estudios de genética de poblaciones en diversos organismos, debido a su alto grado de polimorfismo genético (Estoup et ál. 2002).
Para comparar la diferenciación existente entre subpoblaciones raciales se pueden utilizar índices de distancia genética tales como los estadísticos F (Wright 1965, Nei 1977, Weir y Cockerham 1984), que permiten establecer comparaciones, determinar relaciones filogenéticas y analizar la estructura de la población. Los estadísticos FIS y F IT explican las correlaciones entre 2 unidades gaméticas tomadas al azar de una subpoblación y del total de la población como un todo respectivamente; y nos indica el exceso o déficit de heterocigotos presentes; mientras que el FST es la correlación entre 2 gametos tomados al azar de cada una de las subpoblaciones y mide el grado de diferenciación genética entre 2 subpoblaciones (Wright 1965).
Más recientemente Slatkin (1995) desarrolló la medida RST que se define como la proporción de variación en la longitud de alelos de loci de microsatélites debido a las diferencias entre poblaciones. El RST asume que cada mutación cambia una longitud de un alelo por una unidad repetida (Slatkin 1995).
En Costa Rica no se ha realizado aún ningún análisis de diversidad genética dentro de la población bovina existente. El objetivo del presente estudio es analizar la diversidad genética en subpoblaciones raciales de la población bovina de Costa Rica, por medio de marcadores microsatélites.
Materiales y métodos
Procedencia y estructura de la muestra
Para la realización del presente estudio se contó con 1412 muestras de ADN bovino obtenidas en 744 hatos localizados en distintas zonas ganaderas del país (Cordero et ál. 2013). Las muestras disponibles correspondieron a ADN extraído a partir de folículos pilosos de la cola del bovino. Con base en las recomendaciones de FAO/ISAG (FAO 2011) se procuró maximizar el número de hatos visitados para evitar muestrear animales emparentados, por lo que se limitó a 2 el número de muestras colectadas por establecimiento, salvo aquellos en los cuales se encontraron múltiples tipos raciales.
Las muestras se colocaron en sobres individuales con el número de registro oficial del establecimiento y el sistema de producción según el Sistema Integrado de Registro de Establecimientos Agropecuarios (SIREA 2013), y la identificación del animal dentro de la finca, tipo racial del animal, sexo y edad. El tipo racial se estableció de manera aproximada con base en apreciaciones visuales. Debido a la alta variabilidad racial encontrada, se realizó una clasificación por tipos raciales afines (Cuadro 1), de lo cual se obtuvo finalmente un total de 16 grupos. El razonamiento utilizado para realizar esta agrupación detalla a continuación.
Cuadro 1: Número de animales muestreados por raza.
| Subpoblación | Tipo Racial | Nomenclatura | Número de animales | % |
|---|---|---|---|---|
| 1 | Bos indicus | B_ind | 97 | 6 , 9 |
| 2 | Brahman | Brahman | 263 | 18 , 6 |
| 3 | Bos taurus | B_tau | 22 | 1 , 6 |
| 4 | Bos taurus×Bos indicus | Btau×Bind | 43 | 3 , 0 |
| 5 | Criollas para Carne | Criolla_Car | 103 | 7 , 3 |
| 6 | Criollas para Doble Propósito | Criolla_Dpr | 189 | 13 , 4 |
| 7 | Criollas para leche | Criolla_Lec | 39 | 2 , 8 |
| 8 | Cruces para Carne | Cruce_Car | 68 | 4 , 8 |
| 9 | Cruces para Doble Propósito | Cruce_Dpr | 95 | 6 , 7 |
| 10 | Cruces para leche | Cruce_Lec | 64 | 4 , 5 |
| 11 | Guernsey | Guernsey | 49 | 3 , 5 |
| 12 | Holstein | Holstein | 89 | 6 , 3 |
| 13 | Holstein×Jersey | Hol×Jer | 34 | 2 , 4 |
| 14 | Jersey | Jersey | 158 | 11 , 2 |
| 15 | Pardo Suizo | Pardo Suizo | 49 | 3 , 5 |
| 16 | Simmental | Simmental | 50 | 3 , 5 |
| Población Total | 1412 | 100 |
El grupo clasificado como Bos Indicus (subpoblación 1, Cuadro 1) estuvo conformado mayormente por animales reportados como de tipo "cebú" y algunas razas presentes en números muy reducidos, tales como la Indubrasil, Gyr o Nellore. La subpoblación 2 estuvo conformada por animales reportados como "Brahman", aunque esta denominación en muchos casos corresponde a animales que no son de raza pura. La subpoblación 3 agrupó principalmente animales reportados como cruces entre razas Holstein, Pardo Suizo, Guernsey o Jersey, y algunos pocos animales de raza Chiannina y Hereford. El grupo Bos taurus×Bos indicus (subpoblación 4, Cuadro 1) incluyó principalmente cruces entre razas taurinas como la Holstein, Pardo Suizo o Jersey, con otras cebuinas como la Brahman.
En muchos casos el grupo racial se reportó originalmente como "Criollo". Cabe aclarar que esta denominación es utilizada actualmente en Costa Rica de manera un tanto arbitraria, ya que no corresponde a la definición real del término Criollo, la cual hace alusión a grupos raciales descendientes del ganado tipo Bos taurus procedente de la península Ibérica en el siglo XVI (FAO 2012, Quirós 2006). En la práctica, el término es actualmente utilizado de manera indistinta para describir cruces indefinidos entre razas Bos taurus×Bos indicus, o inclusive para describir ganado tipo Bos indicus de raza indefinida. Por esta razón, para el presente estudio el tipo racial reportado como Criollo se subdividió en 3 grupos en función del sistema de producción (subpoblaciones 5, 6, y 7; Cuadro 1). Lo mismo sucedió con animales reportados originalmente como "Cruce", ya que estos animales son producto de una mezcla indefinida de razas. Para el presente estudio estos casos también fueron clasificados en 3 grupos (subpoblaciones 8, 9 y 10; Cuadro 1) en función del sistema de producción (Carne, Leche o Doble Propósito).
La extracción de ADN del folículo piloso se realizó en el área de Biología Molecular y Genética del Laboratorio de Bioseguridad del SENASA a partir de una muestra de mechones de pelo de la cola con folículo, según el procedimiento descrito por Cordero et ál. (2013). Se analizó un conjunto de 18 marcadores, entre ellos 15 de los recomendados por FAO/ISAG (FAO 2011) para uso en bovinos: TGLA227, BM2113, TGLA53, ETH10, SPS115, TGLA126, TGLA122, INRA23, ETH3, ETH225, BM1824, BM1818, CSRM60, CSSM66 y ILSTS006, más otros 3 marcadores adicionales: SPS113, RM067 y MGTG4B. Los productos de la amplificación fueron sometidos a electroforesis capilar según el procedimiento descrito por Cordero et ál. (2013). Los polimorfismos de los microsatélites fueron discriminados de acuerdo con sus patrones de fluorescencia y tamaño. Los datos se visualizaron e identificaron mediante el software GeneMapper v3.0 (Applied Byosistems 2002).
Cálculo de parámetros de diversidad genética por subgrupo racial
Con base en la información obtenida de los análisis de ADN se estimaron los siguientes parámetros genéticos por subgrupo racial: el número total de alelos (Na) observados; el número efectivo (Ne) de alelos, obtenido como Ne= 1/Σp2 con p= frecuencias alélicas por marcador microsatélite; el nivel de Heterocigosidad observada (Ho), obtenido como la proporción observada de individuos heterocigóticos; el nivel de Heterocigosidad esperada (He), obtenido como 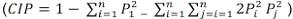 donde pi...pn son las frecuencias de los n alelos; el Contenido de Información Polimórfica y el Coeficiente de Consanguinidad
donde pi...pn son las frecuencias de los n alelos; el Contenido de Información Polimórfica y el Coeficiente de Consanguinidad  Además se compararon las frecuencias alélicas observadas contra las frecuencias alélicas esperadas bajo equilibrio Hardy-Weinberg por medio de la prueba Chicuadrado donde χ2 = estadístico de prueba; fo = frecuencias observadas y fe = frecuencias esperadas. Los cálculos anteriores se realizaron mediante el software especializado GenAlEx v.6.50 (Peakall y Smouse 2006, 2012) e Info-Gen (Balzarini y Di Rienzo 2004).
Además se compararon las frecuencias alélicas observadas contra las frecuencias alélicas esperadas bajo equilibrio Hardy-Weinberg por medio de la prueba Chicuadrado donde χ2 = estadístico de prueba; fo = frecuencias observadas y fe = frecuencias esperadas. Los cálculos anteriores se realizaron mediante el software especializado GenAlEx v.6.50 (Peakall y Smouse 2006, 2012) e Info-Gen (Balzarini y Di Rienzo 2004).
Análisis de estructura genética
Se realizó un Análisis de Varianza Molecular (AMOVA, Excoffier et ál. 1992) mediante el programa GenAlEx v6.5 (Peakall y Smouse 2006, 2012) con el fin de evaluar la existencia de diferenciación genética significativa entre las subpoblaciones bovinas formadas por los distintos grupos raciales identificados. Mediante este análisis se obtuvieron estimados de las varianzas genéticas entre razas (Ver), entre individuos (Vei) e intra-individuos (Vii). A partir de estos se calcularon los estadísticos F (Wright 1950), específicamente, F ST = Ver/(Vii + Vei + Ver), FIS = Vei/(Vii + Vei) y FIT = (Vei + Ver)/(Vii + Vei + Ver), para los cuales se obtuvo un estimado de significancia estadística mediante análisis de remuestreo (Felsenstein 1985) con 1000 permutaciones aleatorias.
Se calcularon además las distancias genéticas entre todos los pares de razas con 2 métodos diferentes: la distancia estándar (DS) de Nei (Nei 1972), calculada como () donde Px y Py son las homocigosis medias sobre loci en la población X e Y, respectivamente; y los estimados de diferenciación genética RST (Slatkin 1995), que se obtuvieron mediante la fórmula: 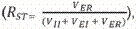 donde VER = varianza entre razas, VII = varianza intra-individuos y VEI = varianza entre individuos. Ambos estimados de distancia genética se calcularon mediante el programa GenAlEx v.6.5 (Peakall y Smouse 2006, 2012). Para RST se evaluó la significancia estadística mediante análisis de remuestreo (Felsenstein 1985) con 1000 permutaciones aleatorias.
donde VER = varianza entre razas, VII = varianza intra-individuos y VEI = varianza entre individuos. Ambos estimados de distancia genética se calcularon mediante el programa GenAlEx v.6.5 (Peakall y Smouse 2006, 2012). Para RST se evaluó la significancia estadística mediante análisis de remuestreo (Felsenstein 1985) con 1000 permutaciones aleatorias.
Para visualizar las relaciones genéticas entre las distintas subpoblaciones raciales, se construyó un dendrograma según el algoritmo Neighbor Joining (NJ) (Saitou y Nei 1987) con la medida de distancia estándar de Nei, según se implementa en el programa Poptree2 (Takezaki et ál. 2010). La consistencia de las ramificaciones del dendrograma se evaluó mediante análisis de remuestreo (Felsenstein 1985) con 1000 permutaciones. Para efectos comparativos se generó un segundo dendrograma con el algoritmo de Ward (Ward 1963) con distancias basadas en el índice de similitud de Dice (Dice 1945), por medio del programa Info-Gen (Balzarini y Di Rienzo 2004).
Resultados y discusión
Diversidad genética dentro de los subgrupos raciales
El número promedio de alelos (Na) por locus en las subpoblaciones raciales fue de 10,3 (Cuadro 2). Los valores obtenidos variaron desde un mínimo de 8 alelos (Holstein×Jersey) hasta un máximo de 13 alelos (Criollas para doble propósito). En lo que se refiere al valor de Ne, el promedio para todas las subpoblaciones raciales fue de 5,04. Los valores más bajos de Ne los presentaron las subpoblaciones Jersey (4,18) y Holstein×Jersey (4,22); mientras que las subpoblaciones Criollas para doble propósito y Bos taurus×Bos indicus presentaron los valores más altos de Ne (5,63 y 5,64 respectivamente).
EI número promedio de alelos detectado en el presente estudio es mayor que el reportado en estudios similares (7,2: Lirón et ál. 2006, 8,5: Egito et ál. 2007, 6,7: Ginja et ál. 2009, 7,5: Acosta et ál. 2012). Las diferencias pueden estar ligadas al hecho de que en los estudios mencionados los muestreos fueron dirigidos a razas específicas, con un menor número de muestras y explotaciones. En el presente estudio el muestreo fue aleatorio con el fin de obtener representatividad de la estructura de la población, lo que redunda en una mayor diversidad racial y genética. Además se contó con mayor número de explotaciones muestreadas en comparación con la mayoría de estudios similares. Otro factor que puede contribuir a las diferencias
Cuadro 2: Número total (Na) y efectivo (Ne) de alelos, heterocigosis observada (Ho) y esperada (He), contenido de información polimórfica (CIP), coeficiente de endogamia (FIS ) y número de marcadores según valor de significancia1 de la prueba Hardy-Weinberg (Prob>X2) para cada uno de los subgrupos raciales.
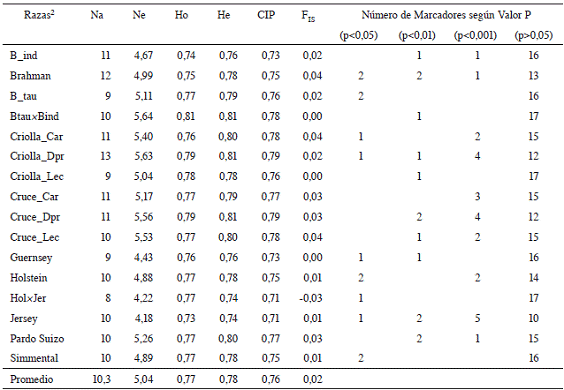
1Significativo (p<0,05; p<0,01; p<0,001); NS= No significativo (p>0,05).2B_ind: Bos indicus, B_tau: Bos taurus, Btau×Bind: Bos taurus×Bos indicus, Criolla_Car: Criollas para Carne, Criolla_Dpr: Criollas para Doble Propósito, Criolla_Lec: Criollas para leche, Cruce_Car: Cruces para Carne, Cruce_Dpr: Cruces para Doble Propósito, Cruce_Lec: Cruces para leche, Hol×Jer: Holstein×Jersey.
En cuanto a la heterocigosis observada (Ho), el promedio entre razas fue de 0,77, con un valor mínimo de 0,73 (Jersey) y un máximo de 0,81 (Bos taurus×Bos indicus). La Heterocigosis esperada (He) promedio resultó ser de 0,78, con variaciones entre 0,74 (Jersey y Holstein×Jersey) a 0,81 (Bos taurus×Bos indicus, Criollas para doble propósito y Cruces para doble propósito). En general las mayores heterocigosis se expresan en los tipos raciales denominados criollos o cruces, lo que es consistente con su heterogeneidad fenotípica.
Cabe destacar que en la mayoría de los grupos raciales se presentó déficit de heterocigotos (Ho<He). Este déficit de heterocigotos se interpreta generalmente como una desviación de los apareamientos aleatorios, en este caso con ligera tendencia a la consanguinidad. En el caso de las poblaciones de ganado bovino, existe un efecto marcado de la selección artificial y el control de los apareamientos realizado por los criadores, lo que sin duda puede jugar un papel importante.
En 3 subpoblaciones (Bos taurus×Bos indicus, Criollas para leche y Guernsey) los valores de He no difieren significativamente de Ho, lo que sugiere un estado de equilibrio HW donde los apareamientos ocurren al azar. En los sistemas ganaderos de Costa Rica las razas criollas lecheras se utilizan principalmente para establecer cruces con Bos Taurus, en especial Holstein y Jersey, y/o Bos indicus con la finalidad de obtener adaptabilidad y resistencia al medio (Vilaboa et ál. 2012). Sin embargo en otros casos se han desechado las razas criollas y han optado por el uso de otras razas o cruces Bos taurus×Bos indicus para la producción de leche, al relegar en importancia, la utilización de las razas criollas lecheras (Vilaboa et ál. 2012). En Costa Rica las razas que más se utilizan en cruzamientos para ganadería de carne son las cebuínas y Simmental y sus respectivos cruces con el fin de mejorar la calidad de la carne y pesos al destete (Hidalgo et ál. 2004).
El promedio de Ho obtenido en este estudio (0,77) es superior a valores reportados en otros estudios similares (0,75: Lirón et ál. 2006, 0,68: Egito et ál. 2007, 0,67: Ginja et ál. 2009, 0,70: Acosta et ál. 2012). En cuanto a la He encontrada en este estudio (0,78), también es superior a otros valores reportados previamente (0,73: Lirón et ál. 2006, 0,74: Egito et ál. 2007, 0,69: Ginja et ál. 2009, 0,73: Acosta et ál. 2012).
Con respecto al contenido de información polimórfica (CIP), se puede observar que todos los microsatélites utilizados estuvieron en la categoría de altamente informativos (>0,50). El coeficiente de endogamia (FIS ) presentó un valor promedio de 0,02, que osciló entre -0,03 (Holstein×Jersey) a 0,04 (Brahman, Criollas para carne y Cruces para leche). Un FIS de 0, como el encontrado en las subpoblaciones raciales Bos taurus×Bos indicus, Criollas para leche y Guernsey, significa que se encuentran en equilibrio HW, puesto que la heterocigosidad observada (Ho) es semejante a la heterocigosidad esperada (He). El resultado en esta subpoblación puede ser debido al cruce entre 2 grupos raciales, lo que expresa un exceso de heterocigotos (Ho>He) y por tanto un nivel de consanguinidad muy bajo. El valor promedio de FIS en este estudio es superior al -0,03 encontrado por Lirón et ál. (2006) y al 0,004 obtenido por Acosta et ál. (2012), e inferior al 0,09 encontrado por Egito et ál. (2007) y al 0,03 reportado por Ginja et ál. (2009).
Con respecto a la prueba de equilibrio de Hardy-Weinberg (HW) la desviación del equilibrio HW no fue significativa (p>0,05) en la mayoría de los loci (Cuadro 2) dentro de las subpoblaciones raciales. El grupo con mayor número de loci en desequilibrio HW (p<0,05) fue Jersey (8 loci), mientras que los grupos Bos taurus×Bos indicus, Criollas para leche y Holstein×Jersey presentaron solo 1 locus en desequilibrio HW.
Análisis de estructura genética
Según los resultados del AMOVA la proporción de la variabilidad genética atribuible a la diferencia entre las subpoblaciones raciales (Ver), entre individuos (Vei) e intra individuos (Vii) fue de 3%, 3% y 94% respectivamente. En estudios similares se han reportado mayores valores para Ver. Lirón et ál. (2006) reportaron que el 8,8% de la variación genética total correspondió a las diferencias entre las poblaciones (razas cebú y taurinas), mientras que el 91,2% se explicó por las diferencias entre e intra individuos. Cañón et ál. (2001) indicaron que alrededor del 7% de la variación genética total correspondió a las diferencias entre los grupos raciales, mientras que el 93% restante correspondió a las diferencias entre e intra individuos. Por otra parte, Egito et ál. (2007) reportaron un valor aún mayor (12%) para la variación genética atribuible a las diferencias entre los grupos raciales.
El valor menor obtenido en el presente estudio puede estar ligado nuevamente a las características del muestreo y a la clasificación de animales en grupos raciales afines. Los grupos raciales combinados (p.e criollo o cruces) están conformados por animales altamente heterogéneos, lo que magnifica la varianza dentro de grupo (entre e intra individuos) en comparación con la varianza entre los grupos (entre razas). Muchos de estos grupos además están altamente relacionados entre sí, lo que se confirma posteriormente en los dendrogramas.
La contribución de los 18 marcadores para la diferenciación de los grupos raciales fue estimada por los estadísticos F (datos no mostrados). Los valores promedio de FIS, FIT y FST fueron de 0,05, 0,02 y 0,03 respectivamente. Al analizar los valores de FST obtenidos para los diferentes marcadores se observó que los marcadores ETH10, ETH225 y TGLA227 son los que más contribuyen a las diferencias entre razas, mientras que los marcadores BM1818, BM1824, ILSTS006, INRA23, MGTG4B, RM067, SPS113 y TGLA53 son los que mostraron menor variabilidad. El valor promedio de FST observado en el presente estudio tiende a ser inferior al obtenido en estudios similares (0,07: Jordana et ál. 2003, 0,10: Egito et ál. 2007, 0,04: Sun et ál. 2008, 0,09: Acosta et ál. 2012).
Como era de esperarse, en general se observan valores de distancias genéticas bajas entre los grupos raciales Bos indicus y sus cruces, y entre los grupos Bos taurus y sus cruces (Cuadro 3). Por el contrario, se observan distancias altas entre los grupos raciales de leche (Bos taurus) con respecto a las razas de carne (Bos indicus). Se observan además distancias reducidas entre las razas criollas y cruces de los grupos raciales anteriores.
Las distancias genéticas más bajas se obtuvieron para grupos genéticamente afines, por ejemplo valores de 0,02 se obtuvieron entre las subpoblaciones Brahman vs. Bos indicus; Criollas para doble propósito vs. Criollas para carne; Cruces para carne vs. Criollas para carne; Cruces para doble propósito vs. Criollas para carne; Cruces para carne vs. Criollas para doble propósito o Cruces para doble propósito vs. Criollas para doble propósito. La reducida distancia genética entre los grupos Criollos y los Cruces, dentro de cada sistema de producción, confirma que el término Criollo se utiliza localmente para describir ganado obtenido por cruzamientos entre razas Bos taurus o Bos indicus, en proporciones variables.
El uso incorrecto o confuso de denominaciones raciales es un problema reportado a nivel global.
Recientemente, una consultoría realizada por la comisión de recursos genéticos de la FAO propuso descontinuar el uso de términos tales como raza nativa, indígena o autóctona (FAO 2012). En su lugar, se ha sugerido que, para efectos de caracterización genética, se utilicen criterios de clasificación basados en adaptabilidad racial. Se considera una raza adaptada aquella que ha permanecido en un país por al menos 40 años o 6 generaciones, lo que se considera suficiente para adecuarse a uno o más de los sistemas tradicionales de producción (FAO 2012). En contraposición se recomienda el uso del término exótico para aquellas razas introducidas recientemente o que son importadas de manera continua.
Por el contrario, las distancias genéticas más altas se observaron entre las subpoblaciones Holstein vs. Bos indicus (0,42); Jersey vs. Bos indicus (0,45); Holstein×Jersey vs. Bos indicus (0,47) y Guernsey vs. Bos indicus (0,48). Estos resultados eran de esperarse, debido a que estos grupos raciales pertenecen a 2 subespecies diferentes (Bos taurus y Bos indicus). En este sentido es importante recalcar que, a pesar de que el número de marcadores es reducido, sí fue posible establecer una discriminación clara a nivel de las frecuencias alélicas entre las subpoblaciones de distintas especies y aún entre razas puras.
Los valores más bajos de distancias genéticas encontrados en este estudio son inferiores al 0,12 reportado por Brasil et ál. (2013) entre las razas Brahman vs. Nellore; así como el 0,10 reportado por Egito et ál. (2007) entre las razas Nellore vs. Gyr. Los valores más altos de distancias genéticas encontrados en este estudio son similares a los reportados por Brasil et ál. (2013) entre las razas Nellore vs. Holstein (0,44); Nellore vs. Jersey (0,48); Brahman vs. Holstein (0,48); Gyr vs. Holstein (0,49); Gyr vs. Jersey (0,50) y Brahman vs. Jersey (0,51), así como a los reportados por Egito et ál. (2007) entre las razas Holstein vs. Gyr (0,34), Jersey vs. Gyr (0,37), Nellore vs. Holstein (0,38) o Nellore vs. Jersey (0,38).
En el Cuadro 3 se presentan también estimados de diferenciación genética RST (Slatkin 1995) entre todos los pares de razas con su significancia estadística. Para la mayoría de las subpoblaciones raciales, la diferenciación genética fue altamente significativa (p<0,001). En general se observó una buena correlación entre los valores de diferenciación genética (RST) por pares de razas y las distancias genéticas de Nei.
Como era de esperarse, se observó una mayor diferenciación entre pares de diferente subespecie (Bos taurus vs. Bos indicus), mientras que se observó la menor diferenciación para grupos raciales más afines en origen. La menor diferenciación genética (RST =0) se presentó entre las subpoblaciones Bos taurus×Bos indicus vs. Criollas para leche; Bos taurus×Bos indicus vs. Cruces para doble propósito; Criollas para doble propósito vs. Cruces para doble propósito; Pardo Suizo vs. Simmental. Además la diferenciación genética en estas subpoblaciones raciales no fue significativa (p>0,05).
Cabe mencionar que los valores de RST entre las subpoblaciones Bos taurus×Bos indicus vs. Criollas para leche; Pardo Suizo vs. Simmental fueron negativos (-0,005 y -0,003 respectivamente) y por tanto el programa los convierte a cero. Los valores negativos pueden ocurrir cuando la variación dentro de poblaciones es mayor que la variación entre poblaciones, que puede ser el caso en estas subpoblaciones ya que están conformadas por animales muy heterogéneos.
Cuadro 3: Matriz de distancias genéticas de Nei (bajo diagonal) y estimados de diferenciación genética (RST sobre la diagonal) entre 16 subpoblaciones raciales1 bovinas de Costa Rica.
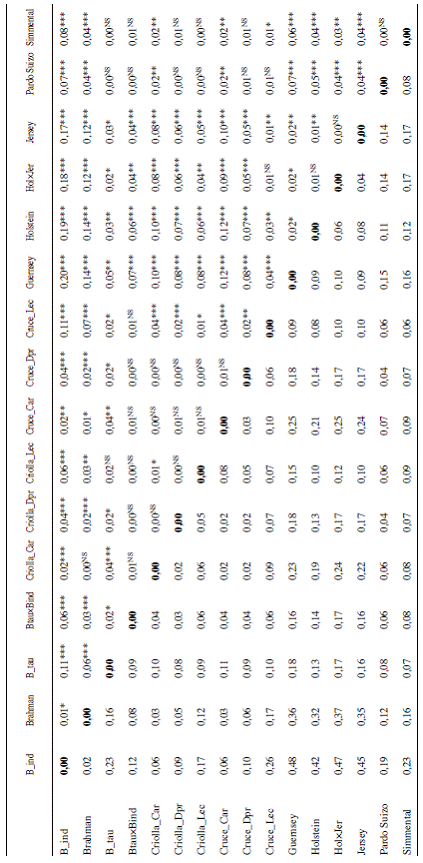
1B_ind: Bos indicus, B_tau: Bos taurus, Btau×Bind:Bos taurus×Bos indicus, Criolla_Car: Criollas para Carne, Criolla_Dpr:Criollas para Doble Propósito, Criolla_Lec: Criollas para leche, Cruce_Car: Cruces para Carne, Cruce_Dpr: Cruces para Doble Propósito, Cruce_Lec: Cruces para leche, Hol×Jer: Holstein×Jersey. NS= No significativo (p>0,05); significativo (*p<0,05; **p<0,01; ***p<0,001).
Por otra parte las subpoblaciones Bos indicus vs. Jersey (0,17); Bos indicus vs. Holstein×Jersey (0,18); Bos indicus vs. Holstein (0,19) y Bos indicus vs. Guernsey (0,20) presentaron la mayor diferenciación genética, al ser además, altamente significativa (p<0,001). Esto sugiere que estos grupos raciales pueden ser claramente discriminados mediante los 18 marcadores utilizados en el presente estudio.
A partir de las distancias estándar de Nei (1972), representadas en el Cuadro 3 se construyó un dendrograma con el algoritmo de Neighbor-Joining (Saitou y Nei 1987) (Figura 1(a)). Se observa la formación de 2 grupos principales, con las razas cebuinas (abajo) y las taurinas (arriba). También se observa cierta separación de las razas Simmental y Pardo Suizo con respecto a las demás razas taurinas. Los valores de confianza para las separaciones nodales, obtenidos por remuestreo, fueron muy variables entre un mínimo de 41 y un máximo de 100. Estos valores tienden a ser más altos para los nodos que separan razas puras, posiblemente porque su estructura genética está más claramente diferenciada.
Todos los grupos raciales que son genéticamente afines se encuentran en los mismos grupos, con la excepción del grupo racial Criollas para leche. Sin embargo, es posible que este grupo racial tenga una importante influencia de grupos raciales Bos indicus. En este sentido es importante recalcar, que el número de microsatélites empleados sí permitió establecer una discriminación clara a nivel de las frecuencias alélicas entre las razas puras.
En la Figura 1(b) se muestra un dendrograma para el cual se utilizó el método de agrupamiento de Ward (Ward 1963) sobre una matriz de distancia basada en el índice de similitud de Dice (Dice 1945). El análisis de conglomerados permitió clasificar básicamente 3 grupos de razas de acuerdo con la subespecie bovina y al sistema de producción a la que pertenecen. Un grupo lo conforman 5 subpoblaciones raciales, con mayor influencia Bos taurus y tradicionalmente lecheras de origen Europeo (Jersey, Holstein×Jersey, Holstein, Guernsey y Cruces para leche) localizados en la parte superior del dendrograma. Por otro lado, un grupo adicional lo conforman 5 subpoblaciones raciales con influencia Bos taurus (Criollas para leche, Bos taurus×Bos indicus, Simmental, Pardo Suizo y Bos taurus) localizadas en una posición intermedia en el dendrograma. En este grupo, principalmente las razas Simmental, Pardo Suizo y Bos taurus han evolucionado como razas Doble Propósito de alta producción, por tanto forman una agrupación adicional.
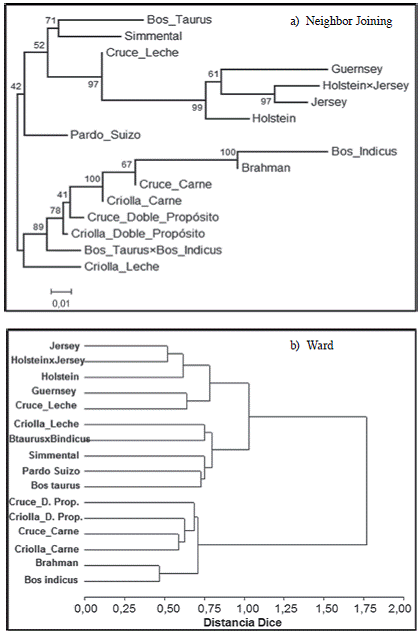
Figura 1: Relaciones genéticas entre 16 subpoblaciones raciales bovinas de Costa Rica obtenidas por los algoritmos a) Neighbor Joining (Saitou y Nei 1987) (valores en bifurcaciones corresponden a niveles de confianza basados en 1000 iteraciones de remuestreo), y b) Ward (Ward 1963).
Finalmente, un grupo lo conforman principalmente 6 subpoblaciones raciales con mayor influencia Bos indicus (Cruces para doble propósito, Criollas para doble propósito, Cruces para carne, Criollas para carne, Brahman y Bos indicus) localizados abajo del dendrograma. Es importante destacar que este subgrupo ha evolucionado históricamente como razas orientadas a la producción de carne, además de ser genéticamente más parecidos por ser originarias de la India.
Estas 3 grandes agrupaciones de razas representan a las subpoblaciones bovinas como las más diferentes, ya que se unen a la mayor distancia genética, debido a sus orígenes geográficos distintos y al sistema de producción.
Los dendrogramas también ratificaron que las denominaciones Criollo y Cruce dentro de un mismo sistema de producción, corresponden en realidad a animales con características genéticas muy similares. Las diferencias genéticas de estos mismos grupos entre los diferentes sistemas de producción (ej. Criollo Carne vs. Criollo Doble Propósito vs. Criollo Leche) está en función principalmente de las proporciones relativas de razas Bos indicus vs. Bos taurus presentes en el cruce.

