










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgronomía Costarricense
Print version ISSN 0377-9424
Agron. Costarricense vol.35 n.1 San Pedro de Montes de Oca Jun. 2011
Efecto de la inoculación de Azotobacter chroococcum y Glomus intraradices en el crecimiento y nutrición de plántulas de papaya en fase de vivero
Effect of time of inoculation of Azotobacter and mycorrhizal fungi on growth and content nutrient of papaya seedlings in nursery phase
Effect of time of inoculation of Azotobacter and mycorrhizal fungi on growth and content nutrient of papaya seedlings in nursery phase
Maricela Constantino*, Regino Gómez1/*, José David Álvarez*, Juan Manuel Pat*, Elda Guadalupe Espín**
1 Correo electrónico: regomez@ecosur.mx
* El Colegio de la Frontera Sur. Carretera Villahermosa-Reforma km 15,5. Ranchería Guineo, sección II. CP 86280. Villahermosa, Tabasco. México.
** Instituto de Biotecnología. Universidad Autónoma de México. Av. Universidad Número 2001, Col. Chamilpa C.P. 62210. Cuernavaca, Morelos C.P. 62250. México.
Dirección para correspondencia
Resumen
En el presente estudio se evaluó la etapa y el número de aplicaciones de los biofertilizantes (Azotobacter chroococcum y Glomus intraradices), sobre el crecimiento, biomasa y nutrición de papaya en fase de vivero. También se estudió el efecto de la materia orgánica y su interacción con los biofertilizantes aplicados. Se realizaron 2 experimentos; en el primero se aplicaron 2 inoculaciones, en semillas y después en plántulas, 30 días después de la emergencia. En el segundo se aplicó una inoculación simple en plántulas, 30 días después de la emergencia. Estos experimentos se realizaron utilizando un diseño factorial 5x4 (5 tipos de sustratos x 4 tipos de biofertilizantes). Todos los tratamientos fueron aplicados como inoculantes simples o combinados (A. chroococcum y G. intraradices) para un total de 20 tratamientos, con un diseño completamente al azar. La doble inoculación (semilla y plántula) promovió un mayor crecimiento y biomasa en el cultivo, en comparación con la inoculación simple (solo en plántulas), cuando se adicionó una dosis intermedia de materia orgánica (25 a 35%) y se aplicó G. intraradices como simple inoculante. Sin embargo, la inoculación simple o combinada no modificó el contenido de nutrientes en las plántulas de papaya.
Palabras clave: Biofertilizantes, Glomus intraradices, rizobacterias, inoculación de semillas, materia orgánica.
Abstract
The effect of time of inoculation (at sowing or after seedling emergence) and the number of applications using 2 biofertilizer (Azotobacter chroococum and Glomus intraradices) were tested on growth, biomass and nutrition of papaya, at the nursery phase. The effect of organic matter and its interaction with the biofertilizers also was evaluated. Two experiments were realized; in the first one 2 inoculations were applied, on seeds and afterwards on seedlings, 30 days post emergency. In the second experiment a single inoculation was applied on seedlings, 30 days post emergence. These experiments were performed using a 5x4 factorial design (5 substrates x 4 biofertilizer types). All treatments were applied as single or combined inoculants (A. chroococcum and G. intraradices) for a total of 20 treatments, in a completely randomized design. The double inoculation (seed and seedling) promoted higher growth and biomass of the crop than single inoculation (only seedlings), when organic matter was added at an intermediate dose (25 or 35%) and G. intraradices was applied as a single inoculant. However, the single or double inoculation did not modify the nutrient content in papaya seedlings.
Keywords: Biofertilizer, Glomus intraradices, rhizobacteria, seed inoculation, organic matter.
Introducción
El cultivo de papaya requiere un suelo fértil y rico en materia orgánica, debido a su rápido crecimiento (Rodríguez 1984). La incorporación de materia orgánica y biofertilizantes son algunas de las prácticas que se han aplicado en la preparación de sustratos empleados en los viveros (Verma et al. 2008). La materia orgánica actúa como una fuente esencial de nutrientes para las plantas y mejora las propiedades físicas, químicas y biológicas del suelo (Mokwunye et al. 1996).
Existen diversos trabajos donde la aplicación de biofertilizantes en la etapa de vivero ha incrementado el porcentaje de germinación, el crecimiento, desarrollo y salud de las plántulas de papaya, que mejoran la calidad y reducen el tiempo de permanencia de las plántulas en el vivero. Dentro de los microorganismos que han sido empleados como biofertilizantes en el cultivo de papaya se encuentran: Azospirillum brasilense y Glomus claroideum (Alarcon et al. 2002), Trichoderma viridae y Azotobacter chroococcum (Cupull et al. 2002), Trichoderma harzianum y hongos micorrízicos (Mesa et al. 2006, Wasy et al. 2010). También es importante mencionar que las plántulas pre-inoculadas con micorrizas son más tolerantes al estrés que sufren al trasplante en comparación a las plantas no micorrizadas o plantas inoculadas al momento del trasplante (Valdés et al. 1993, Waterer y Coltman 1988).
Los métodos de biofertilización más empleados han sido la aplicación en semillas, la inmersión de plántulas y la aplicación en el suelo (Hegde 1992, Sharma et al. 2008), el método de aplicación dependerá del tipo de cultivo. La formulación del inóculo, el método de aplicación y el almacenamiento del producto son críticos para el éxito del los productos biológicos (Chen 2006). Asimismo, el momento de la inoculación (en la siembra o en la emergencia de las plántulas, o varios días después de la aparición de 1 a 4 hojas verdaderas), también parece ser crucial en determinar la colonización exitosa de los inoculantes microbianos e influenciar el crecimiento de las plantas (Bashan 1986).
El objetivo del presente trabajo fue evaluar el efecto de la biofertilización y la materia orgánica en el crecimiento, biomasa y nutrición de plántulas de papaya. También se determinó el mejor momento de la inoculación de los biofertilizantes, en el cultivo de la papaya en fase de vivero.
Materiales y métodos
La investigación se realizó en la etapa comprendida de marzo a junio de 2009, en el campo experimental de El Colegio de la Frontera Sur, Unidad Villahermosa, ubicada en la Ranchería Guineo Segunda Sección, Municipio Centro, Tabasco, México.
Para evaluar el efecto de los biofertilizantes y la materia orgánica en el crecimiento y nutrición de plántulas de papaya en fase de vivero, se realizaron 2 experimentos: en el primer experimento se aplicó una doble inoculación de los biofertilizantes primero en las semillas y 30 días después de la emergencia de las plántulas. En el segundo experimento se aplicó una simple inoculación a plántulas de papaya con 30 días de crecimiento. Cada experimento se realizó de forma independiente. El primer experimento se desarrollo de marzo a mayo y el segundo experimento de abril a junio.
Material microbiológico: En los experimentos se utilizó la cepa de Azotobacter chroococcum, aisladas por el Instituto de Investigaciones Fundamentales en Agricultura Tropical (INIFAT) de la Habana, Cuba y reproducidas en los laboratorios de ECOSUR. Para la formulación del inoculante líquido, A. chroococcum fue cultivado en medio líquido Ashby enriquecido con NH4NO3 (3 g.l-1) y extracto de levadura (0,1 g.l-1), manteniéndose con agitación orbital (150 rpm) a 30°C por 60 h, hasta obtener una concentración de 1x109 unidades formadoras de colonias por ml (ufc.m.l-1). El inóculo micorrízico empleado fue Glomus intraradices, con una concentración de 1050 esporas en 100 g de Tezontle, que incluye raíces de alfalfa con un 85% de colonización, proporcionado por el área de Microbiología de El Colegio de Postgraduados, Montecillo, México.
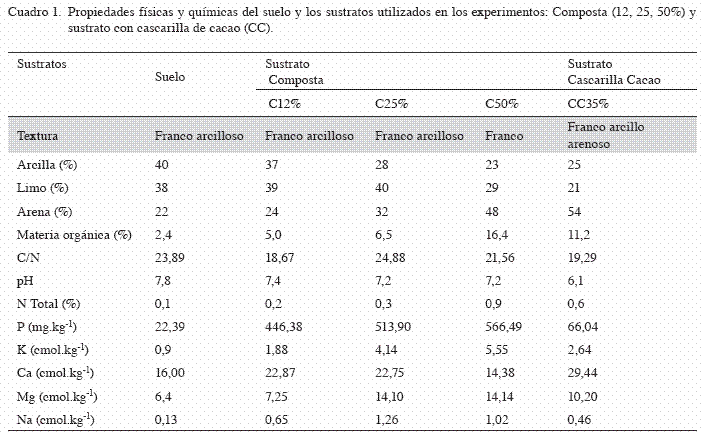
Sustratos: Se utilizaron 2 tipos de sustratos: a) sustrato con composta hecha con residuos de hojarasca (suelo: composta, con base en peso/ peso, en 4 diferentes proporciones porcentuales 100:0, 88:12, 75:25, 50:50%, y b) sustrato con cascarilla de cacao, previamente seca y tamizada; la proporción fue hecha con base en peso/peso y se hizo en proporción porcentual 30:35:35 de arena:suelo:cascarilla de cacao. Después de realizar las mezclas se esterilizaron en autoclave a 1,5 atm por 2 h, por 3 días consecutivos. El sustrato con cascarilla de cacao es utilizado en diversos viveros de papaya de Tabasco, por ser un material disponible en la región. Las características físicas y químicas del suelo y los sustratos se describen en el Cuadro 1.
Material vegetal: Se emplearon semillas de papaya cv. Maradol previamente desinfectadas en una solución de peróxido de hidrogeno al 10% por 3 min y posteriormente fueron lavadas 3 veces con agua destilada estéril. En seguida, se aplicó un tratamiento de pre-germinación que consistió en la inmersión de las semillas en agua destilada estéril a temperatura alternada de 15-35°C por 24 y 4 h respectivamente. Inmediatamente después las semillas fueron tratadas conforme a los experimentos descritos a continuación.
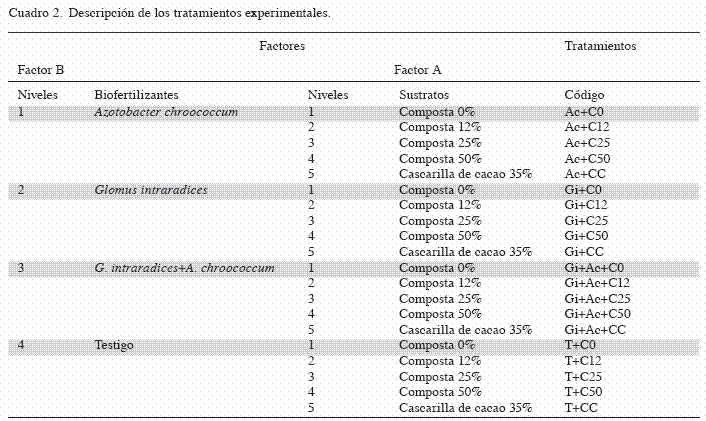
Experimento 1: Evaluación de la doble inoculación de los biofertilizantes. La aplicación de los biofertilizantes en semillas se realizó inmediatamente después del tratamiento pre-germinativo, de acuerdo con los tratamientos establecidos (Cuadro 2). Para el tratamiento con Azotobacter las semillas se sumergieron por una hora en el inoculante líquido bacteriano. Posteriormente, las semillas se sembraron en charolas de germinación con 24 cavidades, sembrándose una semilla por cavidad y se empleo una charola por tratamiento. Las charolas fueron llenadas con el sustrato cascarilla de cacao.
El inóculo de G. intraradices se aplicó directamente en el sustrato, 3 g por cavidad justo antes de la siembra de la semilla. En los tratamientos con las mezclas de A. chroococcum + G. intraradices, las semillas se sumergieron por una hora en el biofertilizante bacteriano y posteriormente se sembró a razón de una semilla por cavidad, donde previamente se había inoculado cada cavidad con 3 g de G. intraradices. A los 30 días después de la emergencia de las plántulas, se realizó la segunda inoculación en el trasplante en macetas plásticas de 1,5 kg. Cada maceta fue llenada con los respectivos sustratos. La inoculación de A. chroococcum se hizo directamente al sistema radical, con la aplicación de 5 ml.plántula-1, mientras que la aplicación de G. intraradices se hizo con 3 g.plántula-1, también al sistema radical. Para el tratamiento combinado de A. chroococcum y G. intraradices, primero se aplicaron los 3 g de inóculo micorrízico y después los 5 ml.plántula-1 del inoculante bacteriano. Las plántulas se mantuvieron bajo una temperatura entre 26 y 38ºC y una humedad relativa promedio de 45%, con riego diario durante 60 días.
Experimento 2: Evaluación de inoculación simple de los biofertilizantes. En la inoculación simple, los biofertilizantes fueron aplicados directamente en el sistema radical de las plántulas con 30 días de crecimiento. Para la obtención de las plántulas, las semillas recibieron el tratamiento pre-germinativo descrito en el experimento 1. Posteriormente, las semillas fueron sembradas en charolas de germinación llenas con el sustrato con cascarilla de cacao y se dejaron crecer por 30 días, bajo las condiciones ambientales de vivero descritas anteriormente. Después de este tiempo, las plántulas fueron trasplantadas a macetas de 1,5 kg con los respectivos sustratos en cada tratamiento.
La biofertilización de las plántulas se realizó al momento del trasplante en el sistema radical, de acuerdo con cada uno de los tratamientos (Cuadro 2). La biofertilización simple y combinada de A. chroococcum y G. intraradices se realizó de igual forma que se hizo en la segunda aplicación del primer experimento.
Diseño de experimento: En los 2 experimentos se aplicó un diseño factorial 5x4 con arreglo completamente al azar, evaluándose 2 factores: el Factor A: Sustrato. Se utilizaron 5 sustratos (composta al 0, 12, 25 y 50% y cascarilla de cacao al 35%, correspondientes a 5 niveles). El Factor B: Biofertilizante. Se utilizaron 2 biofertilizantes, la mezcla y un testigo sin biofertilizar (A. chroococcum, G. intraradices, A. chroococcum + G. intraradices y un Testigo, correspondientes a 4 niveles). Para dar un total de 20 tratamientos con 3 repeticiones cada uno (Cuadro 2).
Variables de crecimiento y nutrición evaluadas. Las variables de crecimiento evaluadas a los 90 días después de la emergencia fueron: altura de la planta (cm), diámetro del tallo (mm), biomasa fresca (g) y biomasa seca (g). El diámetro se midió en la base del tallo (unión del tallo con la raíz). La altura de las plantas se midió de la base del tallo hasta el ápice. El peso seco de las plantas se evaluó después de la deshidratación en estufa a 60°C durante 72 h y posteriormente se pesaron en una balanza de precisión. Para el análisis de los nutrientes, el contenido de N fue determinado por el método semi-micro Kjeldahl (Bremmer 1969). El contenido de P y K fueron determinados por HNO3-HClO4 y fueron medidos por el método espectrofotométrico con la mezcla vanadomolibdica y espectrometría de absorción atómica espectivamente.
Variables microbiológicas evaluadas. La población de A. chroococcum se determinó al pesar 10 g de suelo rizosférico por planta y se agregó en un matraz con 100 ml de agua destilada estéril, por medio de una serie de diluciones de 10-1 a 10-5. Se inocularon por triplicado 100 μl de las 3 últimas diluciones en cajas de petri con medio de cultivo Burk-sacarosa (BS) adicionado con Acido Nalidixico (20 μg.ml-1 de medio de cultivo). El porcentaje de colonización micorrízica se evaluó por medio del método de clareo y tinción de Phillips y Hayman (1978).
Análisis estadístico. Se realizó un ANOVA factorial y la comparación de medias se realizó con la prueba de Rangos múltiples de Duncan (p<0,05). Para conocer la posible variabilidad de los resultados obtenidos entre la doble y la simple biofertilización, se realizó la comparación de las medias con la prueba t de student para 2 muestras independientes. Los análisis se realizaron con el software estadístico STATISTICA versión 8.0.
Resultados
Efecto de inoculación simple y combinada de los biofertilizantes y su interacción con la materia orgánica en la población de A. chroococcum y en la colonización de G. intraradices
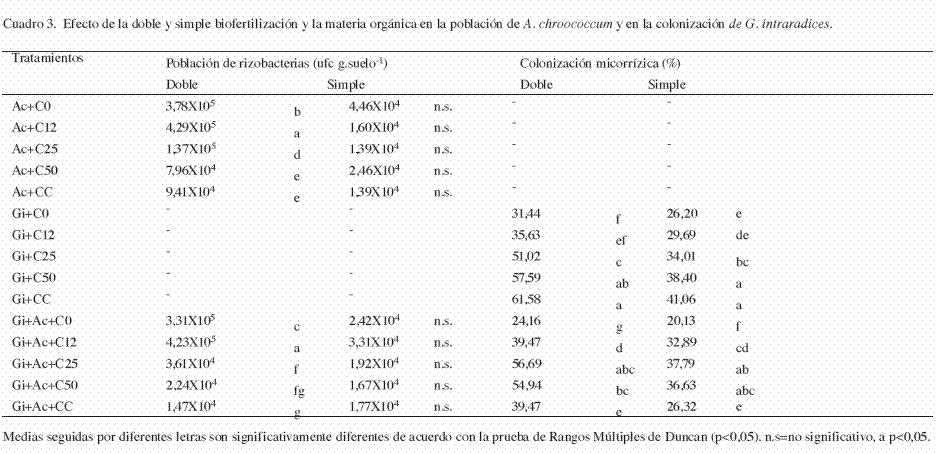
Experimento 1: A los 90 días después de la siembra, con doble biofertilización se observó que la población de A. chroococcum fue mayor a menor contenido de materia orgánica, siendo los tratamientos Ac+C12 y Gi+Ac+C12 los que registraron una concentración bacteriana significativamente mayor (p<0,05) a los demás tratamientos. Además, la población de A. chroococcum fue significativamente mayor (p<0,05) cuando se aplicó sin combinación con G. intraradices (Cuadro 3). La evaluación de la infección micorrízica en raíces de papaya con doble biofertilización, señala que los tratamientos Gi+CC, Gi+C50 y Gi+Ac+C25, fueron los que registraron un mayor porcentaje de colonización micorrízica (61,58; 57,59; 56,69 y 54,94%). En general, la aplicación simple de G. intraradices registró una mayor colonización en comparación a cuando se aplicó en combinación con A. chroococcum (Cuadro 3).
Experimento 2: En la simple biofertilización, no se encontraron diferencias significativas en la concentración de A. chroococcum en ninguno de los tratamientos evaluados, por lo que ni la adición de materia orgánica ni la aplicación combinada con G. intraradices, tuvieron un efecto en su población (Cuadro 3). De igual forma, no se encontraron diferencias significativas en la población de Azotobacter al aplicar una doble o una simple biofertilización (de acuerdo con la prueba t de Student, p>0,05). (Cuadro 3).
En la simple biofertilización, se observó una mayor colonización micorrízica a mayores dosis de materia orgánica adicionada. Los tratamientos Gi+CC, Gi+C50, Gi+Ac+C25 y Gi+Ac+C50 registraron los valores más altos de colonización, mientras que en los tratamientos sin adición de materia orgánica se observaron los valores más bajos de colonización micorrízica (Cuadro 3). Por otra parte, también se encontró que la colonización micorrízica fue significativamente mayor cuando se aplicó una doble inoculación (de acuerdo con la prueba t de Student, p<0,05).
Experimento 1: Efecto de la doble inoculación de los biofertilizantes y su interacción con la materia orgánica en el crecimiento, biomasa y nutrición vegetal
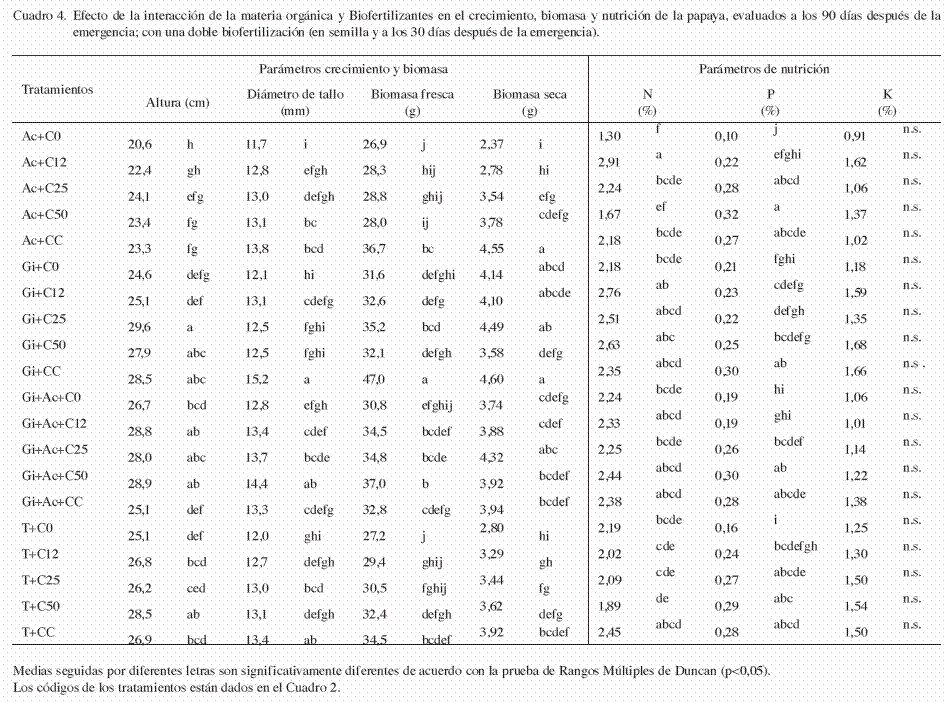
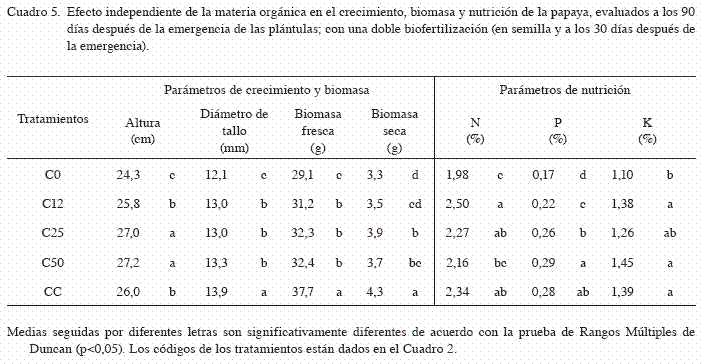
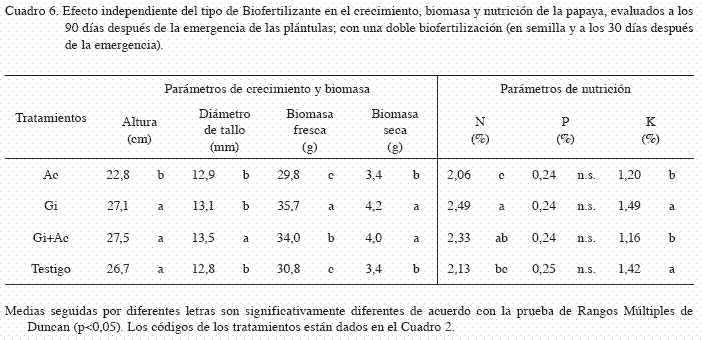
La altura promedio de las plántulas fue de 26,0 cm y fue significativamente afectada por la interacción entre el sustrato y el tipo de biofertilizante (p<0,05). Las plántulas de los tratamientos i+C25 presentaron la mayor altura (29,6 cm) (Cuadro 4). El efecto independiente del factor A (sustrato) en la altura, fue significativamente diferente (p<0,05), las plántulas crecidas en los sustratos con 50 y 25% de composta (C50 y C25) presentaron la mayor altura (27,2 y 27,0 cm, respectivamente) (Cuadro 5). Al evaluar el efecto independiente del factor B (biofertilizantes), se observó que la aplicación de Azotobacter afectó negativamente la altura de las plántulas en comparación a los demás tratamientos (Cuadro 6).
El diámetro de tallo promedio fue de 13,1 mm y fue significativamente afectado por la interacción entre el sustrato y el tipo de biofertilizante (p<0,05). Las plantas crecidas en el tratamiento Gi+CC, resultaron con un mayor diámetro (15,2 mm) y fueron significativamente mayor a los demás tratamientos (p<0,05) (Cuadro 4). El efecto independiente del factor A (sustrato) en el diámetro del tallo, presentó diferencias significativas (p<0,05). Las plántulas con mayor diámetro se registraron con la adición del sustrato CC (13,9 mm) (Cuadro 5). En cuanto al efecto del factor B (biofertilizante), el mayor diámetro del tallo se observó con la inoculación combinada de HMA y Azotobacter (Gi+Ac) (13,5 mm) (Cuadro 6).
La aplicación de materia orgánica incrementó la acumulación de biomasa fresca, obteniéndose en promedio 33,6 g.plántula-1, y fue significativamente afectada por la interacción entre el sustrato y el biofertilizante (p<0,05). Las plántulas crecidas en el tratamiento Gi+CC, acumularon la mayor biomasa fresca (47,0 g) (Cuadro 4). Asimismo, el efecto independiente del factor A (materia orgánica) en la biomasa fresca, presentó diferencias significativas (p<0,05), pero el mayor efecto se debió a la adición del sustrato CC (37,7 g) (Cuadro 5). De igual forma, el efecto independiente del factor B (biofertilizante) afectó significativamente la biomasa fresca (p<0,05), pero el mayor efecto se debió a la aplicación simple de HMA (Gi) (35,7 g) (Cuadro 6).
La biomasa seca promedio obtenida fue de 3,74 g.plántula-1 y fue significativamente afectada por la interacción entre la materia orgánica y el biofertilizante (p<0,05). Las plántulas crecidas en los tratamientos Gi+CC y Ac+CC, acumularon la mayor biomasa seca (4,60 y 4,55 g, respectivamente) (Cuadro 4). Asimismo, el efecto independiente del factor A (sustrato) en la biomasa seca fue significativo (p<0,05). La adición del sustrato CC, registró el mayor contenido de biomasa seca (4,25 g) (Cuadro 5). En cuanto al efecto del factor B (biofertilizante), también se encontraron diferencias significativas en la biomasa seca (p<0,05), pero el mayor efecto se debió a la inoculación simple de HMA (Gi) y en combinación con Azotobacter (Gi+Ac), con valores de 4,18 y 3,96 g, respectivamente (Cuadro 6).
El contenido promedio de nitrógeno en tejido vegetal fue de 2,25% y fue afectado significativamente por la interacción entre el sustrato y el biofertilizante (p<0,05). Las plántulas crecidas en los tratamientos Ac+C12 y Gi+C12, registraron los mayores contenidos de nitrógeno vegetal (2,91 y 2,76%, respectivamente) (Cuadro 4). El efecto independiente del factor A (sustrato) también fue significativo (p<0,05). Las plántulas crecidas en los sustratos con composta al 12%, sustrato CC y composta al 25% (C12, CC y C25), obtuvieron los mayores contenidos de nitrógeno (2,50, 2,34 y 2,27%, respectivamente) (Cuadro 5). De igual forma, el efecto independiente del factor B (biofertilizante) fue significativo (p<0,05), pero el mayor efecto se debió a la aplicación simple de HMA y su combinación con Azotobacter (Gi y Gi+Ac) (2,49 y 2,33%, respectivamente) (Cuadro 6).
El contenido promedio de fósforo fue de 0,24%, y también fue afectado significativamente por la interacción entre el sustrato y el biofertilizante (p<0,05). Las plántulas crecidas en los tratamientos Ac+C50, Gi+CC y Gi+Ac+C50, mostraron los mayores contenidos de fósforo vegetal (0,32, 0,30 y 0,30%, respectivamente) (Cuadro 4). El efecto independiente del factor A (sustrato) también fue significativo (p<0,05), pero el mayor efecto se debió a la aplicación de composta al 50% y sustrato CC (C50 y CC) (0,29 y 0,28%, respectivamente) (Cuadro 5). En contraste, el efecto independiente del factor B (biofertilizante) no fue significativamente diferente (p=0,8501) (Cuadro 6).
El contenido promedio de potasio fue de 1,32%. No se encontró una interacción significativa entre el tipo de sustrato y el biofertilizante (p=0,1122). El efecto independiente del factor A (sustrato) si fue significativo (p<0,05) en el cual, la no adición de materia orgánica disminuyó el contenido de potasio en las plántulas, en comparación con los tratamientos que si recibieron materia orgánica (Cuadro 5). El efecto independiente del factor B (biofertilizante), también resulto significativo (p<0,05), aunque la aplicación de Azotobacer solo y en combinación con HMA (Ac y Gi+Ac) disminuyó los contenidos de potasio vegetal, con respecto a los demás tratamientos (Cuadro 6).
Experimento 2. Efecto de la simple inoculación de Biofertilizantes y su interacción con la materia orgánica en el crecimiento, biomasa y nutrición en plántulas de papaya
La altura promedio de las plántulas que recibieron una simple biofertilización en etapa de plántula fue 22,2 cm y fue significativamente menor (p<0,05) a las que recibieron una doble biofertilización (experimento 1).
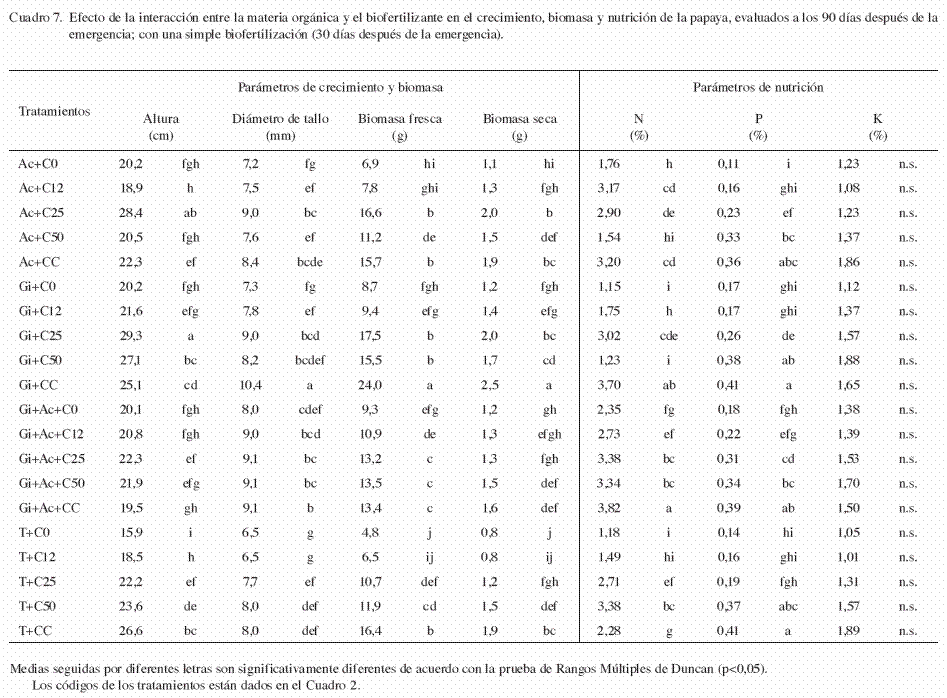
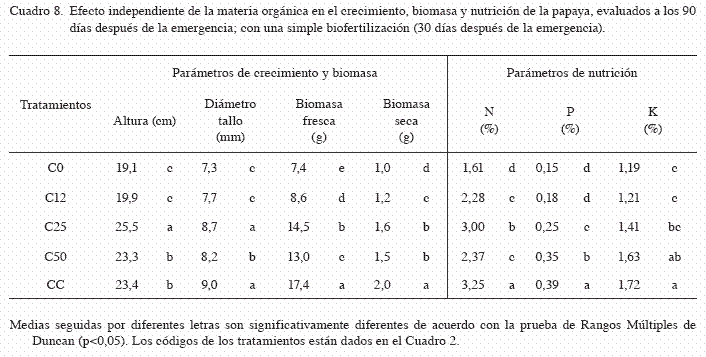
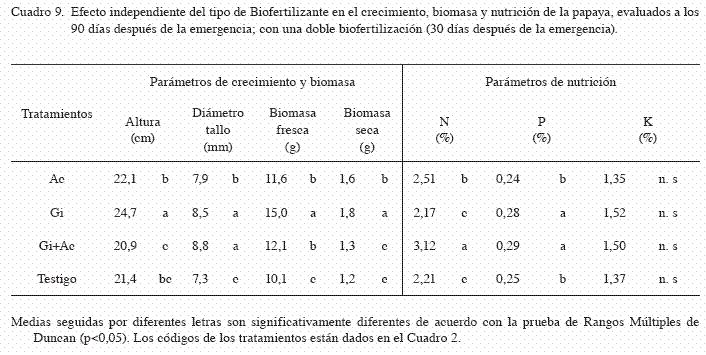
La altura de las plántulas en la simple biofertilización fue significativamente afectada por la interacción del sustrato y el biofertilizante (p<0,05). Las plántulas crecidas en el sustrato Gi+C25 y Ac+C25, fueron más altas que en los demás tratamientos (29,3 y 28,4 cm, respectivamente) (Cuadro 7). El efecto independiente del factor A (sustrato) fue significativo (p<0,05), pero el mayor efecto se debió a la aplicación de composta al 25% (25,5 cm) (Cuadro 8). El efecto independiente del factor B (biofertilizantes) también fue significativo (p<0,05), pero la aplicación independiente de HMA (Gi) registró la mayor altura con respecto a los otros tratamientos (24,7 cm) (Cuadro 9).
El diámetro promedio del tallo de las plántulas que recibieron una simple biofertilización fue 8,2 mm, y es significativamente menor (p<0,05) a las que recibieron una doble biofertilización.
En la simple biofertilización, el diámetro de tallo de las plántulas fue significativamente afectada por la interacción del sustrato y el biofertilizante (p<0,05). Las plántulas crecidas en los Gi+CC, registraron el mayor diámetro de tallo (10,4 mm) (Cuadro 7). El efecto independiente del factor A (sustrato) fue significativo (p<0,05), pero el mayor efecto se debió a la aplicación de sustrato CC y composta al 25% (9,0 y 8,7 mm, respectivamente) (Cuadro 8). El efecto independiente del factor B (biofertilizantes) fue significativo (p<0,05), donde la aplicación sola y combinada de HMA con Azotobacter (Gi y Gi+Ac), incrementaron significativamente el diámetro de tallo con respecto a los demás tratamientos (8,8 y 8,5 mm, respectivamente) (Cuadro 9).
La acumulación promedio de biomasa fresca y seca de las plántulas que recibieron una simple biofertilización fue significativamente menor a las que recibieron una doble biofertilización (p<0,05).
En la simple biofertilización, la biomasa seca y fresca de las plántulas fueron afectadas significativamente por la interacción del sustrato y los biofertilizantes. Las plántulas crecidas en los tratamientos Gi+CC, registraron la mayor biomasa en fresco y seco (biomasa fresca=23,99 g, biomasa seca=2,52 g) (Cuadro 7). El efecto independiente del factor A (sustrato) fue significativo, pero el mayor efecto en la biomasa fresca y seca se observó con la adición de sustrato CC (biomasa fresca=17,37 g y biomasa seca=1,99 g) (Cuadro 8). En el factor B (biofertilizante), la aplicación simple de HMA (Gi) registró la mayor acumulación de biomasa fresca y seca (biomasa fresca=15,0 g, biomasa seca=1,76 g) y fue significativamente mayor a los demás tratamientos (p<0,05) (Cuadro 9).
El contenido promedio de nitrógeno en las plántulas con simple biofertilización fue de 2,5%, pero no fue significativamente diferente (p=0,972) al contenido de N registrado en las plántulas con doble biofertilización.
En la biofertilización simple, el contenido de N fue significativamente afectado por la interacción del sustrato y el biofertilizante (p<0,05). Las plántulas crecidas en los tratamientos Gi+Ac+CC y Gi+CC, obtuvieron los mayores contenidos de nitrógeno en tejido vegetal (3,82 y 3,70%, respectivamente) (Cuadro 7). El efecto independiente del factor A (sustrato) fue significativo, pero el mayor efecto se debió a la adición de sustrato CC (3,24%) (Cuadro 8). Al evaluar el efecto independiente del factor B (biofertilizante), se observó que la inoculación combinada de HMA y Azotobacter (Gi+Ac), incrementaron significativamente l contenido de nitrógeno con respecto a los otros tratamientos (3,12%) (Cuadro 9).
El contenido promedio de fósforo en tejido vegetal, en las plántulas con simple iofertilización fue de 0,26% pero no es significativamente diferente al contenido de P en las plántulas con doble biofertilización (p=0,909).
En la simple biofertilización, el contenido de P fue afectado significativamente por la interacción del sustrato y el biofertilizante. Las plántulas crecidas en el tratamiento Gi+CC, registraron el mayor contenido de P (0,41%) (Cuadro 7). El efecto independiente del factor A (sustrato) fue significativo, donde el mayor efecto se debió a la adición de sustrato CC (0,40%) (Cuadro 8). En cuanto al efecto del factor B (biofertilizantes), la inoculación de HMA combinada con Azotobacter (Gi+Ac), y la inoculación sola de HMA (Gi), promovieron la mayor acumulación de P en tejido vegetal (0,29 y 0,28%, respectivamente) (Cuadro 9).
El contenido promedio de potasio en tejido vegetal en las plántulas con simple biofertilización fue de 1,43% pero no es significativamente diferente (p=0,067) al contenido de K en las plántulas con doble biofertilización.
En la simple biofertilización, el contenido de K no fue afectado significativamente por la interacción del sustrato y el biofertilizante (p=0,4837). En contraste, el efecto independiente del factor A (sustrato) fue significativo (p<0,05), pero el mayor efecto se debió a la adición de sustrato CC y composta al 50% (CC y C50) (Cuadro 8). De igual forma, el efecto del factor B (biofertilizante) no fue significativo en el contenido de potasio vegetal (p=0,3500). En los 2 experimentos (doble y simple inoculación), los mejores resultados correspondieron a la aplicación de 35% de cascarilla de cacao mas la inoculación de G. intraradices de forma independiente.
Discusión
La mayoría de las investigaciones se han dirigido a evaluar solamente una aplicación de biofertilizantes, aunque también se han realizado diversos trabajos en los que se han aplicado inoculaciones sucesivas (Albrecht et al. 1981, Millet et al. 1984, Nowak et al. 2004), pero de acuerdo con Bashan (1986), la aplicación de múltiples inoculaciones tiene un efecto mínimo en el crecimiento vegetal, debido a que muchos de los sitios de inoculación en las raíces ya han sido ocupados por las bacterias.
En contraste, los resultados aquí obtenidos demostraron que la doble inoculación incrementó el crecimiento y la biomasa vegetal en comparación con la inoculación simple de plántulas. Aunque, esto pudo deberse a que en la doble inoculación la primera aplicación de los biofertilizantes se hizo a las semillas y en la inoculación simple los biofertilizantes se aplicaron en las raíces 30 días después de la emergencia; y de acuerdo con Kalpunik et al. (1985) y Bashan (1986), la respuesta de las plantas es más alta cuando las semillas han sido inoculadas, pero es menor cuando las plántulas son inoculadas.
El efecto sinérgico de las PGPR y los HMA sobre el crecimiento vegetal, toma de nutrientes y rendimiento de varios cultivos han sido reportados (Brown y Carr 1984, Veeraswamy et al. 1992, Alarcón et al. 2002, Singh et al. 2004). Aunque, algunos efectos negativos pueden ser obtenidos (Staley et al. 1992, Alarcón et al. 2002); debido a la actividad fisiológica de la bacteria, la cual podría mostrar cierta inhibición sobre el establecimiento y la efectividad fúngica (Paulitz y Linderman 1989). La inoculación de papaya con A. chroococcum y G. intraradices en este estudio, registró un efecto sinérgico solo en algunos de los parámetros evaluados. La respuesta sinérgica del hospedero podría ser principalmente debido a la producción de fitohormonas o reguladores del crecimiento por estos microorganismos más que por pequeños incrementos en la disponibilidad en N y P (Barea et al. 1975, Vessey 2003).
La reducida respuesta de la inoculación simple de Azotobacter en el crecimiento y nutrición de papaya, estuvo influenciado por el momento de inoculación y por el tipo de sustrato. En el experimento 1, donde la rizobacteria se aplicó en semillas y 30 días después de la emergencia de las plántulas, se registró una mayor biomasa seca y contenido de fósforo en comparación al experimento 2, donde la rizobacteria fue aplicada solo en plántulas 30 días después de la emergencia (Cuadro 6). Esto se pudo deber a que la rizobacteria no fue adicionada hasta que las plántulas habían pasado la etapa más receptiva, en la que Azotobacter afecta el crecimiento vegetal (Brown y Burlingham 1968).
Estos resultados coinciden con los resultados obtenidos por Kapulnik et al. (1985), quienes encontraron que la inoculación de semillas de trigo con Azospirillum fue más eficiente que cuando se inocularon plantas, 20 días después de la emergencia, que indica además, que la rizobacteria afectó a las semillas principalmente en la germinación y menos en la etapa de colonización de raíces.
Por otra parte, el cultivo de papaya ha sido considerado como una planta dependiente de la inoculación micorrízica (Jaizme-Vega y Azcon 1995, Sukhada 1989) y de la materia orgánica (Rodríguez 1984) lo cual explicaría los efectos benéficos obtenidos en este trabajo de la interacción entre la materia orgánica y la inoculación de HMA. Resultados similares han sido encontrados en diversos estudios, en el incremento del crecimiento vegetal y biomasa fresca y seca de tallos y raíces (Khade y Rodrígues 2009a), aumento en el contenido total de potasio y fósforo en el tejido vegetal (peciolo) (Khade y Rodrígues 2009b), así como también, incrementos en la actividad enzimática de la fosfatasa acida en raíz (Alarcón 2002). Los incrementos en la biomasa fresca obtenidos en este trabajo podrían ser debido a fitohormonas como auxinas, las cuales se incrementan en plantas inoculadas con micorrizas (Barker y Tagu 2000, Fitze et al. 2005). Mientras que el aumento en la biomasa seca puede deberse a la eficiencia de los HMA en suministrar nutrientes a las plantas.
Es bien conocido que la colonización de los HMA pueden ser tanto incrementados (Gryndler et al. 2006, Albertser et al. 2006) como disminuidos (Calvet et al. 1992, Ravnskov et al. 2006) por la adición de enmiendas orgánicas. Por otra parte, de acuerdo con Osorio et al. (2002), la aplicación de enmiendas orgánicas y la inoculación de HMA incrementaron el crecimiento de plántulas de café, pero este crecimiento fue severamente disminuido cuando no se adicionaron enmiendas al suelo, presentándose bajos contenidos de materia orgánica, pH y una baja disponibilidad de P, B, Ca, Mg y K. Esto concuerda con los resultados encontrados en este estudio, ya que las plántulas que crecieron en los sustratos con adición de materia orgánica y fueron inoculadas con G. intraradices, mejoraron los parámetros de crecimiento, biomasa y nutrición en comparación cuando no se adicionó materia orgánica al sustrato. Pero el mejor tratamiento fue aquel en el que se adicionó un 35% de cascarilla de cacao como materia orgánica, lo cual pudo deberse a que este sustrato presentó un contenido moderado de fósforo (66,04 mg.kg-1) en relación con los demás sustratos a los que también se les adicionaron materia orgánica (Cuadro 1). De acuerdo con Kahiluoto et al. (2000), incrementos en el suministro de P disminuyen la colonización y la efectividad de la colonización micorrízica. Debido a los beneficios en la traslocación de nutrientes que realizan los HMA, en diversas ocasiones a niveles moderados de fósforo se han obtenido mejoras sustanciales en la nutrición de las plantas (Thompson 1987, Yao et al. 2001).
Por otra parte, la aplicación de composta al 12% y sustrato con cascarilla de cacao, estimularon el mayor contenido de nitrógeno en tejido vegetal, lo cual puede deberse a que estos sustratos presentaron una relación C/N menor de 20 (Cuadro 1), y de acuerdo con Day y Shaw (2001) los tratamientos con relaciones C/N<10 permiten una mayor liberación de nutrimentos que aquellos con valores >20, pues se considera que la relación C/N ha sido utilizada como un índice de la velocidad de descomposición de los abonos y la posterior mineralización de los nutrientes.
Según Bianciotto y Bonfante (2002), el establecimiento de la inoculación de PGPR en la rizósfera puede ser afectado por la co-inoculación de HMA. Los resultados aquí obtenidos demostraron que la población de A. chroococcum no se vio afectada por la aplicación combinada con G. intraradices (Cuadro 3), pero si por la materia orgánica adicionada. También se ha observado que la respuesta de la planta a la inoculación con HMA dependerá del nivel de fertilidad del suelo, de la planta hospedera y del hongo (Sharda y Rodrígues 2008). Asimismo, conforme a los porcentajes de colonización micorrízica encontrados y con los resultados obtenidos en el crecimiento y en la nutrición de las plantas, puede establecerse que el beneficio de la simbiosis se expresó aún en presencia de dosis elevadas de P.
Conclusión
La doble biofertilización esto es, en semilla y plántula, estimuló el crecimiento y biomasa de las plántulas en comparación a la simple biofertilización. Asimismo, la mejor opción para iniciar la biofertilización en el cultivo de la papaya es la inoculación directa a las semillas con A. Chroococcum y G. Intraradices. La adición combinada de cascarilla de cacao al 35% más la inoculación de G. intraradices, registró la mejor interacción sobre el crecimiento y la biomasa de las plántulas de papaya, aunque, la incorporación de composta al 25% con G. intraradices, también puede ser empleada como un tratamiento alternativo cuando no se disponga de cascarilla de cacao. En las condiciones establecidas en este trabajo, se encontró un efecto sinérgico entre los factores de crecimiento y contenidos de fósforo en planta.
Literatura citada
Alarcón A., Davis F.T. Jr., Egila J.N., Fox t.C., Estrada-Luna A.A., Ferrera-Cerrato R. 2002. Short term effects of Glomus claroideum and Azospirillum brasilense on growth and root acid phosphatase activity of Carica papaya L. under phosphorus stress. Rev. Latinoam. Microbiol. 44:31-37. [ Links ]
Albertser A., Ravnskov S., Green H., Jensen D.F., Larsen J. 2006. Interactions between the external mycelium of the mycorrhizal fungus Glomus intraradices and other soil microorganisms as affected by organic matter. Soil Biol Biochem 38:1008-1014. [ Links ]
Albrecht S.L., Okon Y., Lonnquist J., Burris R.H. 1981. Nitrogen fixation by corn-Azospirillum associations in a temperate climate. Crop Science 21:301-306. [ Links ]
Barea J.M, Azcon R., Hayman D.S. 1975. Possible synergistic interactions between Endogyne and phosphate-solubilizing bacteria in low phosphate soils, pp. 409-417. In: F.E. Sanders, B. Mose, P.B. Tinker PB (eds). Endomycorrhizas. Academic Press,
London. [ Links ]
Barker S.J., Tagu D. 2000. The roles of auxinas and cytokinins in mycorrhizal symbioses. Journal of Plant Growth Regulation.19:144-154. [ Links ]
Bashan Y. 1986. Significance of timing and level of inoculation with rhizosphere bacteria of inoculation with rhizosphere bacteria on wheat plants. Soil Biol. Biochem. 18:297-301. [ Links ]
Brown M.E., Burlinghams K. 1968. Production of plant growth substances by Azotobacter chroococcum. Journal of General Microbiology 53, 135-144. [ Links ]
Brown M.E., Carr G.R. 1984. Interactions between Azotobacter chroococcum and vesicular-arbuscular mycorrhiza and their effects on plant growth. Journal of Applied Bacteriology. 56:429-437. [ Links ]
Calvet C., Estaun V., Camprubi A. 1992. Germination, early mycorrhizal growth and infectivity of a vesicular–arbusular mycorrhizal fungus in organic substrates. Symbiosis 14:405-411. [ Links ]
Chen J.H. 2006. The combined use of chemical and organic fertilizer and/or biofertilizer for crops growth and soil fertility. International Workshop on Sustained Management of the Soil-Rhizosphere System for Effcicient Crop Production and Fetrtilizer Use. Land Developmente Department Bangkok, Thailand. [ Links ]
Cupull S.R., Guerra P.G., Cupull S.M.C., Ferrer V.M., Pérez N.C. 2002. Efecto de Trichoderma viride y Azotobacter chroococcum en la estimulación de la germinación y el desarrollo de posturas de Carica papaya Lin. Centro Agrícola 4:30-33. [ Links ]
Day M., Shaw K. 2001. Biological, chemical and physical processes of composting, pp. 18-22. In: P. Stofella y B. Kahn (eds). Compost utilization in horticultural cropping systems. Lewis, U.S.A. [ Links ]
Fitze D., Wiepning A., Kaldorf M., Ludwigmüller J. 2005. Auxins in the development of an arbuscular mycorrhizal symbiosis in maize. J Plant Physiol. 162:1210-1219. [ Links ]
Gryndler M., Larsen J., Hršelová H., Řezáčová V., Gryndlerová H., Kubát J. 2006. Organic and mineral fertilization, respectively, increase and decrease the development of external mycelium of arbuscular mycorrhizal fungi in a long-term field experiment. Mycorrhiza 16:159-166. [ Links ]
Hegde S.V., Brahmaprakash G.P. 1992. A dry granular inoculant of Rhizobium for soil application. Plant Soil 144:309-311. [ Links ]
Jaizme-Vega M.C., Azcon R. 1995. Responses of some tropical and subtropical cultures to endomycorrhizal fungi. Mycorrhiza 5:213-217. [ Links ]
Kahiluoto H., Ketoja E., Vestberg M. 2000. Promotion of utilization of arbuscular mycorrhiza through reduced fertilization 1. Biossays in a growth chamber: Plant and soil. 227:191-206. [ Links ]
Kapulnik Y., Gafny R., Okon Y. 1985. Effect of Azospirillum spp. inoculation on root development and NO,- uptake in wheat (Triticum aestivum cv. Miriam) in hydroponic systems. Can. J. Bot. 63:627- 631. [ Links ]
Khade S.W., Rodrigues B.F. 2009b. Studies on effects of arbuscular Mycorrhizal (Am.) Fungi on mineral Nutrition of Carica papaya L. Not. Bot. Hort. Agrobot. Cluj. 37:183-186. [ Links ]
Khade W.S., Rodrigues B.F. 2009a. Studies on arbuscular mycorrhization of papaya. African Crop Science Journal. 17:155-165. [ Links ]
Mesa R.J.R., Gómez C.J.L., Rodríguez C.O., Parets S.E., Soto O.R. 2006. Efecto de Trichoderma y micorrizas en la producción de posturas de Carica papaya L. Centro Agrícola. 3:75-81. [ Links ]
Millet E., Avivi Y., Feldman M. 1984. Yield response of various wheat genotypes to inoculation with Azospirillum brasilense. Plant and Soil 80:261-266. [ Links ]
Mokwunye A.U., De Jager A., Smaling E.M.A. 1996. Restoring and Maintaining the Productivity of West African Soil: Key to sustainable Development. (eds.) In: Miscellaneous Fertilizer Studies Nº. 14. [ Links ]
Nowak J., Nowak J., Sharma V.K., A’Hearn E. 2004. Endophyte enhancement of transplant performance in tomato, cucumber and sweet pepper. Acta Hort. (ISHS) 631:253-263. [ Links ]
Osorio N.W., Alzate J.M., Ramirez G.A. 2002. Coffee seedling growth as affected by mycorrhizal inoculation and organic amendment. Communications in Soil Science and Plant Analysis 33:1425-1434. [ Links ]
Paulitz T.C., Linderman R.G. 1989. Interactions between fluorescent pseudomonas and VA mycorrhizal fungi. New Phytol. 113:37-45. [ Links ]
Ravnskov S., Jensen B., Knudsen I.M.B., Bodker L., Jensen D.F., Karlinski L., Larsen J. 2006. Soil inoculation with the biocontrol agent Clonostachys rosea and the mycorrhizal fungus Glomus intraradices results in mutual inhibition, plant growth promotion and alteration of soil microbial communities. Soil Biol Biochem 38:3453–3462. [ Links ]
Rodríguez H. 1984. Nutrición en frutales. Psidium guajaba y Carica papaya. Boletín de reseñas. Cítricos y otros frutales. CIDA. MINAGRI. Cuba. pp. 27-51. [ Links ]
Sharma A., Parmar D.K., Kumar P., Singh Y., Sharma R.P. 2008. Azotobacter soil amendment integrated with cow manure reduces need for NPK fertilizers in Sprouting Broccoli. Internal Journal of vegetable Science. 14:273-285. [ Links ]
Singh R., Behl R.K., Singh K.P., Jain P., Narula N. 2004. Performance and gene effects for wheat yield under inoculation of arbuscular mycorrhiza fungi and Azotobacter chroococcum. Plant and Soil Environ. 50:409-415. [ Links ]
Staley T.E., Lawrence E.G., Nance E.L. 1992. Influence of a plant growth-promoting pseudomonas and vesicular-arbuscular mycorrhizal fungus on alfalfa and birdsfoot trefoil growth and nodulation. Biol Fertil Soils 14:175-180. [ Links ]
Sukhada M. 1989. Response of papaya (Carica papaya L.) to VAM fungal inoculation. In: Mycorrhizae for Green Asia (A. Mahadevan, N. Raman and K. Natarajan) Madras, Alamer Printing works. Royapettah, Madras. [ Links ]
Thompson J.P. 1987. Decline of vesicular-arbuscular mycorrhizae in Long Fallow Disorder of field crops and its expression in phosphorus deficiency of sunflower. Australian Journal of Agriculture Research. 38:847-867. [ Links ]
Valdés M., Reza-Alemán F., Furlan V. 1993. Response of Leucaena esculenta to endomycorrhizae and Rhizobium inoculation. World J Microbiol Biotechnol. 9:97-99. [ Links ]
Veeraswamy J., Padmavathi T., Venkateswarlu K. 1992. Interaction effects of Glomus intraradices and Azospirillum lipoferum on sorghum. Indian J. Microbiol. 32:305-308. [ Links ]
Verma R.K., Jamaluddin, Dadwal V.S., Thakur A.K. 2008. Economics of biofertilizer application on production of planting propagules of teak in a commercial nursery. Indian Forester. 134:923-929. [ Links ]
Vessey J.K. 2003. Plant growth promoting rhizobacteria as biofertilizers. Plant and Soil 255:571-586. [ Links ]
Wasy A.A., Shyamalamma S., Nache G.V. 2010. Effect of bio-inoculants on nursery establishment of papaya cv. Solo. Acta Hort. (ISHS) 851:389-394. [ Links ]
Waterer D.R., Coltman R.R. 1988. Phosphorus concentration and application interval influence growth and mycorrhizal infection of tomato and onion transplants. J. Am. Soc. Hortic. Sci. 113:704- 708. [ Links ]
Yao Q., Li X.L., Feng G., Christie P. 2001. Influence of extrametrical hyphae on mycorrhizal dependency of wheat genotypes. Communications in soil Science and Plant Analysis.32:3307-3317. [ Links ]
Correspondencia a: Regino Gómez. El Colegio de la Frontera Sur. Carretera Villahermosa-Reforma km 15,5. Ranchería Guineo, sección II. CP 86280. Villahermosa, Tabasco. México. / Correo electrónico: regomez@ecosur.mx
Maricela Constantino. El Colegio de la Frontera Sur. Carretera Villahermosa-Reforma km 15,5. Ranchería Guineo, sección II. CP 86280. Villahermosa, Tabasco. México.
José David Álvarez. El Colegio de la Frontera Sur. Carretera Villahermosa-Reforma km 15,5. Ranchería Guineo, sección II. CP 86280. Villahermosa, Tabasco. México.
Juan Manuel Pat. El Colegio de la Frontera Sur. Carretera Villahermosa-Reforma km 15,5. Ranchería Guineo, sección II. CP 86280. Villahermosa, Tabasco. México.
Elda Guadalupe Espín. Instituto de Biotecnología. Universidad Autónoma de México. Av. Universidad Número 2001, Col. Chamilpa C.P. 62210. Cuernavaca, Morelos C.P. 62250. México.
Recibido: 30/08/10 Aceptado: 07/03/11

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}