










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgronomía Costarricense
Print version ISSN 0377-9424
Agron. Costarricense vol.34 n.1 San Pedro de Montes de Oca Jun. 2010
Nota técnica
Comunidad de arvenses en un cultivo de maní (Arachis hypogaea) y su relación con la rata Sigmodon hirsutus en Alajuela,
Costa Rica
Costa Rica
Javier Monge1/*, Allan Chavarría**, Esteban Duverrán**
1/Correo electrónico: javier.monge@ucr.ac.cr
* Centro de Investigaciones en Protección de Cultivos (CIPROC), Escuela de Agronomía, Universidad de Costa Rica. San José, Costa Rica.
** Escuela de Agronomía, Universidad de Costa Rica. San José, Costa Rica.
Dirección para correspondecia
Resumen
Se caracterizó la comunidad de arvenses en un ciclo de producción de maní (Arachis hypogaea) y se relacionó con las capturas de la rata de campo (Sigmodon hirsutus). Se realizaron 4 muestreos mensuales de arvenses; se evaluó la riqueza, diversidad, cobertura, altura y densidad por parcela, estado fenológico y biomasa. Para el muestreo de roedores se usó una cuadrícula con 40 trampas distanciadas a 10 m entre sí, durante 2 noches consecutivas por mes. La comunidad de arvenses estuvo constituida por 22 especies, a través del ciclo de producción de maní. Ageratum conyzoides estuvo presente en todos los muestreos y fue la especie dominante de la comunidad de arvenses. Las especies de hoja angosta estuvieron presentes a través de todo el ciclo de producción, con una mayor representatividad en el primer muestreo, aunque con una menor riqueza que la mostrada en el último muestreo. Se reconoce la ocupación de S. hirsutus en un agroecosistema dominado por especies de hoja ancha, lo cual evidencia su adaptabilidad a diferentes hábitats, ya que es una especie común en agroecosistemas dominados por especies de hoja angosta.
Palabras clave: Comunidad de arvenses, diversidad, maní, Arachis hypogaea, rata de campo, Sigmodon hirsutus.
Abstract
Community of weeds in a groundnut crop (Arachis hypogaea) and its relationship to the rat Sigmodon hirsutus in Alajuela, Costa Rica. We have characterized the weed community during a production cycle of peanut (Arachis hypogaea) and related it to the catch of the field rat (Sigmodon hirsutus). Four monthly samples of weeds were done, assessing the richness, diversity, coverage, height and plot density, biomass and phenological state. For the sampling of rodents, a grid with 40 traps spaced 10 m apart was used, during 2 consecutive nights per month. The weed community was composed of 22 species, through the cycle of peanut production. Ageratum conyzoides was present in all samples and was the dominant species of the weed community. Narrow-leaf species were present throughout the production cycle, with greater representation in the first sampling, although with less richness than the one shown in the last sampling. The occupation of S. hirsutus of an agroecosystem dominated by broadleaf species is recognized, which demonstrates its adaptability to different habitats, as it is a common species in agroecosystems dominated by narrow-leaf species.
Keywords: Weed community, diversity, peanut, Arachis hypogaea, field rat, Sigmodon hirsutus.
Introducción
El componente vegetal de un agroecosistema está constituido por el cultivo y las arvenses que se encuentran dentro o en los alrededores del área de producción. Entre el cultivo y las arvenses se establecen diferentes relaciones que les afectan directa e indirectamente. Es así, como a las arvenses se le ha llamado malezas, para hacer referencia a la interferencia ejercida al cultivo, manifestado por la competencia por nutrientes, agua y luz, recursos que cuando son escasos comprometen el adecuado desarrollo de las plantas, así como el efecto de alelopatía hacia los cultivos (Doll 1996).
También existen efectos indirectos, cuando las arvenses establecen relaciones con otros organismos y con el suelo, que pueden favorecer o perjudicar al cultivo. En la medida que las arvenses le proporcionen alimento y refugio a especies que perjudican al cultivo, se considera que afectan negativa e indirectamente al cultivo; sin embargo, las arvenses pueden ser comparables con los cultivos de cobertura al reconocer los beneficios potenciales de estos (Gliessman 2002). Por lo tanto, la relación entre el cultivo y las arvenses no puede considerarse solo positiva o negativa, sino que existe un continuo entre estos 2 extremos, manifestado de diversas maneras y en intensidades diferentes.
Para aprovechar los beneficios que aportan las arvenses a los cultivos y evitar los efectos negativos, es necesario conocer la vegetación acompañante de los cultivos, y determinar las diversas relaciones que se establecen entre sí, así como aquellas con otros organismos que habitan en el agroecosistema. Aún cuando los actuales sistemas de producción agrícola han sido simplificados, (Altieri 1992) es evidente la complejidad implícita en esta trama de relaciones ecológicas que se establecen entre los subsistemas de un agroecosistema, según lo esquematizado por Hart (1985), por lo que solo un estudio integral permitiría adquirir un conocimiento completo de lo que ocurre en un ecosistema. Sin embargo, esta misma complejidad, dificulta el desarrollo de un estudio integral, por lo que para fines prácticos se fragmenta la realidad, para poder establecer relaciones particulares entre unos pocos componentes del sistema, teniendo claro que es una visión parcial del funcionamiento del agroecosistema.
Uno de los componentes de agroecosistema lo representan las especies vertebradas, dentro de las que se encuentran aves, mamíferos, anfibios y reptiles. En el agroecosistema con maní se encuentran representantes de esas clases de vertebrados, que incluyen roedores y en particular la rata de campo Sigmodon hirsutus (Monge 2008a). Esta especie fue recientemente reconocida como especie nueva (Peppers et al. 2002 y Carroll y Bradley 2005), se encuentra en Centroamérica desde Nicaragua hasta Panamá y al norte de Colombia y Venezuela (Musser y Carleton 2005). Su ecología es poco conocida (Monge 2008b), y se le considera una especie plaga para la producción agrícola y silvícola en Centroamérica (Monge 2009).
En el presente caso, se propone caracterizar la comunidad de arvenses, en términos de riqueza y dominancia de especies, así como de diversidad, en períodos mensuales durante el ciclo de producción de un cultivo de maní y relacionarlo con los sitios de captura de la rata de campo (Sigmodon hirsutus).
Materiales y métodos
El estudio se realizó en la Estación Experimental Agrícola Fabio Baudrit Moreno (EEAFBM) de la Universidad de Costa Rica (UCR) ubicada en La Garita, Alajuela, Costa Rica, en las coordenadas geográficas 10º01' Latitud Norte y 84º16' Longitud oeste, a una altitud de 840 msnm; durante los meses de setiembre a diciembre de 2007. El sitio se caracteriza climáticamente por una temperatura promedio anual de 23,3ºC, 6,6 horas de brillo solar promedio, una precipitación anual de 1745,4 mm y una humedad relativa de 79%. Existen 2 estaciones climáticas, la lluviosa entre junio a noviembre y la seca entre diciembre y mayo. El lote de estudio tuvo una extensión de 4800 m2 (120 x 40 m), mientras que el área de muestreo fue de 4000 m2 (100 x 40 m).
El lote estaba bordeado por franjas de vegetación arvense que la separaban de otras actividades agrícolas, además de la existencia de calles de aproximadamente 5 m de ancho y distancias de más de 10 m de otros cultivos. El cultivo se estableció el 8 de agosto de 2007 y se utilizó la variedad de maní americano. El sistema de siembra fue en lomillos, separados a 0,8 m entre sí, y a 30 cm entre plantas. En cuanto a fertilización se utilizó 200 kg.ha-1 de 10-30-10 en el momento de la siembra, además se aplicó terbufós al fondo de los huecos de siembra. En cuanto al control de malezas, se usó herbicidas en el momento de la siembra en preemergencia para el maní y pre-postemprano a las malezas. Los herbicidas utilizados fueron: pendimetalina 1,5 kg ia.ha-1 + imazetapir 14 g ia.ha-1 + 450 de paraquat + Dash al 0,4%. Se incluyó paraquat en la mezcla para eliminar las malezas ya emergidas antes de que saliera el maní. Un mes después se aplicó imazetapir 14 g ia.ha-1 + fluzifop butil 120 g ia.ha-1. El volumen de aplicación fue de 200.l.ha-1.
Cada mes, durante el ciclo de producción de maní, se establecieron 24 parcelas de 0,25 m2 (50 x 50 cm) distribuidas al azar en el área de producción. En cada parcela se determinó las especies de arvenses presentes y para aquellas cuya identificación no fuera segura en el campo, se consultó la guía de hierbas y arbustos comunes en cafetales y otros cultivos (Nilsson et al. 2005), así como a especialistas del Programa de Malezas de la EEAFBM y del Laboratorio de Arvenses de la Escuela de Agronomía de la UCR. De cada especie se determinó la cobertura, la altura de las plantas y la cantidad de individuos por parcela. Luego se colectó todas las especies de arvenses presentes en las parcelas y se determinó el peso fresco por especie, en el laboratorio. Dada la alteración provocada por el muestreo, los puntos utilizados se excluían de muestreos posteriores. Para caracterizar y comparar la comunidad de arvenses entre muestreos mensuales se usó como referencia la riqueza, la cobertura por parcela, la altura, densidad, estado fenológico y biomasa en peso fresco por especie. Para estimar la diversidad de cada muestreo se usó el índice de Shannon (H) (Krebs 1985), cuya fórmula de cálculo es H=-∑pi log pi, donde pi representa la proporción de cada especie en la población (Somarriba 1999).
Para el muestreo de roedores se establecieron 40 estaciones fijas de muestreo, en forma de cuadrícula, en 4 filas con 10 trampas cada una, con distanciamiento entre trampas de 10 m. El muestreo consistió en trampeos mensuales de 2 noches consecutivas, utilizándose trampas de golpe (Marca Victor®) grandes (17,5 x 8,5 cm). Se utilizó como cebo, la mezcla de avena, maíz quebrado y banano maduro, en proporciones similares. Para cada captura se registraba la fecha y el sitio en donde esta ocurrió. Los individuos fueron clasificados por sexo y edad. Para la edad se utilizó como referencia el peso corporal, considerándose jóvenes a los individuos con peso interior a 60 g, adulto joven de 60 a 113 g y adulto viejo los que tuvieron un peso superior a 113 g (Monge 2008a).
Para caracterizar el hábitat de los roedores, se tomó como referencia aquellos puntos de muestreo de arvenses cercanos a sitios de captura de roedores que estuvieran a una distancia no mayor de 10 m y que fueran muestreados en el mismo mes de realizada la captura.
Resultados y discusión
Comunidad de arvenses en un cultivo de maní
La comunidad de arvenses durante el ciclo de producción de maní estuvo constituida por 22 especies, de las cuales 13 son de hoja ancha de las familias Asteraceae (7), Rubiaceae (3), Commelinaceae (1), Euphorbiaceae (1) y Lamiaceae (1), y 9 de hoja angosta de las familias Poaceae (8) y Cyperaceae (1).
Dos especies, Melampodium divaricatum y Ageratum conyzoides (conocida popularmente como Santa Lucía) estuvieron presentes en los 4 muestreos, cuya tendencia de frecuencia difirió entre sí, ya que, la de M. divaricatum disminuyó durante el período de estudio, mientras que la de A. conizoides fue creciente. Otras especies como Cyperus rotundus (coyolillo), Cynodon dactylon (zacate de Bermuda) e Ixophorus unisetus (zacate de Honduras) estuvieron presentes en 3 de los muestreos. Aún cuando ninguna especie de hoja angosta en particular, estuvo presente en todos los muestreos, en ellos siempre hubo presencia de este tipo de vegetación, con una mayor representación en el muestreo de diciembre.
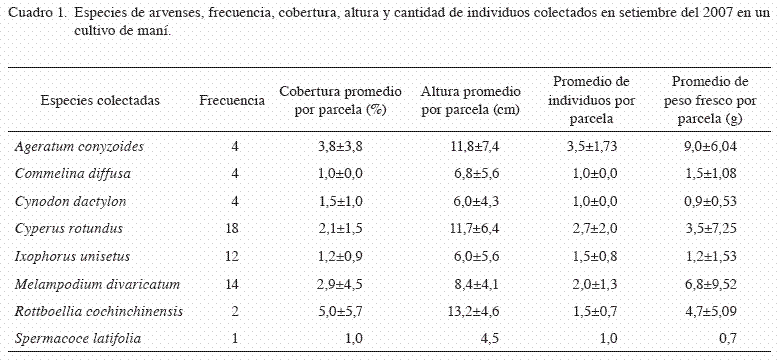
En el primer muestreo realizado en setiembre, a un mes de establecido el cultivo, la comunidad de arvenses estuvo representado por 8 especies. Tres especies fueron las dominantes: C. rotundus, M. divaricatum y I. unisetus, con frecuencias de aparición en 18, 14 y 12 parcelas, respectivamente, del total de 24 parcelas (Cuadro 1). En cuanto a la cantidad de especies de arvenses por parcela osciló entre 1 y 4 especies, con un promedio de 2,5±0,9. El conocimiento de la riqueza se logró en un 75% (6 especies) con las primeras 6 parcelas, y el 100% (7 especies) con 20 parcelas. El índice de diversidad de Shannon tuvo un valor de 2,31, el cual es inferior a lo esperable para ecosistemas naturales, cuyo valor oscila entre 3 y 4 (Gliessmann 2002).
El área de cobertura por parcela de las diferentes especies osciló entre 1 y 16%, donde Rottboellia cochinchinensis, A. conyzoides y presentaron mayor cobertura promedio. El porcentaje de cobertura promedio por parcela para todas las arvenses fue de 5,4±4,9, con respecto al área ocupada por el cultivo de maní o por área sin vegetación. La altura promedio de las plantas osciló entre 2 y 21,5 cm, donde las especies M. divaricatumR. cochinchinensis, A. conyzoides y C. rotundus mostraron una mayor altura promedio. En cuanto a la cantidad promedio de individuos por especie por parcela fue entre 1 y 3,5, para A. conyzoides y C. rotundus, que resultaron ser las más abundantes por parcela.
Para la biomasa, en términos de peso fresco por especie por parcela, se obtuvieron valores entre 0,9 y 9,0 g, para A. conyzoides y M. divaricatum, especies que mostraron pesos promedio superiores, por parcela.
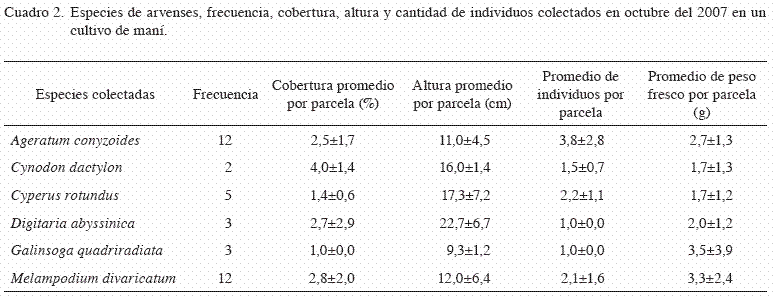
Para el segundo mes (octubre) se redujo la riqueza a 6 especies, donde M. divaricatum mantuvo el predominio; aumentó A. conizoides; y se redujo la de C. rotundus, con frecuencias de aparición en 12 parcelas para las 2 primeras y en 5 para la última (Cuadro 2). La cantidad de especies por parcela fue de 1 y 3, con un promedio de 1,5±1,1. La riqueza de especies fue conocida en un 83,3% (5 especies) en las primeras 6 parcelas, y el 100% con 9 parcelas. Para este mes la diversidad bajó con respecto al mes anterior, pues se obtuvo un valor de 1,87. Este índice es influenciado por la riqueza de especies, así como la proporción de cada especie (Pla 2006), por lo que su valor aumenta a medida que aumenta la riqueza y los individuos estén representados más homogéneamente (Somarriba 1999). Esto explica en este caso, la reducción del índice de diversidad, ya que la riqueza del segundo mes fue de un 75% con respecto al anterior, y la equidad entre las especies tiende a disminuir al aumentar la proporción de la especie predominante.
El área de cobertura por especie por parcela fue entre 1 y 6%, donde C. dactylon presentó mayor cobertura promedio, aunque no se diferenció significativamente de las otras especies. En cuanto al porcentaje de cobertura por parcela, que incluyó todas las especies, aumentó a 7,5±6,9, sin que difiera significativamente de lo observado en el mes anterior. Con respecto a las alturas de las plantas, en este segundo muestreo se observó un incremento con valores de 7 a 27 cm, para Digitaria abyssinica (alambrillo) y C. rotundus con mayor altura promedio. La cantidad promedio de individuos de cada especie por parcela se mantuvo y en este caso entre 1 y 3,8, donde A. conyzoides mostró mayor densidad.
Para la biomasa, en términos de peso fresco por especie y por parcela, los valores promedio oscilaron entre 1,7 y 3,5 g, sin que exista una diferencia significativa entre especies.
Para el tercer mes de muestreo (noviembre) la comunidad de arvenses estuvo representada por 6 especies, al igual que en el mes anterior, pero con diferencias en su composición. En este muestreo aparecen Acmella oppositifolia y Richardia scabra (chiquizacillo), aunque con poca representación, y reaparece I. unisetus (Cuadro 3). A. conyzoides muestra un predominio con la frecuencia de aparición en 18 parcelas, seguido de Galinsoga quadriradiata (mielcilla) con 10, donde ambas muestran un incremento con respecto al mes anterior. La cantidad de especies por parcela osciló entre 1 y 3 con un promedio de 1,7±0,8. El 67% de la riqueza de especies fue conocida con las primeras 3 parcelas y el 100% con 17 parcelas. El índice de diversidad continuó su tendencia a la baja en este mes, con 1,52, de una especie (A. conyzoides) claramente dominante.
El área de cobertura por especie por parcela osciló entre 1 y 10% con una mayor cobertura promedio de A. oppositifolia, aunque no difiere de las otras especies, mientras que para todas las especies de arvenses la cobertura promedio por parcela fue de 6,2±3,3. La altura promedio osciló entre 12 y 45 cm, donde A. conyzoides e I. unisetus alcanzaron una mayor altura promedio. La cantidad de individuos por especie y por parcela osciló entre 1 y 4,4, y ubica a A. conyzoides con mayor densidad por parcela, aunque no difiere significativamente de las otras 2 especies.
Con respecto a la biomasa por especie y parcela, esta osciló entre 1,7 y 16,8 g, lo cual aumentó considerablemente el ámbito observado en el mes anterior. La especie con mayor peso promedio fue A. conyzoides, seguido de A.oppositifolia, sin que existan diferencias significativas entre especies.
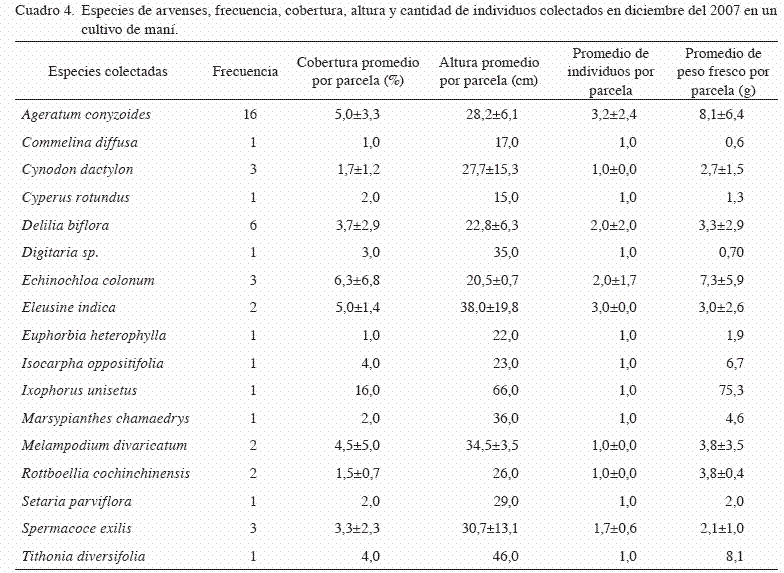
En el muestreo de diciembre, la riqueza de arvenses aumentó considerablemente, encontrándose 17 especies con un claro predominio de A. conyzoides, que se identificó en 16 de las 24 parcelas, seguida de Delilia biflora (lentejas), con una frecuencia de aparición en 6 parcelas mientras que el resto de especies con una frecuencia de 3 o menos (Cuadro 4). Para ese mes, el índice de diversidad se incrementó y superó todos los valores anteriores ya que fue de 2,57 por mes. Este valor ha sido influenciado principalmente por la riqueza de especies que constituyeron la comunidad de arvenses en este mes, además que se observa una mayor equidad entre las especies, a pesar de que una de ellas sigue mostrando un claro predominio.
La cantidad de especies por parcela fue de 1,9±1,1 y no evidenció diferencia significativa entre muestreos durante el ciclo de producción de maní. Dada la riqueza de especies presentes en este mes, se requirió de una mayor cantidad de parcelas con respecto a los meses anteriores, ya que el 70% de la riqueza se obtuvo con 11 parcelas, y 20 parcelas para encontrar el 100% de las especies presentes en el área de estudio.
El área de cobertura por especie por parcela osciló entre 1 y 16%, con una mayor cobertura promedio mostrada por Echinocloa colonum, seguida por A. conyzoides y Eleusine indica (pata de gallina), sin que existan diferencias significativas con respecto al resto de las especies. Para todas arvenses, la cobertura promedio aumentó con respecto a los meses anteriores a 9,0 7,4.
En ese mes, la altura de las plantas osciló entre 15 y 66 cm, donde I. unisetus presentó mayor tamaño, seguido de E. indica y M. divaricatum. La cantidad promedio de individuos por parcela para las diferentes osciló entre 1 y 3,25, con una mayor densidad A. conyzoides y E. indica.
Con respecto a la biomasa, durante este mes osciló entre 1,3 y 75,3 g por especie y parcela, aunque con excepción de I. unisetus, las otras especies se mantuvieron en un ámbito de 1,3 a 8,1 g.
Al considerar las prácticas de manejo dadas al cultivo, se evidencia la influencia de este en la comunidad de arvenses, en donde el primer mes tuvo una riqueza relativamente baja y una diversidad intermedia, lo cual indica que las labores relacionadas con el control inicial de malezas permitieron la presencia de pocas especies, en donde C. rutundus y M. divaricatum lograron superar el efecto de estas labores. Para los siguientes 2 meses, la diversidad se redujo, con predominio de 2 y 3 especies de hoja ancha, o sea el mismo tipo de vegetación del cultivo, lo cual es indicativo de una limitante de aplicar herbicidas para este tipo de vegetación, dado el posible efecto negativo sobre el cultivo. Finalmente, para diciembre se evidencia un aumento considerable de la riqueza de arvenses, lo cual puede ser explicado por una suspensión en las labores relacionadas con el control de malezas a partir del segundo mes, ya que para esa época se ha superado la etapa crítica para el cultivo, período estimado en los primeros 40 días (Anónimo 1991). A su vez, diciembre corresponde a la época seca, lo cual puede influir en forma importante en el desarrollo de una comunidad de arvenses diferente al de la época lluviosa.
Con respecto a la fenología de las especies de arvenses durante el ciclo de producción de maní, se tiene que durante el primer muestreo (setiembre) la mayoría se encontraban en estado vegetativo con excepción de A. conizoides que estaba en floración, especie relativamente poco frecuente en ese mes. Para octubre, la mayoría de las especies tenían plantas tanto en estado vegetativo como en floración, de las cuales las más frecuentes (A. conizoides y M. divaricatum) tenían plantas en floración en un 25% de las parcelas en donde se les encontró. En noviembre y diciembre, el estado de floración fue el predominante, lo cual se relaciona con un próximo período de producción de semillas que pueden servir de alimento a los roedores, aunque los roedores también pueden aprovechar otras partes de diferentes especies de arvenses, como ha sido probado para la especie congénere S. hispidus (Fleharty y Olson 1969, Kincaid y Cameron 1982).
Monitoreo de roedores en un cultivo de maní
Durante el período de estudio se capturaron 11 individuos de la especie S. hirsutus, lo cual se logró en los 2 últimos meses de un total de 4. En noviembre se capturaron 3 hembras: 2 jóvenes y una adulta joven, y 3 machos: uno joven, uno adulto y un adulto viejo.
En diciembre se capturaron 5 individuos, 2 hembras, 1 joven y 1 adulta vieja, la cual estaba preñada; 2 machos, 1 adulto y 1 adulto viejo, y un individuo joven que no se contabilizó pues no pudo ser sexado, dado el deteriorado estado de sus genitales por efecto de las hormigas.
En noviembre las capturas se realizaron a una distancia de 10 m del borde. Para diciembre, las capturas se concentraron en el borde del sector suroeste, con solo una captura a 10 m del borde este.
De las 6 capturas realizadas en noviembre, 5 de ellas fueron en parcelas cercanas al muestreo de arvenses. Una captura realizada en el borde norte tuvo 5 parcelas cercanas de muestreo de arvenses, en las cuales estuvo presente A. conyzoides, con una área de cobertura de 5% o menos, y una altura entre 23 y 38 cm. En esas parcelas también se encontró otra especie de arvense, pero su presencia varió entre las 5 parcelas, de 1 a 2% de cobertura y una altura de 23 a 32 cm.
En la segunda captura, también cercana al borde norte, habían 3 parcelas cercanas de muestreo de arvenses y en ellas fue común la presencia de A. conyzoides. En este caso, esta especie logró alcanzar en una de las parcelas una cobertura del 10% y las alturas oscilaron entre 25 y 32 cm.
Finalmente, en la última captura localizada cerca del costado sur, en la parcela de muestreo cercana se identificaron también 3 especies de arvenses, además de A. conizoides con una altura de 45 cm y una cobertura del 5%, la cual superó a las otras 2 especies, G. quadriradiata y A. oppositifolia, que presentaron una altura de 15 cm y una cobertura de 1 y 2%.
Respecto a las 5 capturas de diciembre, 4 de ellas se hallaron en parcelas cercanas del muestreo de arvenses. Dos capturas de este mes se realizaron en la misma estación de muestreo, ubicada al centro en dirección norte a sur y en el borde este del lote de estudio. En la parcela cercana se identificaron las arvenses Tithonia diversifolia (mirasol) y E. colonum, con alturas de 46 y 20 cm, respectivamente. Otras capturas también ubicadas en el borde este, y del centro hacia el sur, se dieron donde habían 3 especies de arvenses, siendo D. biflora común para ambos sitios de captura, acompañadas de A. conyzoides y C. dactylon, así como M. divaricatum y C. diffusa para la otra parcela.
En términos generales, es notoria la presencia de A. conyzoides en las parcelas de muestreo cercanas a los sitios de captura de roedores; sin embargo, se debe resaltar que esta ha sido la especie de arvense más común en el lote de estudio, predominando en los meses en que hubo captura de roedores. Dada la presencia en 3 parcelas de otras especies de arvenses en varias parcelas cercanas a sitios de muestreo en noviembre, tales como G. quadriradiata, presente en 4 de las 11 parcelas cercanas al sitio de captura de roedores y M. divaricatum se hizo difícil establecer una relación entre los sitios de captura de roedores y la presencia de otra especie de arvense, aparte de A. conizoides.
En diciembre no hubo una tendencia clara, ya que se identificaron otras arvenses (Cuadro 4) con poca coincidencia en las parcelas de muestreo cercanas a los sitios de captura, donde es común D. biflora y A. conizoides que se presentaron en 2 de 4 parcelas.
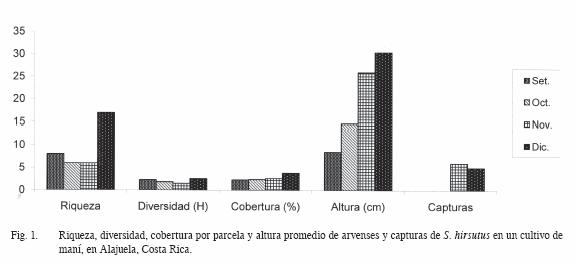
Al considerar los criterios usados para describir la comunidad de arvenses y su relación con la cantidad de capturas en los diferentes muestreos, la riqueza de especies parece no haber influido en forma considerable. La cantidad de capturas entre noviembre y diciembre no difirió en forma importante (Figura 1). Respecto a la diversidad, se utilizó como referencia el Índice de Shannon. La diversidad varía muy poco, lo cual no logró explicar el comportamiento de las capturas de roedores, excepto en los 2 últimos meses. En cuanto a la cobertura porcentual de arvenses por parcela, el promedio no alcanzó el 5% del área de la parcela, tampoco explica la cantidad y el momento de las capturas realizadas. En cuanto a la altura de las plantas, si parece existir una mayor relación, ya que en noviembre y diciembre, se muestra una vegetación de mayor altura, lo cual podría relacionarse con un mejor hábitat para este roedor. Sin embargo, debe considerarse que el cultivo para estos meses también muestra una mayor cobertura, lo que puede propiciar condiciones que justifiquen una mayor presencia de roedores dentro del área de producción de maní.
Respecto a la representatividad de las especies de hoja ancha y de hoja angosta en los meses que hubo captura de roedores, se observó un evidente dominio de especies de hoja ancha. Para diciembre, hay una mayor equidad en términos de riqueza para cada tipo de vegetación, cuando se encuentra 9 especies de hoja ancha y 8 de hoja angosta; presentan además superioridad en términos de individuos en una relación de 3,6:1 (hoja ancha:hoja angosta). Estas características del hábitat nos indicaron que la rata de campo (Sigmodon hirsutus) se adapta a diferentes ambientes, como el utilizado en este caso con un predominio de especies vegetales de hoja ancha, máxime que el cultivo es de este tipo de vegetación, mientras que también se le encuentra en agroecosistemas dominados por especies de hoja angosta, como caña de azúcar, arroz y maíz (Monge 2009), así como en hábitat naturales que incluyen diferentes tipos de bosque y pastizales (Bonoff y Janzen 1980). Otras especies del mismo género se relacionan con hábitats dominados por pastos o con presencia de estos, como el caso de S. hispidus (Cameron y Spencer 1981), S. fulviventer (Baker y Shump 1978a), S. ochrognathus (Baker y Shump 1978b) y S. leucotis (Shump y Baker 1978). Sin embargo, el predominio de especies de hoja angosta al inicio del ciclo de producción (Cuadro 1) no fue determinante para su ocupación por parte de los roedores.
En este sentido, se puede considerar que esta especie se adapta a las condiciones de una variedad amplia de ambientes y a su vez, posiblemente se comportaría como una especie oportunista que aprovecha los recursos de cada hábitat, pero no solo de disponibilidad de alimento, sino también de sitios de refugio, lo cual puede explicar su ausencia en los primeros muestreos. A su vez, dada que la disponibilidad de vainas de maní hasta el último mes, sugiere que la presencia inicial de este roedor obedece a la protección que le puede suministrar el desarrollo de la vegetación representada tanto por el cultivo como por la comunidad de arvenses. De igual manera, aunque no se ha determinado en este estudio, es posible que además de protección, algunas especies de arvenses le proporcionen algún tipo de alimento.
Conclusiones
La comunidad de arvenses varió a través del ciclo de producción del agroecosistema con maní (A. hypogaea), con predominio de especies de hoja ancha a partir del segundo mes, posiblemente influenciada por las labores agrícolas, en particular las relacionadas con el control de malezas. La especie A. conizoides (Santa Lucía) mostró dominancia en la comunidad de arvenses del agroecosistema con maní, y eventualmente representó un elemento importante en el hábitat ocupado por la rata de campo (S. hirsutus). La mayor altura de las arvenses junto con la cobertura alcanzada por el cultivo de maní, proporcionó un hábitat más adecuado para los roedores y de ahí el por qué de su presencia en la segunda mitad del ciclo de producción. En este sentido, se puede considerar que existe una relación beneficiosa de la comunidad de arvenses para los roedores dentro de este agroecosistema. La rata de campo (S. hirsutus) mostró adaptabilidad al ambiente dominado por especies de hoja ancha, aunque es una especie de roedor común en agroecosistemas dominados por especies de hoja angosta.
Agradecimientos
Los autores agradecen al Programa de Malezas de la EEAFBM por permitir realizar el estudio en sus áreas de producción, así como al Ing. Steven Brenes y al Dr. Franklin Herrera por la colaboración en la identificación de arvenses y al M.Sc. Rodolfo WingChing por sus comentarios al manuscrito. El estudio fue financiado por la Vicerrectoría Investigación de la Universidad de Costa Rica, a través del proyecto 813-A6-171.
Literatura citada
Anónimo 1991. Maní (Arachis hypogaea L.). In: Ministerio de Agricultura y Ganadería. 1991. Aspectos técnicos sobre cuarenta y cinco cultivos agrícolas de Costa Rica. Dirección General de Investigación y Extensión Agrícola. San José, Costa Rica. Consultado el 16 set. 2008. Disponible en http://www.mag.go.cr/ bibioteca_virtual_ciencia/tec-mani.pdf. [ Links ]
Altieri M.A. 1992. El rol ecológico de la biodiversidad en agroecosistemas. Agroecología y desarrollo 4. Consultado el 16 set. 2008. Disponible en línea http://www.clades.cl/revistas/4/rev4art1.htm. [ Links ]
Baker R.H., Shump. 1978a. Sigmodon fulviventer. Mammalian Species 94:1-4. [ Links ]
Baker R.H., Shump. 1978b. Sigmodon ochrognathus. Mammalian Species 97:1-2. [ Links ]
Bonoff M.B., Janzen D.H. 1980. Small terrestrial rodents in eleven habitats in Santa Rosa National Park, Costa Rica. Brenesia 17:163-174. [ Links ]
Cameron G.N., Spencer S.R. 1981. Sigmodon hispidus. Mammalian Species 158:1-9. [ Links ]
Carroll D.S., Bradley R.D. 2005. Systematics of the genus Sigmodon: DNA sequences from betafibrinogen and cytochrome b. The Southwestern Naturalist 50(3):342-349. [ Links ]
Doll J.D. 1996. Dinámica y complejidad de la competencia de malezas. In: R. Labrada, J.C. Caseley y C. Parker. Manejo de malezas para países en desarrollo. Estudio FAO Producción y Protección Vegetal-120. Organización de las naciones Unidas para la Agricultura y la Alimentación. Roma, Italia. En línea: http://www.fao.org/docrep/T1147S/t1147s00. HTM. [ Links ]
Fleharty E.D., Olson L.E. 1969. Summer food habits of Microtus ochrogaster and Sigmodon hispidus. J. Mamm. 50(3):475-486. [ Links ]
Gliessman S.R. 2002. Agroecología: procesos ecológicos en la agricultura sostenible. Centro Agronómico Tropical de Investigación y Enseñanza (CATIE). Turrialba, Costa Rica. 359 p. [ Links ]
Hart R.D. 1985. Agroecosistemas. Conceptos básicos. Centro Agronómico Tropical de Investigación y Enseñanza (CATIE). Turrialba, Costa Rica. 159 p. [ Links ]
Kincaid W.B., Cameron G.N. 1982. Dietary variation in three sympatric rodents on the Texas coastal prairie. J. Mamm. 63(4):668-672. [ Links ]
Krebs C.J. 1985. Ecología. Estudio de la distribución y la abundancia. Segunda edición. Harper y Row Latinoamericana. México, D. f. México. 753 p. [ Links ]
Monge J. 2008a. Estructura poblacional y actividad reproductiva de la rata de campo (Sigmodon hirsutus) durante un ciclo de producción de maní (Arachis hypogaea) en Costa Rica. Agronomía Costarricense 32(2):161-167. [ Links ]
Monge J. 2008b. Estado del conocimiento sobre la rata de campo (Sigmodon hirsutus) en Costa Rica. Manejo Integrado de Plagas y Agroecología 79-80:1-6. [ Links ]
Monge J. 2009. Roedores plaga de América Central. Editorial de la Universidad de Costa Rica y Organismo Internacional Regional de Sanidad Agropecuaria (OIRSA). San José, Costa Rica. 146 p. [ Links ]
Musser G.G., Carleton M.D. 2005. Superfamily muroidea. In: Wilson, and D.M. Reeder (eds). Mammals species of the world. A taxonomic and geographic reference. Third edition. The Johns Hopkins University Press. Baltimore, USA. 2142 p. [ Links ]
Nilsson V., Sánchez-Vindas P., Abarca R.M. 2005. Hierbas y arbustos comunes en cafetales y otros cultivos. Herbario Juvenal Valerio, Universidad Nacional. Heredia, Costa Rica. 246 p. [ Links ]
Peppers L.L., Carroll D.S., Bradley R.A. 2002. Molecular systematics of the genus Sigmodon (Rodentia:Muridae): evidence from the mitochondrial cytochrome-b gene. J. Mamm. 83(2):396-407. [ Links ]
PLA L. 2006. Biodiversidad: Inferencia basada en el índice de Shannon y la riqueza. Interciencia 31(8):583- 590. [ Links ]
Somarriba E. 1999. Diversidad Shannon. Agroforestería en las Américas 6(23):72-74. [ Links ]
Shump K.A., Baker. R.H. 1978. Sigmodon leucotis. Mammalian Species 96:1-2. [ Links ]
Correspondeica a: Javier Monge. Centro de Investigaciones en Protección de Cultivos (CIPROC), Escuela de Agronomía, Universidad de Costa Rica. San José, Costa Rica. Correo electrónico: javier.monge@ucr.ac.cr
Allan Chavarría. Escuela de Agronomía, Universidad de Costa Rica. San José, Costa Rica.
Esteban Duverrán. Escuela de Agronomía, Universidad de Costa Rica. San José, Costa Rica.
Recibido: 11/09/09 Aceptado: 13/02/10

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}