










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgronomía Costarricense
Print version ISSN 0377-9424
Agron. Costarricense vol.34 n.1 San Pedro de Montes de Oca Jun. 2010
Efecto de la aplicación de abonos orgánicos en la supresión de Pythium myriotylum en plantas de tiquisque (Xanthosoma sagittifolium)
1/ Correo electrónico: lidieth.uribe@ucr.ac.cr
* Instituto Tecnológico de Costa Rica. Cartago, Costa Rica.
** Centro de Investigaciones Agronómicas, Universidad de Costa Rica. San José, Costa Rica.
*** Centro de Investigación en Protección de Cultivos, Universidad de Costa Rica. San José, Costa Rica.
Dirección para correspondencia
Resumen
Los abonos orgánicos ejercen efecto supresivo sobre patógenos de planta; su capacidad supresora varía de acuerdo al tipo de abono y al sistema planta-patógeno. Se evaluó el efecto supresor de diferentes abonos orgánicos en el sistema tiquisque-Pythium myriotylum. Se determinó la madurez, estabilidad, y actividad microbiana de los diferentes abonos, así como el efecto del tipo de compostaje (compost vs vermicompost), el material de origen (estiércol vs broza de café) y el grado de madurez, sobre la supresión del patógeno. El efecto supresivo sobre el desarrollo de la enfermedad en plantas de tiquisque se estableció por medio de una escala visual de síntomas a los 3, 6 y 9 días del transplante. Los abonos presentaron relaciones C/N entre 6 y 15 y ninguno afectó la germinación o la longitud de las raíces de plántulas de pepino. Los abonos maduros clasificaron como estables y los inmaduros, con excepción del compost de broza, como inestables. La menor incidencia de pudrición de raíces por P. myriotylum se obtuvo con el uso de vermicompost a base de estiércol maduro, mientras que la menor severidad de la enfermedad ocurrió cuando el suelo se enmendó con el compost y el vermicompost maduro a base de estiércol. En términos generales los abonos obtenidos a partir de estiércol fueron más supresivos, y presentaron una mayor actividad microbiana, que los producidos a base de broza de café. Se concluye que el tipo de compostaje, el origen y el grado de madurez tienen influencia sobre la capacidad supresora.
Palabras clave: Compost, vermicompost, Pythium myriotylum, abonos orgánicos, tiquisque, Xanthosoma sagittifolium.
Abstract
Effect of the application of compost and vermicompost on the suppression of Pythium myriotylum in cocoyam plants (Xanthosoma sagittifolium). Organic amendments have been used as a substrate which is able to suppress soil-borne plant pathogens. Since these effects vary depending on the type of compost and the plant-pathogen system, the aim of this work was to evaluate the suppressive effect of different organic fertilizers against cocoyam root rot disease, caused by Pythium myriotylum. For this purpose, the effect of the composting process (compost vs. vermicompost), origin (animal manure vs. coffee fruit-peel pulp) and degree of maturity were tested. Stability and microbial activity of the organic amendment were also determined. The suppressive effect over disease development on cocoyam was established by using a visual severity-scale on days 3, 6, and 9 after transplant. The organic fertilizers presented C/N ratios between 6 and 15. Neither affected the germination or root-length of cucumber seedlings. The mature organic fertilizers classified as stable; and the immature ones, with the exception of the coffee-pulp compost, as unstable. The treatment with the best result in reducing root rot disease incidence was the mature, vermicomposted animal manure. The lowest disease severity was obtained when the soil was amended with the mature, composted or vermicomposted, animal manure. In general, the organic amendments based on animal manure were more suppressive than the ones based on coffee pulp. These results indicate that the type of composting, original material and maturity influence the suppressivity of the organic amendments.
Keywords: Compost, vermicompost, Pythium myriotylum, organic amendments cocoyam, Xanthosoma sagittifolium.
Introducción
El compost y otros tipos de abonos orgánicos aplicados al suelo tienen el potencial de promover el control biológico de enfermedades de plantas (Hoitink y Grebus 1994, Craft y Nelson 1996, Hoitink y Boehm 2001, Noble y Coventry 2005, Termorshuizen et al. 2006, Danon et al. 2007). Estos abonos pueden introducir agentes de biocontrol al suelo y proporcionar alimento para su establecimiento y actividad (Hoitink y Grebus 1994, Termorshuizen et al. 2006, Danon et al. 2007); pueden mejorar la condición de la raíz y aportar nutrientes a la planta, lo que favorece un crecimiento adecuado del cultivo que le permita tolerar las enfermedades o escapar de la infección (Huber 1980, Huber 1991). Esta característica de los abonos orgánicos para impedir el desarrollo de enfermedades en las plantas se conoce como supresividad. La supresión biológica por medio del compost involucra mecanismos de antagonismo directo como la competencia, la antibiosis y el hiperparasitismo, así como mecanismos no antagónicos tales como la resistencia sistémica inducida en las plantas (Hoitink et al. 1997, Lievens et al. 2001, Diánez et al. 2007, Suárez-Estrella et al. 2007, Malandraki et al. 2008). Con base en estos mecanismos existen 2 tipos de supresividad: la general, debida a la alta diversidad de microorganismos presentes en el abono que crea condiciones desfavorables para el desarrollo de la enfermedad, y la específica, en la cual uno o pocos microorganismos son responsables del control de un determinado patógeno (Hoitink et al. 1997).
Los abonos orgánicos mejor conocidos son el compost y el vermicompost. En el proceso de compostaje acontece la transformación microbiana de la materia orgánica bajo condiciones controladas: en los primeros días ocurre un ligero incremento de la temperatura que va de 20 a 45ºC, producto de la descomposición de azúcares (fase mesofílica), que puede alcanzar posteriormente temperaturas de 55 a 70ºC (fase termofílica) durante la degradación de la celulosa, en la que ocurre la disminución de la población microbiana. Una vez transcurrida ésta fase se da inicio al proceso de maduración del compost, donde al disminuir la temperatura, ocurre la recolonización por microorganismos que pueden ser antagónicos a organismos fitopatógenos (Bollen 1993, Paul y Clark 1996, Hoitink et al. 1997, Atlas y Bartha 2002, Scheuerell y Mahaffee 2005, Diánez et al. 2007). En esta fase, los remanentes orgánicos son degradados a una tasa más lenta (Lazcano et al. 2008).
Por su parte, el vermicompostaje involucra la bio-oxidación y estabilización del material orgánico por la acción de lombrices que aerean, acondicionan y fragmentan el sustrato y por microorganismos que degradan bioquímicamente la materia orgánica. En este proceso se distingue una fase activa donde las lombrices procesan el desecho y modifican su estado físico, composición microbiana y una fase similar a la de maduración, marcada por el desplazamiento de las lombrices hacia capas más frescas del desecho (Domínguez 1997, Lazcano et al. 2008).
La estabilidad y la madurez del compost han sido estudiadas como factores influyentes en la supresividad de enfermedades (Hoitink y Grebus 1994, Hoitink et al. 1997, Suarez-Estrella et al. 2007). La estabilidad está relacionada con el grado de descomposición de la materia orgánica y puede ser expresada como una función de la actividad microbiana en el compost, que se evalúa por pruebas respirométricas (Chen y Inbar 1993, Wu et al. 2000, Adani et al. 2006). La madurez se refiere al grado de descomposición de los compuestos fitotóxicos orgánicos producidos durante la fase de composteo y se evalúa a través de bioensayos con plantas indicadoras (Wu et al. 2000, CCREF 2001, Gómez Brandon et al. 2008).
El efecto supresor de los abonos sobre patógenos de plantas varía de acuerdo al tipo de abono y al sistema planta-patógeno (Termorshuizen et al. 2006). Encontraron en estudios realizados a partir de 120 bioensayos, que involucran 18 tipos de compost y 7 patosistemas, que en el 54% de los casos hubo supresión significativa de la enfermedad, mientras que solo en el 3% ocurrió un aumento; y que los patógenos fueron afectados en forma diferente por los compost. Por otro lado Bonanomi et al. (2007) indican, que la capacidad de las enmiendas orgánicas para suprimir la enfermedad varió con el patógeno estudiado, observándose supresión en más del 50% de los casos de Verticillium, Thielaviopsis, Fusarium y Phytophthora, mientras que para Rhizoctonia solani la supresión fue alcanzada solo en el 26% de los casos. Estos resultados indican que existe gran potencial para el uso de abonos orgánicos, por lo que hay que realizar mayor investigación sobre los mecanismos que actúan en la supresión de enfermedades.
La supresividad se evalúa normalmente al adicionar el abono orgánico a un sustrato inoculado con el patógeno de interés y en el cual se cultiva una planta hospedera. Es conveniente además, evaluar el compost en un sustrato no inoculado, para estudiar su efecto directo sobre el crecimiento de la planta (Erhart y Burian 1997).
El cultivo del tiquisque (Xanthosoma sagittifolium (L.) Schoot & Endl.), que pertenece a la familia de las aráceas (Araceae) y se cultiva en regiones tropicales al oeste de África, América Central y el Caribe, Oceanía y Asia (Tambong 2001, Perneel et al. 2006), es afectado por el patógeno Pythium myriotylum var. araceanum que causa la enfermedad conocida como mal seco (Pacumbaba et al. 1992, Perneel et al. 2006, Saborío 2007). Este organismo causa pudrición de raíces y en la mayoría de las plantas termina por destruir completamente el sistema radical. Como síntoma indirecto de la destrucción de las raíces ocurre el marchitamientom en el área foliar, que se inicia por las hojas más viejas. En plantas adultas se producen periódicamente raíces, lo que mantiene la planta viva permitiendo el brote de algunas hojas nuevas (Laguna et al. 1983, Gómez 1993, Arauz 1998, Perneel et al. 2006). El efecto supresor de abonos orgánicos sobre Pythium ultimum (Schüler et al. 1989, Garcia et al. 2004, Scheuerell y Mahaffee 2005, Veeken et al. 2005), P. graminícola (Craft y Nelson 1996) P. aphanidermatum (Zmora-Nahum et al. 2008), Pythium sp (Diánez et al. 2007) ha sido documentado. Existe sin embargo poca información del efecto de abonos orgánicos sobre el sistema Tiquisque- Pythium myriotylum. Adiobo (2006) encontró una incidencia significativamente menor de la enfermedad causada por P. myriotylum en tiquisque, en 3 suelos que fueron enmendados con compost que en el suelo sin enmendar; sin embargo, el nivel de supresión fue satisfactorio solo en uno de los sitios estudiados.
En este trabajo se analiza el uso de abonos orgánicos de diferentes orígenes y en distintos grados del proceso de preparación, en el control del daño ocasionado por P. myriotylum var. Aracearum en plantas de tiquisque.
Materiales y métodos
La investigación se realizó en el Centro de Investigaciones Agronómicas (CIA) de la Universidad de Costa Rica.
Establecimiento del inóculo
Se utilizó el hongo Pythium myriotylum var. aracearum proveniente de una planta de tiquisque con síntomas de daño por mal seco. El hongo se cultivó en placas con medio Parc (Singleton et al. 1992) y se subcultivó en agar agua (Somasegaran y Hoben 1985). Posteriormente se inoculó en arroz a fin de obtener un sustrato para la subsecuente inoculación del hongo en el suelo. Para esto se utilizaron erlenmeyers de 125 ml con 26 g de arroz y 14 ml de agua, que se autoclavaron 2 veces, en días consecutivos, durante 25 min a 121ºC. Cada erlenmeyer se inoculó con 4 secciones (5 mm de diámetro) de agar y dentro el cultivo puro de P. myriotylum. Estos cultivos se incubaron a 25ºC en oscuridad durante una semana, agitando el frasco cada 2 días y hasta que los granos estuviesen colonizados.
Material Vegetal Se utilizaron plantas provenientes de cultivo in vitro con 12 semanas de aclimatización.
Abonos Los abonos fueron recolectados a partir de diferentes productores. Se utilizó compost a base de estiércol de cabra, vermicompost a partir de estiércol bovino, compost y vermicompost a partir de broza de café. Los abonos se colectaron para su uso en su etapa final de compostaje (abono maduro) y cuando el proceso estaba incompleto (abono inmaduro).
Madurez y estabilidad del abono
Para evaluar la madurez de los abonos, a una muestra compuesta de cada bono se le midió la relación carbono nitrógeno (C/N), y el efecto del abono sobre la germinación y crecimiento radicular de plantas de pepino (CCREF 2001). La relación C/N se determinó de acuerdo a las técnicas de Bremner y Tabatabai (1971) y Walkley y Black (1938). El efecto de los abonos sobre las plantas de pepino se evaluó a través del uso de extractos del abono. Los extractos se prepararon con 240 ml de cada compost y 150 ml de agua destilada; las mezclas se agitaron por 2 h en un agitador rotatorio y se centrifugaron a 2000 rpm durante 30 min. El sobrenadante se filtró a través de un filtro Wattman Nº 2 y el filtrado se utilizó para el bioensayo. Los extractos se evaluaron por triplicado; se adicionaron 3 ml de cada extracto en una placa de petri en la que se colocaron previamente 10 semillas de pepino sobre un papel de filtro. Como control se utilizó agua destilada. Las placas se incubaron en la oscuridad a temperatura ambiente. Se midió el porcentaje de germinación y la longitud de la radícula al día 4. La estabilidad de los abonos se evaluó por medio de la determinación de la evolución de CO2 (CCREF 2001) y la respiración, por el método de Anderson (1982).
Evaluación de la supresividad de diferentes abonos sobre Pythium myriotylum en plántulas de tiquisque
Se establecieron los siguientes tratamientos:
1. Suelo + Pythium (Control)
2. Suelo + Compost broza inmaduro + Pythium (CBI)
3. Suelo + Compost broza maduro + Pythium (CBM)
4. Suelo + Compost estiércol de cabra inmaduro + Pythium (CEI)
5. Suelo + Compost estiércol de cabra maduro + Pythium (CEM)
6. Suelo + Vermicompost broza inmaduro + Pythium (VBI)
7. Suelo + Vermicompost broza maduro + Pythium (VBM)
8. Suelo + Vermicompost estiércol bovino inmaduro + Pythium (VEI)
9. Suelo + Vermicompost estiércol bovino maduro + Pythium (VEM)
Para evaluar el efecto de los abonos sobre el crecimiento de la planta sana se implementaron además los mismos tratamientos sin la inoculación con Pythium myriotylum. Se midió el incremento en la altura de la planta después de 13 días de aplicados los tratamientos.
El suelo utilizado provenía de la zona productora de tiquisque en la localidad de Muelle de San Carlos, Costa Rica. Este se desinfectó 2 veces, en días consecutivos, mediante el autoclavado a 120ºC durante 1 h. Posteriormente se mezcló con cada uno de los abonos en una proporción de (90:10 %v/v) suelo: abono, se distribuyó en macetas de 650 ml de capacidad y se incubó por 4 días antes de la siembra de las plantas. En cada maceta se transplantaron 4 plantas de 3 meses de haber sido aclimatadas. Siete días después del transplante (ddt), se inoculó el suelo con el patógeno en los tratamientos correspondientes. En cada pote se distribuyó 20 granos de arroz colonizado con P. myriotylum, colocados en 5 puntos alrededor de las plantas. Se observó y evaluó el avance de la enfermedad y se utilizó como base, la presencia de síntomas en las plantas, mediante la escala visual de severidad establecida en la Figura 1, la cual se diseño específicamente para el ensayo. Se determinó la incidencia y la severidad como se describe a continuación.
Incidencia=Número de plantas que presentan algún grado de lesión por repetición.
Índice de severidad=∑nigi/N, donde: ni=número de plantas en la categoría i, gi es el grado de la categoría i. N=número total de plantas evaluadas por repetición.
Reaislamiento del patógeno
Para comprobar la presencia del patógeno, 15 días después del transplante, se colocaron en agar agua secciones de raíz extraídas a partir de plantas que presentaron síntomas de la enfermedad. Los platos se incubaron y se observó el crecimiento del patógeno sobre el medio de cultivo y al microscopio (Figura 2), donde se identificaron las formas y estructuras características de P. myriotylum.
Análisis estadísticos
Para las variables germinación y longitud de la radícula de pepino, se usó un diseño completamente al azar con 3 repeticiones y se realizó análisis de varianza a las variables utilizadas. La comparación de medias se hizo en todos los casos utilizando LSD Fisher (p=0,05). Para el análisis del efecto de los diferentes abonos sobre la supresión de Pythium se utilizó un diseño irrestricto al azar con 9 tratamientos y 4 repeticiones por tratamiento; cada repetición constó de 4 plantas. A las variables incidencia y severidad se les realizó un análisis de varianza a los 3, 6 y 9 días. Para la separación de medias se utilizó LSD Fisher (p=0,05).
Resultados y discusión
Madurez, estabilidad y actividad microbiana de los abonos
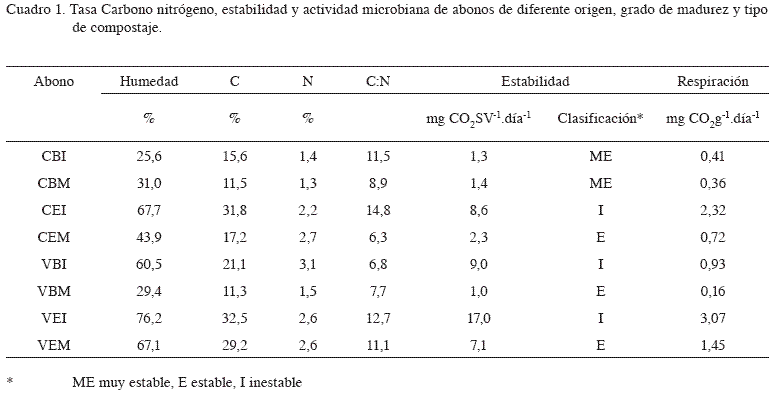
Al comparar el contenido de carbono en los distintos abonos, se encontró que este fue mayor en los abonos inmaduros que en los abonos maduros (Cuadro 1), lo cuál se debe a que durante la bioxidación de los productos ocurre la pérdida de CO2 (Paul y Clark 1996, Lazcano et al. 2008), principalmente por la disminución de compuestos carbonados fácilmente degradables (Gómez-Brandón et al. 2008). El contenido de N, por el contrario, fue similar entre abonos maduros e inmaduros del mismo origen, con excepción del vermicompost de broza; esta similitud se debe a que durante el proceso de compostaje, el N se pierde a una tasa más baja que el carbono (Gómez-Brandón et al. 2008, Lazcano et al. 2008). Los abonos a base de estiércol tuvieron contenidos de N más altos que los abonos a base de broza (con excepción del VBI), ya que el estiércol es un material naturalmente rico en N (Zmora-Nahum et al. 2007).
Todos los abonos estudiados presentaron una tasa C/N menor de 20, considerada apropiada para los abonos orgánicos (Paul and Clark 1996), ya que altas tasas C/N causan la inmovilización de N (Chen and Inbar 1993, Paul and Clark 1996). En este trabajo, la tasa C/N fue mayor en los compost inmaduros que en los compost maduros; cuando el desecho orgánico es composteado, la tasa C:N generalmente decrece y luego se estabiliza en el rango de 10 a 15 (Chefetz et al. 1996, Paul y Clark 1996, Lazcano et al. 2008). Si bien la disminución en la relación C/N es un indicador útil de la madurez del abono, debe utilizarse con precaución y para cada compost por separado, ya que varía mucho de un compost a otro (Ouatmane et al. 2000). Esto es particularmente importante en estiércoles en los que la relación C/N puede estar dentro de valores considerados como maduros a pesar de que el proceso se encuentre en las etapas iniciales de composteo (Gómez-Brandon et al. 2008). En el caso del vermicompost, las lombrices modifican el estatus químico y físico de los residuos orgánicos, lo que reduce gradualmente la tasa C/N (Dominguez et al. 1997).
Como se observa en el Cuadro 1, los abonos maduros clasificaron como estables y los inmaduros, con excepción del compost de broza inmaduro, clasificaron como inestables. El compost a base de broza inmaduro presentó además una actividad microbiana muy baja, que puede deberse al bajo contenido de humedad presente en la muestra de abono, o a que el abono haya sido recolectado en un período tardío del proceso de maduración ya que según los análisis químicos, se trata de un abono cuyas características corresponden más a un abono maduro (Cuadro 1).
Cuando se evaluó el efecto de los extractos de los diferentes abonos sobre la germinación y la longitud de la radícula de plantas de pepino, no se observó diferencias significativas en el porcentaje de germinación (97,8%±2,4) y la longitud de la radícula (7,1 cm±0,5), lo que indica que los abonos no tuvieron un efecto fitotóxico sobre las plantas.
Evaluación de la supresividad de diferentes abonos sobre Pythium myriotylum en invernadero
Las plantas de tiquisque que no fueron inoculadas con P. myriotylum no presentaron síntomas de la enfermedad durante los 9 días de evaluación; tanto el suelo autoclavado como los abonos utilizados, se encontraban libres de P. myriotylum. No se observó tampoco efecto fitotóxico sobre las plantas de tiquisque (Cuadro 2), lo que concuerda con los resultados obtenidos en el bioensayo con plantas de pepino. Si bien no hubo diferencias entre los abonos aplicados con respecto al control, las plantas a las que se les adicionó compost de estiércol inmaduro (CEI) presentaron un incremento en la altura significativamente mayor que las plantas a las que se aplicó compost a base de broza, compost y vermicompost de estiércol maduro (Cuadro 2). Este efecto puede deberse a la mayor cantidad de C en este abono inmaduro, así como a una mayor descomposición reflejada en la medida de estabilidad del abono y que podría traducirse en un mayor aporte de nutrientes.

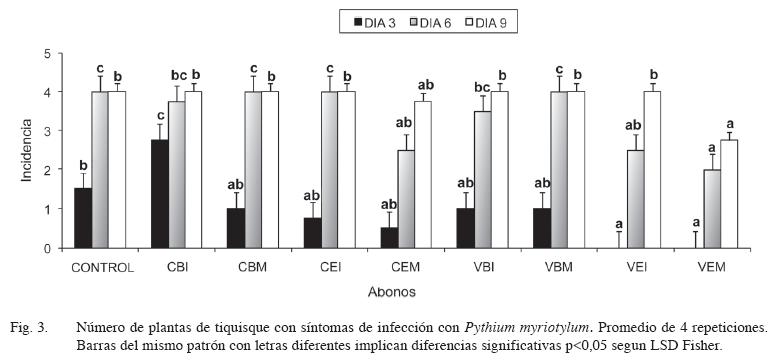
Las plantas cultivadas en suelo sin abono y luego inoculadas con el patógeno (tratamiento control), empezaron a mostrar síntomas a partir del tercer día del transplante (Figura 3) y la enfermedad avanzó rápidamente hasta producir la muerte de todas las plantas en el transcurso de una semana, lo que indica que P. miriotylium presentó un alto potencial de patogenicidad sobre las planas de tiquisque y que el suelo autoclavado utilizado en el experimento, no presentó factores evidentes de supresividad sobre el patógeno.
Los resultados indican que a los 3 días después de la inoculación (ddi), únicamente los 2 tratamientos de vermicompost a base de estiércol (VEI y VEM) no presentaron plantas con síntomas de la enfermad y la incidencia fue significativamente menor que el control (Figura 3). El tratamiento CBI presentó una mayor incidencia de la enfermedad que los tratamientos restantes. A los 6 ddi, los tratamientos CEM, VEI y VEM tuvieron significativamente menos plantas enfermas que el tratamiento control, mientras que a los 9 días solamente el tratamiento VEM presentó una menor incidencia que el control.
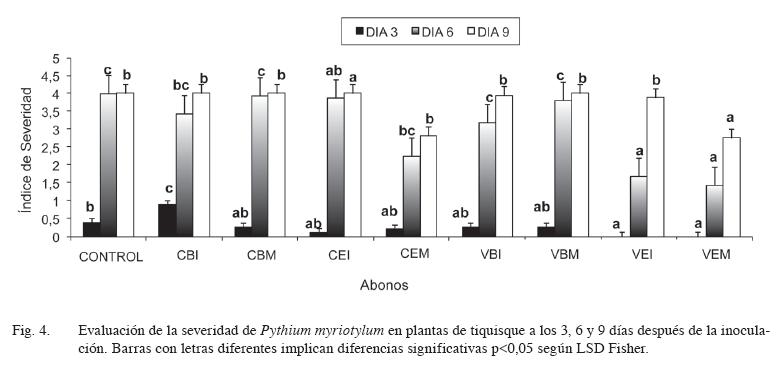
En cuanto a la severidad, en los tratamientos enmendados con vermicompost de estiércol (VEI, VEM), la severidad fue significativamente menor que en el control; de manera contrastante, la adición de compost de broza inmaduro (CBI) causó, a los 3 días de la inoculación, una severidad significativamente mayor que dichos tratamientos (Figura 4). A los 6 días, el tratamiento correspondiente al compost de estiércol maduro (CEM) y los vermicompost a base de estiércol tanto maduro (VEM) como inmaduro (VEI), fueron los que presentaron menor índice de severidad. Al día 9 solamente los tratamientos VEM y CEM presentaron una severidad significativamente menor que el tratamiento control.
En términos generales, los abonos a base de estiércol fueron más supresivos para la enfermedad causada por P. myriotylum, que los abonos a base de broza de café, lo que podría deberse a que los abonos de estiércol presentaron una mayor actividad microbiana que los abonos a base de broza; así, su adición al suelo podría crear un ambiente de alta actividad microbiana, donde los microorganismos compiten por nutrientes y/o producen antibióticos que reducen la sobrevivencia y crecimiento del patógeno (Hoitink et al. 1997, Diánez et al. 2007). El uso de abonos orgánicos provee, en este trabajo, un inóculo microbiano que puede aportar un factor supresivo al suelo autoclavado e inoculado con Pythium. Este tipo de supresión, debida a la competencia con microorganismos presentes en el compost, es especialmente importante en el caso de patógenos con propágulos pequeños como Pythium y Phytophthora que, al tener pocas reservas, requieren de fuentes externas de C y otros nutrientes y por lo tanto se ven afectados por la competencia con las altas poblaciones de microorganismos presentes en el compost (Hoitink et al. 1997, Diánez et al. 2007). En forma concordante, van Os et al. (2001), en estudios realizados con Pythium macrosporum, encontraron que una alta biomasa y actividad microbiana indujeron la supresión de crecimiento de Pythium. En este mismo sentido Craft y Nelson (1996) indican que altas poblaciones de microorganismos, hongos y actinomicetos presentes en el compost, fueron los agentes supresivos en la enfermedad “dampingoff” causada por Pythium graminicola. Adiobo (2006) encontró que la supresividad de 3 tipos de compost sobre P. myriotylum se perdió al autoclavar los abonos y que fue parcialmente recuperada al restaurar la microflora original ya que inoculó los abonos con pequeñas cantidades del abono no tratado. Además, el compost que presentó una mayor supresividad fue el que contuvo una mayor cantidad de microorganismos.
Adicionalmente, en el rumen de herbívoros, en el estiércol y en abonos orgánicos a base de estiércol, se encuentran altas poblaciones de microorganismos celulolíticos (Varel y Dehority 1989, Galindo et al. 2008); estos microorganismos producen celulasas, enzimas que son especialmente importantes en la degradación de la pared celular de oomycetes tales como Pythium, constituida principalmente por celulosa y beta glucanos (Inglis y Kawchuk 2002, El-Tarabily 2006, Rossman y Palm 2006). Los abonos de origen animal han sido utilizados como fuente de microorganismos celulolíticos para la degradación de broza, residuos animales (Kim et al. 2004, Terry et al. 2004) y como fuente de microorganismos supresores de Pythium (Inglis y Kawchuk 2002).
Al respecto, Ringer et al. (1997) encontraron que abonos obtenidos de diferentes tipos de desechos animales presentaron supresividad hacia Pythium y Rhizoctonia. Específicamente, compost obtenidos de estiércol de vaca causaron una mayor supresión sobre Pythium. Mora et al. 1987 en el diseño de un manejo integrado de la tristeza del aguacate, causada por Phytophthora cinamoni, encontraron que los tratamientos en los que se aplicó estiércoles presentaron la menor cantidad de aislamientos de Phytophthora, mayor peso seco de raíces, mejor apariencia de follaje y mayor producción de frutos; estos tratamientos exhibieron los niveles mas bajos de incidencia de la enfermedad. Los autores sugieren que el estiércol induce supresividad del hongo, aumenta el número de hongos y bacterias antagónicas y el contenido de nutrientes de los suelos. Asi mismo, Rodríguez-Guzmán et al. 1992 encontraron para este mismo cultivo que la aplicación de estiércol disminuyó la población de Phytophthora y Pythium, la incidencia de la enfermedad y aumentó el desarrollo radical del hospedero presentándose, con la adición del estiércol, condiciones similares a los obtenidos en suelos supresivos.
Un factor a considerar en el uso de abonos para el control de enfermedades es que abonos maduros muy estabilizados podrían tener poblaciones de microorganismos poco activas y con poco efecto supresor como en el caso de CBM y el VBM, o una capacidad limitada para sostener la actividad de la biomasa microbiana en el suelo (Hoitink et al. 1993, Suárez-Estrella et al. 2007). Zmora-Nahum et al. (2008), analizó el efecto del proceso de maduración sobre 3 tipos de compost supresivos contra Slerotium rolfsii y Pythium aphanidermatum y encontraron que la pérdida de supresión se asoció entre otros, a una disminución en la actividad microbiana. De acuerdo con Hoitink et al. (1997), el compost demasiado estabilizado no mantiene poblaciones de biocontroladores eficaces y no es por lo tanto adecuado para la supresión de enfermedades.
Los abonos orgánicos varían en su efecto para suprimir enfermedades de la planta. En la presente investigación, los abonos obtenidos a partir de estiércol fueron más supresivos, posiblemente por una mayor actividad microbiana, que los producidos a base de broza de café. Se concluye que el vermicompost a base de estiércol de vaca tiene un efecto supresivo sobre el sistema Tiquisque -P. myriotylum, por lo que debe investigarse el origen de esta supresividad. En investigaciones futuras se debe evaluar el uso de proporciones mayores de abono, las cuales podrían mejorar el nivel de supresión obtenido.
Agradecimientos
Este trabajo se financió con fondos de los proyectos 230-04-FI. Uso de abonos orgánicos para el control de enfermedades de importancia agrícola, Fondo de Incentivos MICIT-CONICIT y VI 733-A5-008, Vicerrectoría de Investigación, Universidad de Costa Rica.
Este trabajo se financió con fondos de los proyectos 230-04-FI. Uso de abonos orgánicos para el control de enfermedades de importancia agrícola, Fondo de Incentivos MICIT-CONICIT y VI 733-A5-008, Vicerrectoría de Investigación, Universidad de Costa Rica.
[ Links ]
Adiobo A. 2006. Biological control of cocoyam (Xanthosoma sagittifolium (L.) Schott) root rot disease caused by Pythium myriotylum Dreschl.: importance of soil organic matter content and cultural practices. Tesis de doctorado, Universidad de Gante, Flandes, Bélgica. 181 p. [ Links ]
Anderson J. 1982. Soil Respiration, pp. 831-895. In: A. Page, R. Miller y D. Keeney (eds). Methods of Soil Analysis. Part 2: Chemical and Microbiological Properties. Soil Science Soc. Am. Inc. [ Links ]
Arauz L.F. 1998. Fitopatología, un enfoque agroecológico. Editorial de la Universidad de Costa Rica, Costa Rica. 467 p. [ Links ]
Atlas R.M., Bartha R. 2002. Ecología microbiana y microbiología ambiental. 4º edición en español. Pearson Educación S.A, Madrid. 677 p. [ Links ]
Bollen G.J. 1993. Factors involved in inactivation of plant pathogens during composting of crop residues, pp. 301-318. In: H.A.J. Hoitink y H.M. Keener (eds). Science and Engineering of Composting: Design, Environmental, Microbiological and Utilization Aspects. Renaissance Publications. [ Links ]
Bonanomi G., Antignani V., Pane C., Scala F. 2007. Suppression of soilborne fungal diseases with organic amendments. Journal of Plant Pathology 89:311-324. [ Links ]
Bremner J.M., Tabatabai M.A. 1971. Use of automated combustion techniques for total carbon, total nitrogen and total sulphur analysis of soil, pp. 1-16. In: L.M. Walsh (ed). Instrumental Methods for Analysis of Soils and Plant Tissue. Soil Science Society of America. [ Links ]
Chefetz B., Hatcher P.G., Hadar Y., Chen Y. 1996. Chemical and biological characterization of organic matter during composting of municipal solid waste. Journal of Environment Quality 25:776-785. [ Links ]
Chen Y., Inbar Y. 1993. Chemical and spectroscopical analyses of organic matter transformations during composting in relation to compost maturity, pp. 551-600. In: H.A.J. Hoitink y H.M. Keener (eds). Science and Engineering of Composting: Design, Environmental, Microbiological and Utilization Aspects. Renaissance Pubblications. [ Links ]
Composting Council Research and Education Foundation (CCREF) 2001.Test Methods for the Examination of Composting and Compost (TMECC). Disco compacto. Houston, USA. United States Composting Council (USCC), USDA. 2001, August 21. [ Links ]
Craft C., Nelson E. 1996. Microbial properties of composts that suppress damping-off and root rot of creeping bentgrass caused by Pythium graminicola. Applied and Environmental Microbiology 62:1550- 1557. [ Links ]
Danon M., Zmora-nahum S., Chen Y., Hadar Y. 2007. Prolonged compost curing reduces suppression of Sclerotium rolfsii. Soil Biology and Biochemistry 39:1936–1946. [ Links ]
Diánez F., Santos M., Tello J.C. 2007. Suppressive effects of grape marc compost on phytopathogenic oomycetes. Archives of Phytopathology and Plant Protection 40:1-18. [ Links ]
Domínguez J., Edwards C.A., Subler S. 1997. A comparison of composting and vermicomposting. BioCycle: 57-59. [ Links ]
El-Tarabily K.A. 2006. Rhizosphere-competent isolates of streptomycete and non-streptomycete actinomycetes capable of producing cellwall-degrading enzymes to control Pythium aphanidermatum damping-off disease of cucumber. Canadian Journal of Botany 84:211-222. [ Links ]
Erhart E., Burian K. 1997. Evaluating quality and suppressiveness of Austrian biowaste composts. Compost Science and Utilization 5:15-25. [ Links ]
Galindo J., González N., Delgado D., Sosa A., Marrero Y., González R., Aldana A.I., Moreira O. 2008. Efecto modulador de Leucaena leucocephala sobre la microbiota ruminal. Zootecnia Tropical 26(3):249-252. [ Links ]
Garcia C., Pascal J.A., Mena E., Hernández T. 2004. Influence of the stabilisation of organic materials on their biopesticide effect in soils. Bioresource Technology 95:215-221. [ Links ]
Gómez L. 1993. El “mal seco” del tiquisque. Taller Aplicaciones de la biotecnología en raíces, tubérculos y pejibaye. Cultivo de Tejidos. Centro de Investigaciones Agronómicas. Universidad de Costa Rica. s.p. [ Links ]
Gómez-Brandón M., Lazcano C., Domíngez J. 2008. The evaluation of stability and maturity during the composting of cattle manure. Chemosphere 70:436-444. [ Links ]
Hoitink H.A.J., Boehm M.J. 2001. Control biológico en comunidades microbianas del suelo: un fenómeno de dependencia de sustrato FORO. Manejo Integrado de Plagas (Costa Rica) N°. 62 p 4-17. [ Links ]
Hoitink H.A.J., Boehm M.J., Hadar Y. 1993. Mechanisms of suppression of soilborne plant pathogens in composts-amended substrates, pp. 601-621. In: H.A.J. Hoitink y H.M. Keener (eds). Science and Engineering of Composting: Design, Environmental, Microbiological and Utilization Aspects. Renaissance Pubblications. [ Links ]
Hoitink H.A.J., Grebus M. 1994. Status of biological control of plant diseases with composts. Compost Science and Utilization 2:6-12. [ Links ]
Hoitink H.A.J., Stone A.G., Han D.Y. 1997. Supresión de enfermedades mediante compost. Agronomía Costarricense 21(1):25-33. [ Links ]
Huber D.M. 1980. The role of mineral nutrition in disease, pp. 381-406. In: J.G. Horsfall y E.B. Cowling (eds). Plant Disease, an Advanced Treatise Vol 3. Academic Press. [ Links ]
Huber D.M. 1991. The use of fertilizers and organic amendments in the control of plant disease, pp. 405-494. In: D. Pimentel (ed). Handbook of Pest Management in Agriculture. CRC Press. [ Links ]
Inglis G.D., Kawchuk L.M. 2002. Comparative degradation of oomycete, ascomycete, and basidiomycete cell walls by mycoparasitic and biocontrol fungi. Canadian Journal of Microbiology 48:60-70. [ Links ]
Kim T.I., Jeong K.H., Ham J.S., Yang C.B., Chung I.B., Kim M.K., Kim K.N. 2004. Isolation and characterization of cellulase secreting bacterium from cattle manure: Application to Composting. Compost Science & Utilization 12:242-248. [ Links ]
Laguna I., Salazar L., López J. 1983. Enfermedades fungosas y bacterianas de aráceas: Xanthosoma spp. y Colocasia esculenta (L.) Schott en Costa Rica. Turrialba, Costa Rica. CATIE. 32 p. [ Links ]
Lazcano C., Gómez-Brandón M., Domíngez J. 2008. Comparison of the effectiveness of composting and vermicomposting for the biological stabilization of cattle manure. Chemosphere 72:1013-1019. [ Links ]
Lievens B., VAES K., Coosemans J., Ryckeboer J. 2001. Systemic resistance induced in cucumber against Pythium root rot by source separated household waste and yard trimmings composts. Compost Science and Utilization 9:221-229. [ Links ]
Malandraki I., Tjamos S.E., Pantelides I.S., Paplomatas E.J. 2008. Thermal inactivation of compost suppressiveness implicates possible biological factors in disease management. Biological Control 44:180-187. [ Links ]
Mora G., Teliz D., Garcia R. 1987. Manejo integrado de la tristeza (Phytophthora cinnamomi) del aguacate (Persea americana). Cuarta evaluación anual. Revista Mexicana de Fitopatología 6:76-81. [ Links ]
Noble R., Coventry E. 2005. Suppression of soil-borne plant diseases with composts: A review. Biocontrol Science and Technology 15:3-20. [ Links ]
Ouatmane A., Provenzano M.R., Hafidi M., Senesi N. 2000. Compost maturity assessment using calorimetry, spectroscopy and chemical analysis. Compost Science and Utilization 8:124- 134. [ Links ]
Pacumbaba R.P., Wutoh J.G., Eyango S.A., Tambong J.T., Nyochembeng L.M. 1992. Isolation and pathogenicity of rhizosphere fungi of cocoyam in relation to cocoyam root rot disease Journal of Phytopathology 135:265-273. [ Links ]
Paul E.A., Clark F.E. 1996. Soil Microbiology and Biochemistry. 2° ed. Academic Press, California. 340 p. [ Links ]
Perneel M., Tambong J.T., Adiobo A., Floren C., Saborio F., Lévesque A., Höfte M. 2006. Intraspecific variability of Pythium myriotylum isolated from cocoyam and other host crops. Mycological Research 110:583-593. [ Links ]
Ringer C., Millner P., Teerlink L., Lyman B. 1997. Suppression of Seedling Damping-Off Disease in Potting Mix Containing Animal Manure Composts. Compost Science and Utilization 5:6-14. [ Links ]
Rodríguez-Guzman M.P., Garcia R., Teliz D., Salazar G. 1992. Ecología de la enfermedad “tristeza del aguacatero” bajo manejo experimental del cultivo del aguacate en la región de Atlixco, Puebla, México. Agrociencia serie Protección vegetal 3:115-135. [ Links ]
Rossman A.Y., Palm M.E. 2006. Why are Phytophthora and other Oomycota not true Fungi? APSnet, The American Phytopathological Society, 3340 Pilot Knob Road, St. Paul, MN 55121-2097, USA. (Downlowded 29/01/2010. Available online: http://www.apsnet.org/online/feature/oomycetes/). [ Links ]
Saborío F. 2007. Cocoyam and Breeding. In: S. Ochatt y S.M. Jain (eds). Breeding of neglected and underutilized crops, species and herbs., Science Publishers, New Hampshire, USA. 447 p. [ Links ]
Scheuerell S.J., Mahaffee W.F. 2005. Microbial recolonization of compost after peak heating needed for the rapid development of damping-off suppression. Compost Science and Utilization 13:65-71. [ Links ]
Schüler C., Biala J., Bruns C., Gottschall R., Ahlers S., Vogtmann H. 1989. Suppression of root rot on peas, beans and beetroots caused by Pythium ultimum and Rhizoctonia solani through the amendment of growing media with composted organic household waste. Journal of Phytopathology 127:227-238. [ Links ]
Singleton L., Mihail J., Rush C. 1992. Methods for research on soilborne phytopathogenic fungi. APS Press, Minnesota. 265 p. [ Links ]
Somasegaran P., Hoben H. 1985. Methods in legume-Rhizobium technology. University of Hawai. Niftal Project and Mircen. Hawai. 367 p. [ Links ]
Suárez-Estrella F., Vargas-García C., López M.J., Capel C., Moreno J. 2007. Antagonistic activity of bacteria and fungi from horticultural compost against Fusarium oxysporum f.sp.melonis Crop Protection 26:46-53. [ Links ]
Tambong J.T. 2001. The cocoyam root rot disease pathogen, Pythium myriotylum: Pathogenicity, isozyme characterization, nature of resistance and biological control. Thesis submitted in fulfillment of the requirements of the degree of doctor (Ph.D.) in Applied Biological Sciences-Agronomy. Faculteit Landbouwk Undige en Toegepaste Biologische Wetenschappen. Universiteit Gent. p 1-17. [ Links ]
Termorshuizen A.J., Van Rijn E., Van Der Gaag D.J., Alabouvette C., Chen Y., Lagerlöf J., Malandrakis A.A., Paplomatas E.J., Rämert B., Ryckeboer J., Steinberg C., Zmora-Nahum S. 2006. Suppressiveness of 18 composts against 7 pathosystems: Variability in pathogen response. Soil Biology and Biochemistry 38:2461-2477. [ Links ]
Terry A.I., Bermúdez R.C., Rodríguez S., Fernández M. 2004. Selección de un inóculo para labiodegradación anaerobia de la pulpa de café. Tecnología Química 24:64-71. [ Links ]
Van Os G.J., Van Ginkel J.H. 2001. Suppression of Pythium root rot in bulbous Iris in relation to biomass and activity of the soil microflora. Soil Biology and Biochemistry 33:1447-1454. [ Links ]
Varel V.H., Dehority B.A. 1989. Ruminal cellulolytic bacteria and protozoa from bison, cattle-bison hybrids, and cattle fed three alfalfa-corn diets. Applied and Environmental Microbiology 55:148-153. [ Links ]
Veeken A.H.M., Blok W.J., Curci F., Coenen G.C.M., Termorshuizen A.J., Hamelers H.V.M. 2005. Improving quality of composted biowaste to enhance disease sppressiveness of compost-amended, peat-based potting mixes. Soil Biology and Biochemistry: 2131-2140. [ Links ]
Walkley A., Black C.A. 1938. An examination of the Degtajareff’s method for determination soil organic matter and a proposed modification of the chromic acid titration method. Soil Science 37:29-38. [ Links ]
Wu L., Ma L.Q., Martinez G.A. 2000. Comparison of methods for evaluating stability and madurity of biosolids compost. Journal of Environmental Quality 29:424-429. [ Links ]
Zmora-Nahum S., Danon M., Hadar Y., Chen Y. 2008. Compost curing reduces suppression of plant diseases. Compost Science and Utilization 16:250-256. [ Links ]
Zmora-Nahum S., Hadar Y., Chen Y. 2007. Physico-chemical properties of commercial composts varying in their source materials and country of origin. Soil Biology and Biochemistry 39:1263-1276. [ Links ]
Correspondencia a: Silvia Artavia. Instituto Tecnológico de Costa Rica. Cartago, Costa Rica.
Lidieth Uribe. Centro de Investigaciones Agronómicas, Universidad de Costa Rica. San José, Costa Rica / Correo electrónico: lidieth.uribe@ucr.ac.cr
Francisco Saborío & Leida Castro. Centro de Investigaciones Agronómicas, Universidad de Costa Rica. San José, Costa Rica.
Luis Felipe Arauz. Centro de Investigación en Protección de Cultivos, Universidad de Costa Rica. San José, Costa Rica.
Recibido: 18/02/10 Aceptado: 12/05/10

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}