











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkAntecedentes
Laurito (1999) describió 2 dientes laterales de tiburón de muy pequeña talla, < 2 mm, correspondientes a los ejemplares CLM-163 (CFM-4009) y CLM-164 (CFM-4011), hoy resguardados en la Sección de Geología del Museo Nacional de Costa Rica; los cuales en su momento dicho autor asoció al género Scylliorhinus e incluso erigió una especie nueva cuya validez se analiza más adelante; en el caso del segundo espécimen, dicho autor destacó su proximidad morfológica a la especie Scylliorhinus joneti Cappetta, 1970 del Mioceno inferior de Francia.
En el presente trabajo realizamos una revisión taxonómica y una nueva descripción de los ejemplares supra citados, para ello realizamos nuevas comparaciones con el fin de establecer una determinación taxonómica más apropiada, conforme a los avances en la morfología dental comparada de condrictios de las últimas dos décadas.
Los ejemplares proceden de la localidad fosilífera de Alto Guayacán, 8 km al suroeste de la ciudad de Siquirres. Estratigráficamente fueron recolectados en la Formación Uscari, en una de sus secciones más jóvenes con una edad Mioceno Superior cuspidal - Plioceno Inferior basal equivalentes a las biozonas N17, N18 y N19 de foraminíferos planctónicos, correspondientes con las épocas Messiniano-Zancleano (Figura 1).
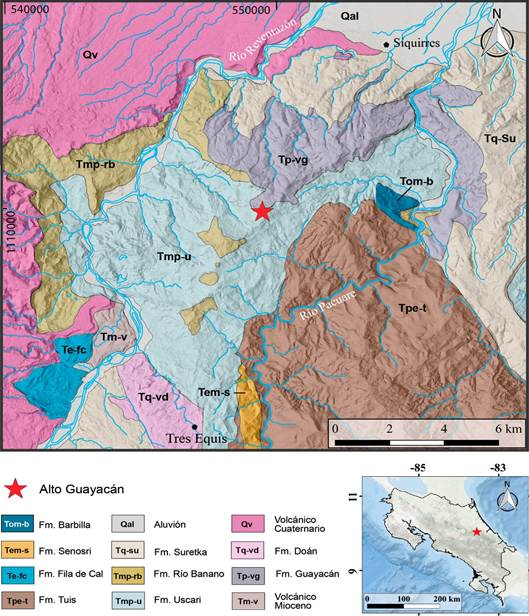
Figura 1 Mapa geológico de una parte de la vertiente Caribe (entre las cuencas de los ríos Reventazón y Pacuare) y de ubicación de la localidad fosilífera de Alto Guayacán en el contexto local y en Costa Rica, cuyos sedimentos marinos corresponden con la Formación Uscari (Tmp-u) en la sección noroccidental de la cuenca Limón Sur, las redes de drenaje están representadas por líneas celestes y los poblados más importantes están representados por pequeños pentágonos negros. Datos geológicos compilados a partir de observaciones propias y los trabajos de: Bottazzi, Obando y Fernández (2021), Cervantes (1989), Denyer y Alvarado (2007), Fernández (1987), Laurito (1999), Segura, Badilla y Obando (2011) y Tournon (1984).
Paleontología
Orden Orectolobiformes Applegate, 1972
Familia Parascyliidae Gill, 1862
Género Parascyllium Gill, 1862
Parascyllium sp.
Sinonimia:
-Scylliorhinus sp. en Laurito (1999), pág. 54, lám. 8, figs. 1(a-c).
-Scylliorhinus ambiatlanticus en Laurito (1999), pág. 55, lám. 8, figs. 2(a-e).
Materiales: El CFM-4009 (CLM-163) diente anterior lateral inferior, figura 2(1a-1c) y el CFM-4011 (CLM-164) diente anterior de ubicación incierta, figura 2(2a-2e).

Figura 2 Parascyllium sp. de la localidad de Alto Guayacán, Fm. Uscari. Ejemplar CFM-4009 en vistas 1a. vestibular, 1b. linguo basal y 1c. mesial y ejemplar CFM-4011 en vistas 2a. superior, 2b. labial, 2c. basal, 2d. mesial y 2e. lingual.
Características dentales: Coronas infladas y altas, con cúspides altas, volcadas en sentido lingual y ápice coronal aparentemente redondeados, en ambos ejemplares se observa evidencia de desgaste de los ápices. Las caras lingual y vestibular de las coronas, son tersas y convexas, limitadas por un filo suave no aserrado que se extiende de manera continua a las cúspides accesorias. Estas últimas son notablemente pequeñas en comparación con la cúspide principal, infladas y volcadas hacia atrás. En el caso del ejemplar CFM-4009, las cúspides laterales (fig. 2.1c) se asientan sobre cortas proyecciones de los lóbulos radiculares, mientras que en el caso del ejemplar CFM-4011 las cúspides laterales o accesorias se proyectan a manera de alas, ampliamente extendidas hacia los lados y hacia la región lingual, culminando con proyecciones redondeadas ligeramente infladas, sobre sendos lóbulos radiculares.
En el ejemplar CFM-4009, la protuberancia lingual, aunque relativamente ancha y amplia en sentido vertical, es poco prominente, carece de ornamentación y no llega a proyectarse por delante de la cara vestibular de la raíz.
En el ejemplar CFM-4011, la protuberancia lingual basal de la corona es ancha, alta, inflada y carente de ornamentación (figs. 2.2a y 2.2b) y se proyecta ligeramente por sobre la cara vestibular de la raíz, pero no llega a cubrirla o envolverla.
En ambos casos se observa un cuello lingual angosto en el ejemplar CFM-4009 (figs. 2.1b y 2.1c) y un cuello lingual más ancho en el ejemplar CFM-4011 (figs. 2.2d y 2.2e).
Las raíces son abultadas con conspicuas y redondeadas proyecciones labiales, y amplias y alargadas proyecciones linguales, sus caras basales son planas y de contorno general “triangular”. Los lóbulos están separados por un recto, alargado y notable surco mediano que se proyecta verticalmente en la cara lingual (figs. 2.1b, 2.2c y 2.2e) y contiene los forámenes nutricios.
Distribución paleogeográfica y actual: El registro de Parascyllium sp. para el Mesiniano - Zancleano de Alto Guayacán, constituye el único registro fósil conocido del género para el Caribe y la paleoprovincia neógena Gatuniense-Caribeña, así como de todo el Cenozoico neotropical americano. Tampoco se registra en el Neógeno del Continente Americano de allí la relevancia de su registro.
Durante el Paleógeno la distribución del género Parascyllium fue mucho más amplia, registrándose por ejemplo en el Paleoceno temprano de la provincia de Chubut, Argentina en la Formación Salamanca (Bernárdez Rodríguez, 1997) y el Paleoceno temprano de Suecia (Siverson, 1993); desafortunadamente en ninguna de estas publicaciones se ilustran los ejemplares. Por su parte, Adnet (2006) registra Parascyllium sp. en el Lutetiano, Eoceno medio, del suroeste de Francia (Cappetta, 2012, fig. 151, pág.164) (ver figura 3).
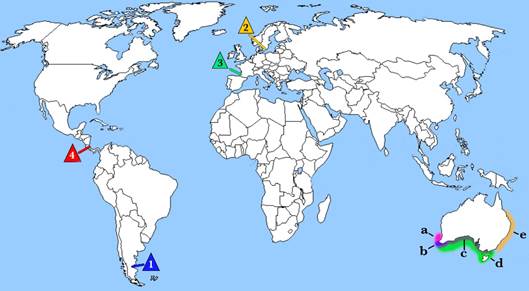
Figura 3 Distribución de los registros fósiles del género Parascyllium: Daniano medio - superior de: 1. Formación Salamanca, Chubut, Argentina y 2. Limhamn quarry, Suecia; 3. Luteciano medio de Angoumé, Landes en el suroeste de Francia y 4. Zancleano-Messiniano, Formación Uscari, Costa Rica. Distribución de las especies actuales de Parascyllium: a. P. sparsimaculatum b. P. elongatum, c. P. variolatum, d. P. ferrugineum y e. P. collare.
En la actualidad todas las especies conocidas de Parascyllium habitan las plataformas continentales e insulares de Australia meridional, no habitando en otras regiones del globo (ver fig. 3 a-e).
Discusión
La morfología general de la corona recuerda a la de algunos géneros de la familia Scylliorhinidae, en particular a Poroderma y Scylliorhinus, sobre manera la del ejemplar CFM-4009. Sin embargo, en dichos géneros la corona no se inclina o vuelca lingualmente de manera tan radical formando un ángulo de casi 90 grados (fig. 2.1c), lo que sí se observa en el género Parascyllium.
Al analizar la raíz, entre las diferencias morfológicas más notables que se observan a nivel basal, se nota que los lóbulos radiculares en Scylliorhinidae tales como Scylliorhinus, Cephaloscyllium y Aulohalaelurus son palmeados y/o se proyectan de manera conspicua en sentido vestibular y que el canal mediano se ensancha notablemente por detrás de los lóbulos radiculares (ver láminas 8, 10 y 12 de Herman, Hovestadt-Euler y Hovestadt, 1990). Mientras que en Parascyllium, los lóbulos radiculares si bien se proyectan ligeramente, son mucho más angostos y el canal mediano se mantiene con un ancho uniforme a lo largo del eje vestíbulo lingual, cuasi rectilíneo, abriéndose ligeramente hacia la región vestibular (figs. 2.1b, 2.2c), ver las láminas 23 y 25 de Herman, Hovestadt-Euler y Hovestadt (1992), y además se eleva a lo largo de la protuberancia lingual (figs. 2.1b, 2.2e), lo que no sucede en Scylliorhinidae.
De las especies actuales del género Parascyllium concordamos con la observación hecha por el Dr. J. Kriwet (com. escrita 4/12/2022) de que los ejemplares recuerdan a los de la especie Parascyllium collare, pero con la diferencia de que en los ejemplares de Alto Guayacán las cúspides laterales son mucho más pequeñas. Además, los dientes de Parascyllium collare, mayoritariamente presentan una ornamentación a manera de rugosidades verticales en la cara vestibular, algo que no se observa en los ejemplares del presente estudio.
Conclusiones
Las características dentales de los especímenes analizados en este estudio, sugieren que su morfología general es muy similar a la que presentan algunas especies de Parascyllium, aunque también recuerdan a la morfología observable en algunos Scylliorhinidae. Sin embargo, las características radiculares, discutidas arriba, son concluyentes en cuanto a la pertenencia de los ejemplares CFM-4009 y CFM-4011 al género Parascyllium y no a Scylliorhinus.
En cuanto a la especie Scylliorhinus ambiatlanticusLaurito 1999, consideramos prudente declararla nomen nudum, no solo por su pertenencia a otro género, sino, además, porque al parecer ambos especímenes podrían ser co-específicos y por lo tanto la descripción original de dicha especie no abarcó las características dentales de los 2 ejemplares.
Por último, el género Parascyllium que en la actualidad está confinado a las aguas meridionales de Australia, registra 5 especies (fig. 3), todas habitantes de la plataforma continental con rangos de profundidad variable entre los 5 y los 435 m de profundidad (Compagno, Dando y Fowler, 2005; Kyne, Heupel, White y Simpfendorfer, 2021). Su biología es prácticamente desconocida y son consideradas especies de hábito bentónico. Lo anterior permite caracterizar la paleoecología de las especies fósiles, como tiburones bentónicos habitantes de las plataformas y de los taludes superiores tanto insulares como continentales durante el Cenozoico y Cuaternario.

