











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Los Hippariones son un conjunto polifilético de caballos tridáctilos que vivieron durante el Mioceno y Plioceno del Nuevo y Viejo Mundo. Cuatro géneros han sido reconocidos para América del Norte y América Central, estos son: Hipparion sensu stricto, Neohipparion, Nannipus y Cormohipparion (cf. MacFadden, 1984).
En América Central se registran las especies Cormohipparion ingenuum (Leidy, 1885) y Cormohipparion plicatile (Leidy, 1887) en el Hemphilliano Temprano de la Localidad de Rancho Lobo en Honduras (Webb y Perrigo, 1984; Hulbert, 1988a y 1988b). Por su parte, en la localidad de Lago Alajuela y en la formación homónima en la región central de Panamá, MacFadden et al., 2017 registran Cormohipparion sp. con una edad Clarendoniano tardío o Hemphilliano temprano (Cl3 - Hh1).
El molar CFM-5371, objeto de estudio del presente informe, fue recuperado en el lecho del Río Pataste, en las proximidades del pueblo Pataste Arriba, Cantón de Guatuso, Distrito San Rafael, Provincia de Alajuela, en las coordenadas 10º33’52’’N y 84º40’08’’ W, hoja cartográfica Monterrey 3247 I IGN, ver figura 1.
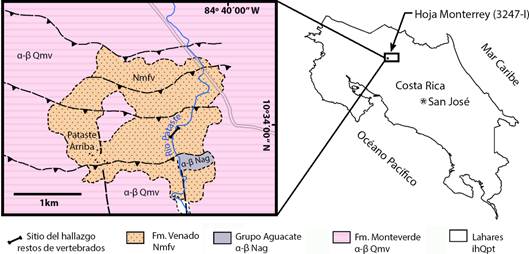
Fig. 1 mapa geológico de la localidad próxima a Pataste, redibujado y fundamentado en Rojas (2019) con la ubicación del sitio del hallazgo.
Geología
De acuerdo con Huapaya et al., 2017 y Rojas (2019) se puede establecer que el fósil procede de la Formación Venado (Nmfv) de edad Mioceno Medio - Superior, caracterizada por una litología variada compuesta de areniscas, limolitas, lutitas, calizas bioclásticas, brechas, conglomerados y capas de carbón (Obando, 1986), ver figura 1. En esta área la Formación Venado está sobreyacida por basaltos, andesitas basálticas y traquiandesitas basálticas del Grupo Aguacate del Mioceno. Además de brechas, tobas y lahares subordinados pertenecientes a la Formación Monteverde del Pleistoceno Inferior y lahares (ihQpt) del Reciente, sensu Rojas (2019)
Paleontología sistemática
Familia EQUIDAE Gray, 1821
Subfamilia EQUINAE Gray, 1821
Tribu HIPPARIONINI Quinn, 1955
Género CORMOHIPPARION Skinner & MacFadden, 1977
Cormohipparion quinniWoodburne, 1996
Sinonimia:
-Cormohipparion sphenodus (Cope), 1889 en MacFadden, 1984 Fig. 127, pág. 159 y págs. 156 a 162 en parte.
Material: un molar superior M1 o M2 aislado, depositado en la colección de fósiles del Museo Nacional de Costa Rica, bajo el acrónimo CFM-5371 (figs. 2 y 3).
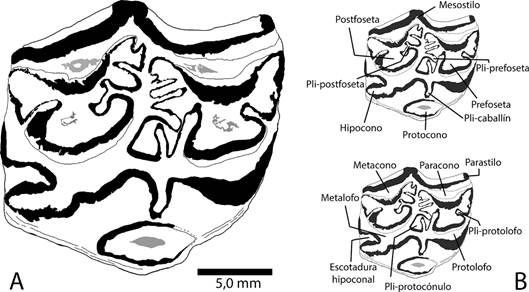
Fig. 2 Cormohipparion quinni Woodburne 1996 A. detalle de la superficie oclusal del ejemplar CFM-5371, B. nomenclatura dental utilizada en este trabajo, basada en Stirton, 1941; Skinner y MacFadden, 1977 y MacFadden, 1984.
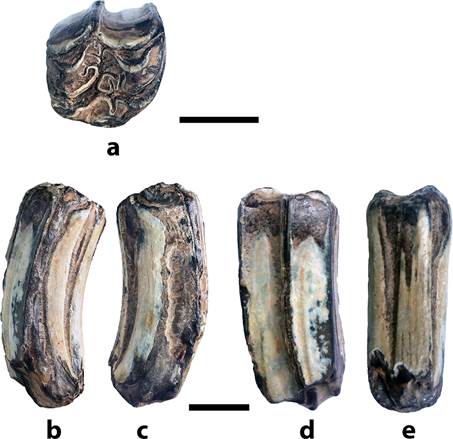
Fig. 3 CFM-5371, molar superior derecho M1 o M2 de Cormohipparion quinni Woodburne 1996 en vistas: a. oclusal, b. comisural, c. mesial, d. vestibular y e. lingual. Escalas visuales equivalen a 1 cm.
Descripción: El ejemplar CFM-5371 corresponde a un M1 o M2 derecho, la superficie oclusal es de contorno cuasi cuadrado. La prefoseta y postfoseta tienen forma de media luna con varios pliegues en el borde de esmalte. Las plicaciones en la prefoseta son numerosas, se observa un único pli-protolofo, tres plis de la prefoseta, el pli-protocónulo es doble y desarrollado con la pared de esmalte muy recta. Las plicaciones de la postfoseta: un pli hipóstilo sencillo, dos plis postfoseta sencillos y uno doble.
El parastilo y el metastilo están ausentes, mientras que el mesostilo es angular y pronunciado.
El hipocono es elongado, el surco hipoconal es abierto. El paracono, metacono, metalofo y protolofo son angostos. El pli-caballín es simple y amplio. El protocono es ovalado y está aislado del protolofo.
Distribución paleogeográfica y edad (14.8 -12.5 Ma)
Cormohipparion quinniWoodburne 1996, ha sido registrado en sedimentos de los Sand Canyon Beds en el Estado de Colorado, en el Estado de Nebraska en las localidades de Devil’s Gulch Horse Quarry, Sawyer Quarry y Valentine Railway Quarries en sedimentos de Devil’s Gulch Member, y las localidades de Railway Quarry, Schoettger Quarry, Nenzel Quarry y Devil’s Jump Off Q. correspondientes a sedimentos de Crooston Bridge Member, ambos miembros de Valentine Formation (sensu, Woodburne, 1996 y 2007). En el Estado de South Dakota, en las cercanías de South Bijou en Charles Mix County, en sedimentos del Bone Hash Layer Hill, Fort Randall Formation (Pagnac, 2011). Todas las localidades corresponden al Mioceno Medio con edades mamífero norteamericana Barstoviano Tardío (Ba 2).
En México, Bravo-Cuevas y Ferrusquía-Villafranca (2008) registran la especie Cormohipparion aff. C. quinniWoodburne 1996 en la localidad de Nejapa en el Estado de Oaxaca, en sedimentos fluvio lacustrinos de la Formación el Camarón del Mioceno Medio y edad mamífero norteamericana Barstoviano Tardío temprano (Ba 2), ver figura 4.
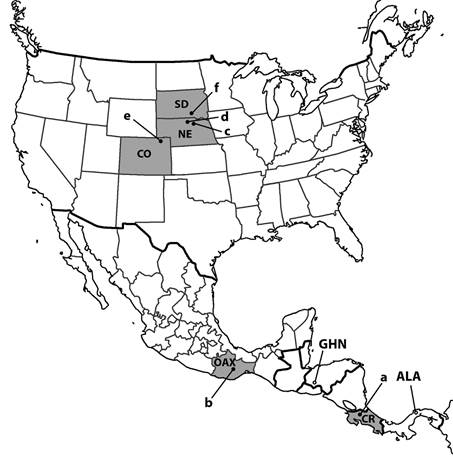
Fig. 4 Localidades Barstoviano tardías donde se ha registrado Cormohipparion quinni Woodburne 1996 en América del Norte y Central; a. Río Pataste, Costa Rica; b. Nejapa en el Estado de Oaxaca, México; c. Localidades del Crooston Bridge Member y d. Localidades del Devil’s Gulch Member en el Estado de Nebraska; e. Sand Canyon Beds en el Estado de Colorado y f. Bone Hash Layer Hill en el Estado de South Dakota. Además, se indican otras localidades de América Central donde se registra el género Cormohipparion como: GHN. Formación Gracias en Honduras con las especies Cormohipparion ingenuum (Leidy, 1885) y Cormohipparion plicatile (Leidy, 1887) de edad Hemphilliano Temprano, y ALA. Formación Alajuela en los alrededores del lago del mismo nombre en la región central de Panamá, donde se registró Cormohipparion sp.
Discusión
El diente hallado corresponde a un molar superior el M1 o M2 de mediano tamaño mesodonto a hypsodonto, con patrón oclusal complejo y característico protocono aislado lo que nos confirma que se trata de hipparionino tridáctilo. El protocono tiene un contorno elíptico con una razón de proporción de 0,53, ligeramente mayor que la registrada para C. quinniWoodburne 1996 que es de 0,50 a 0,51, pero esa ligera inconsistencia puede deberse a que el protocono está erosionado; tal razón protoconal es coincidente con la especie C. matthewi, pero el M1 o M2 de ésta especie tiene un índice de plicaciones sumamente complejo 2:8:6:1 en el caso del ejemplar CFM-5371 posee un índice de pliegues o plicaciones de la prefoseta y postfoseta de 1:5:4:1, un índice que ha sido registrado unicamente para los M1 de C. quinniWoodburne 1996 (ver tabla 21 de Woodburne, 2007), lo que descarta a las otras especies del género Cormohipparion y de otros géneros de Hiparionini. Además, el protocono carece del “spur” o gancho en su extremo anterolabial que es característico de la especie C. goorisi, (ver figs. 2 y 3, Cuadro 1).
Cuadro 1 Medidas en milímetros del molar superior de Cormohipparion quinni.
| M1 o M2 | Protocono | ||||||||
| CFM-5371 | L | PRR | A | AMST | %Des | LO | AO | PLI | índice |
| 5.74 | 0.53 | 3.09 | 40,91 | 9,01 | 18.04 | 17.22 | 1 | 1:5:4:1 | |
Protocono: L= largo, A= ancho, PRR= razón protoconal; AMST= altura del mesostilo; LO= largo oclusal; AO= ancho oclusal; índice= # total de pliegues presentes en la prefoseta y postfoseta; Pli= # de pliegues en pli-caballín, %Des= porcentaje de desgaste del molar.
Por otra parte el pli-protocónulo o lazo de la prefoseta es bastante complejo de contorno trapozoidal elongado y un lazo triangular adherido, en general es más simple que el observado en C. occidentale y notablemente más complejo que en las restantes especies del género.
El pli-caballín en el ejemplar CFM-5371 es simple, está constituido por un único pliegue notoriamente largo, curvo y ligeramente grueso lo que permite diferenciarlo de las especie C. matthewi, C. fricki, C. occidentale y C. goorisi donde los M1 o M2 presentan un pli-caballín que suele ser corto, tener 2 o más pliegues o ser más complejo.
El molar recuperado presenta un relieve oclusal alto y las cúspides dentales: el metacono y el paracono, son redondeadas, el índice de hipsodoncia es 2,27. El patrón de desgaste es muy similar al observado en otros molares de C. quinniWoodburne 1996, lo que sugiere una dieta mixta.
Paleoambiente
Obando (1984) sugiere un escenario de ambiente parálico con producción de carbón, influenciado por las mareas, sistemas fluviales y vulcanismo del arco de islas.
Por otra parte, un patrón de dieta mixta sugiere un probable ambiente con parches de pastos asociados a ambientes de sabanas y bosque tropical, en nuestro caso bosque basal. Lo anterior por extrapolación del mesodesgaste extendido observado en molares de C. quinniWoodburne 1996 del Sur de México y las Grandes Planicies de Estados Unidos sensu Bravo-Cuevas y Priego-Vargas (2009).
Aunque el desarrollo de sabanas y pastizales con zacates fueron comunes y están bien documentados en el Mioceno Medio (18 a 12 Ma) de América del Norte (Janis, Damuth y Theodor, 2004; Strömberg, 2002, 2004; Retallack, 2007), incluso para el sureste de México (Ferrusquía-Villafranca, 2003), donde Bravo-Cuevas y Priego-Vargas (2009) proponen que se desarrollaron ambientes tipo “sabanas tropicales”, no se puede afirmar al cien por ciento que estos ambientes con pastizales de zacates alcanzaran latitudes tan bajas como Costa Rica. Sin embargo el registro de C. quinniWoodburne 1996, supone en alguna medida la existencia pastos tropicales.
Conclusiones
Por primera vez se registra en América Central Meridional la especie Cormohipparion quinniWoodburne 1996, lo que amplía el rango de distribución paleobiogeográfica de la misma, desde el Medio Oeste de los Estados Unidos hasta el Norte de Costa Rica.
El biocrón conocido para la especie C. quinniWoodburne 1996 es Barstoviano Tardío correspondiente al Mioceno Medio, 14 a 12 Ma, por lo que se asume tal edad para los sedimentos de la Formación Venado que portan los restos de madera fosilizada asociada con el diente. Cabe aclarar que el diente no fue encontrado ¨in situ¨ aunque su estado excepcional de conservación sugiere que proviene de las proximidades, ya que no evidencia transporte excesivo. Además, no presenta rastros calcáreos, ni ningún tipo de cemento carbonatado, por lo que se puede concluir que proviene de una de las facies clásticas de la Formación Venado, común en el área dónde se halló el ejemplar.
El molar presenta una combinación de relieve alto con cúspides redondeadas, lo que es característico de una dieta que incluye material abrasivo (cf. Bravo-Cuevas y Priego-Vargas, 2009) y de allí que se pueda intuir la presencia de parches de pastos en el neotrópico del sur de América Central durante el Mioceno Medio y una dieta mixta para C. quinniWoodburne 1996.
El presente hallazgo constituye el registro más antiguo de un mamífero terrestre para Costa Rica.

