











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
El género Otodus está constituido por una serie de cronoespecies, producto de probable evolución anagenética, es decir, cuando una especie “A” da origen a una única nueva especie “B” que le sustituye en el ecosistema, produciendo incluso formas transicionales. Ello ha originado una serie de discusiones que gravitan en torno a los únicos restos fósiles que de estos grandes tiburones comúnmente se encuentran, sus dientes, notables por su gran tamaño. Por lo que los megalodontes o mejor dicho a partir de sus dientes fósiles como única evidencia tangible se han erigido varias hipótesis evolutivas y sobre todo taxones cuyos límites entre unos y otros a veces no fue tan claro. En la actualidad su problemática taxonómica, básicamente comprende dos líneas de pensamiento, muy bien resumidas por Perez, Godfrey, Kent, Weems y Nance (2019), una que sigue básicamente las ideas de Cappetta (2012) quien considera que la presencia o ausencia de aserramiento no son caracteres suficientes para determinar o garantizar un género específico y quien además propone el uso de tres subgéneros: Otodus (Otodus) para los no aserrados, Otodus (Carcharocles) para los irregularmente aserrados con cúspides accesorias y Otodus (Megaselachus) con aserramiento regular y cúspides accesorias laterales reducidas, presentes solo en estadios juveniles o que del todo no están presentes tanto en juveniles como en adultos, y en los que además el margen basal de la raíz es paralelo a la línea basal del cuello lingual que separa la corona de la raíz. En esta última categoría se incluyen las especies Otodus (Megaselachus) chubutensis (Ameghino 1901) y Otodus (Megaselachus) megalodon (Agassiz 1835).
La segunda línea de pensamiento, mantiene un único género Carcharocles para todas las formas, tanto las que presentan aserramiento como para las que no y que consideran que Carcharocles chubutensis y Carcharocles megalodon evolucionaron a partir de Carcharocles angustidens, donde Carcharocles chubutensis retiene las cúspides accesorias en los especímenes adultos y C. megalodon no, sin embargo los juveniles de C. megalodon pueden presentar cúspides accesorias (cf. Applegate y Espinosa-Arrubarrena, 1996; Pimiento, Ehret, MacFadden y Hubbell, 2010) a manera de cúspides vestigiales según lo deducen Perez, Pimiento, Hendy, González-Barba y MacFadden, 2017 para varios especímenes de Carcharocles megalodon de Lago Bayano en Panamá.
Purdy et al. (2001), notaron que en muchos especímenes de Carcharocles subauriculatus (otro sinónimo para Carcharocles chubutensis comúnmente utilizado por algunos autores como especie válida), los dientes anteriores perdieron las cúspides laterales accesorias, por el contrario los dientes laterales presentaban cúspides laterales muy bien desarrolladas. Esta característica variación en la presencia o ausencia de cúspides laterales a nivel genérico para Perez et al., 2019, podría estar más asociada al tipo de dieta de algunos grupos poblacionales que a una transición ontogenética, ya que se ha observado que las proporciones de perdida de las cúspides laterales varían de una secuencia dental a otra.
De acuerdo a lo anteriormente expuesto, la posición de ambas líneas de pensamiento taxonómico son igualmente válidas y debo decir que durante la redacción del presente artículo y otros anteriores y su posterior revisión, diferentes colegas nos han inducido a utilizar una u otra, pero debo decir que estas mismas personas por una u otra razón han utilizado ambas líneas en diferentes momentos, lo cual también es entendible.
En todo caso hay que decantarse por una de las 2 opciones y por ello se indicará a lo largo del escrito, como igualmente válida su sinónimo, el cual resaltaremos en negrita.
Por otra parte, es importante tener en cuenta que la especie C. chubutensis u O. (Megaselachus) chubutensis aparece en el Mioceno temprano, la cual se cree derivó de la especie Oligoceno tardía O. (Carcharocles) angustidens o Carcharocles angustidens (Agassis 1835) y que luego en algún momento del Mioceno Medio basal entre los 16,4 y 14 Ma, según lo deducido de los hallazgos en América del Norte, se dio la transición de la especie O. (Megaselachus) chubutensis a O. (Megaselachus) megalodon o Carcharocles megalodon, cuando esta última especie perdió las cúspides accesorias en los individuos adultos. Sin embargo durante la transición, los juveniles de O. (Megaselachus) megalodon del Mioceno Medio a Superior presentan cúpsides accesorias, algo que se ha observado de manera muy infrecuente en las formas del Plioceno basal (Pimiento y Balk, 2015 y Perez et al., 2019).
El objetivo del presente trabajo es describir y analizar la taxonomía de 4 ejemplares del subgénero Otodus (Megaselachus), depositados en la Colección de Fósiles de la Sección de Geología del Museo Nacional de Costa Rica bajo los acrónimos CFM-911, CFM- 1500, CFM-5398 y CFM-5399, todos procedentes del sur del Península de Nicoya, provincia de Puntarenas (Fig. 1).
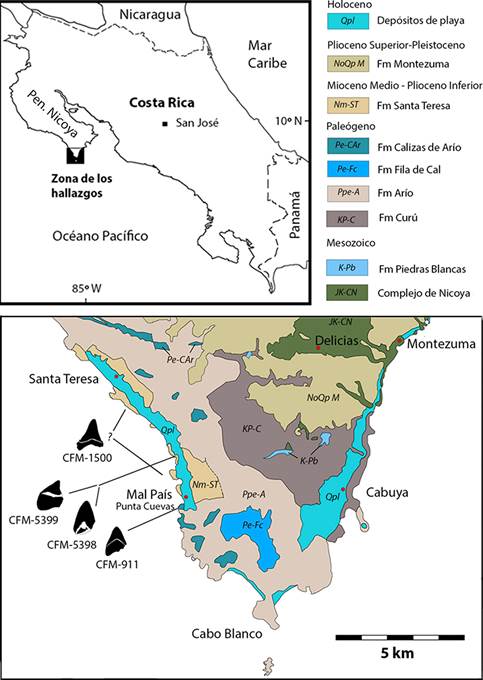
Fig. 1 Mapa geológico del extremo sur de la península de Nicoya y de ubicación de los hallazgos. Mapa geológico simplificado de la hoja Cabuya (3144-I), redibujado y adaptado de Denyer, Aguilar y Montero (2013).
Paleontología
Orden Lamniformes Berg, 1958
Familia Otodontidae Glickman, 1964
Género Otodus Agassiz, 1838
Género Carcharocles Jordan y Hannibal, 1923
Otodus (Megaselachus) chubutensis (Ameghino)
Mioceno Inferior - Mioceno Medio (20,2 - 17 Ma) Perez et al., 2019
Sinonimia: Para una detallada revisión de la sinonimia de la especie ver Laurito (2015).
- Carcharocles chubutensis (Ameghino) Perez et al., 2019
-Carcharodon megalodon (Charlesworth) var. chubutensis Ameghino en Zbyszewski y Moitinho D’Almeida, 1950.
-Procharcharodon megalodon chubutensis en Holec, Hornáček y Sýkora, 1995.
-Carcharodon subauriculatus (Agassiz) en Aguilera y Rodrigues de Aguilera, 2004.
-Megaselachus subauriculatus (Agassiz, 1838) en Laurito, Calvo, Valerio, Calvo y Chacón (2014)
-Otodus chubutensis en Maisch, Becker y Chamberlain et al., 2019.
Material: CFM-1500, un diente inferior lateral, ver figs. 2a-d.

Fig. 2 CFM-1500, diente inferior lateral de Otodus (Megaselachus) chubutensis (Ameghino (1901 a 1902)) procedente de la Formación Santa Teresa y de edad Mioceno Inferior. En vista: a. lingual, b. labial, c. perfil y d. detalle de las cúspides accesorias en norma labial.
Descripción: un diente inferior lateral derecho de 80,46 mm de altura y 67,20 mm de ancho, con corona triangular ligeramente sigmoidea y elevada de 60,52 mm (ver Cuadro 1), que se enangosta hacia el ápice, el cual está roto, y se inclina ligeramente hacia la comisura; su cara lingual es suavemente convexa y ligeramente inflada; la cara labial es ligeramente inflada pero con una convexidad mucho menos pronunciada.
Cuadro 1 Datos biométricos dentales de CFM-911 un diente bastante completo y CFM-5398 un diente del cual solo se preservó la corona, ambos determinados como Otodus (Megaselachus) megalodon (Agassiz, 1835) y el CFM-1500 correspondiente a la especie Otodus (Megaselachus) chubutensis (Ameghino).
| Ejemplar | LT | AM | BL | AC | CA | AR |
| CFM-911 | 81,79 | 60,97 (roto) | 17,13 | 59,14 | 51,90 (roto) | 58,56 (roto) |
| CFM-1500 | 80,46 | 67,20 | 18,15 | 60,52 | 68,31 | 66,48 |
| CFM-5398 | n. d. | n. d. | n. d. | 75,63 | 20,95 | n. d. |
LT longitud total del diente, AM ancho máximo, BL diámetro bucolingual, AC altura de la corona, CA ancho de la corona, AR ancho de la raíz.
La corona se extiende sobre los extremos radiculares, donde se presentan sendas cúspides accesorias o secundarias. La cúspide comisural, no abarca toda la extensión del lóbulo radicular y presenta tres cúspides diferenciadas sutilmente aserradas, la cúspide accesoria mesial parcialmente preservada, está constituida por un único elemento redondeado.
Los filos de la corona son fuertemente aserrados, e incluso algunas de las cúspides menores que conforman el aserramiento están dispuestos en pares que comparten una misma base.
La región radicular abarca un tercio de la longitud total de la altura del diente, su porción basal es cóncava y presenta una protuberancia lingual inflada pero no prominente, un cuello lingual conspicuo que se extiende hasta la base de las cúspides laterales accesorias, en tanto que la cara radicular labial es relativamente plana.
Discusión
El diente CFM-1500, corresponde al ejemplar mejor preservado, hasta ahora hallado en Costa Rica de Otodus (Megaselachus) chubutensis, aunque muestra evidencia de que estuvo sujeto a procesos bioerosivos de previo a su sepultamiento definitivo, como parte de los procesos tafonómicos, los ichnofósiles más evidentes son algunas perforaciones circulares en la cara labial de la raíz, incluso se observa un pequeño tubo, pero no se puede asegurar que hallan sido hechas por Entobia sp., son similares a las ilustradas por Maisch et al., 2019 para ejemplares de Otodus (Megaselachus) chubutensis y Otodus (Megaselachus) megalodon.
Por otra parte, se considera que se trata de un diente inferior porque los dientes inferiores de Otodus (Megaselachus) chubutensis son similares en cuanto a tamaño y forma a los superiores, pero la principal característica de los inferiores es que la corona es relativamente más angosta (Kent, 2018).
Su longitud de 80,46 mm es relativamente grande, lo que sugiere, pertenecía a un individuo probablemente adulto, sobre todo si se tiene en cuenta que se trata de un diente lateral posterior y que la altura máxima, hasta hoy reportada para un diente anterior de Otodus (Megaselachus) chubutensis es de 130 mm sensu Case (1980). Otros caracteres que permiten identificar la especie, son las relativamente pequeñas cúspides accesorias comparadas con el tamaño de la corona; en especies como Otodus (Carcharocles) angustidens del Oligoceno, las cúspides secundarias son comparablemente más grandes en relación al tamaño de la corona y el aserramiento en general es más fino en la base de la corona y se vuelve muy grueso hacia el ápice, mientras que en la especie Otodus (Megaselachus) chubutensis, a pesar de que el aserramiento es muy conspicuo, también es muy uniforme e incluso se vuelve ligeramente más fino hacia la cúspide.
Distribución paleobiogeográfica
Otodus (Megaselachus) chubutensis (Ameghino, 1901 a 1902) ha sido ampliamente registrado en el Mioceno Temprano de Europa (Leriche, 1926, Zbyszewski y Moitinho D’Almeida, 1950, Menesini, 1968 y 1974, Holec, Hornáček y Sýkora., 1995, Marsili, Carnevalle, Danese, Bianucci y Landini, 2007). También, en África, en el Mioceno Temprano de los lechos de Moghra en la depresión de Qattara en Egipto (Cook, Murray, Simmons, Attia y Chatrath, 2010) y en Asia en el Mioceno Inferior de la India (Sahni y Mehrotra, 1981). En América del Norte se ha registrado en el Mioceno Inferior de Pollack Farm en Delaware (Purdy, 1998) y Calvert Cliffs en la Bahía de Chesapeake en Maryland (Visaggi y Godfrey, 2010; Kent, 2018), y en las Formaciones Trent y Pungo River en Lee Creek Mine, North Carolina (Case, 1980, Purdy et al., 2001, Perez et al., 2019), todas en Estados Unidos; en México la especie se registra en el Mioceno Inferior de la Península de Baja California y en la Formación El Encanto en el Estado de Veracruz (Guzmán-Guzmán, González-Barba, Sánchez-Romero y Romero-Zárate, 2014, Jiménez-Castañeda, 2017). En América del Sur Otodus (Megaselachus) chubutensis (Ameghino, 1901 a 1902) ha sido registrada en el Mioceno Inferior de las formaciones Chilcatay en Ica, Perú (Alván, Apolín y Chacaltana., 2007) y Pirabas en Brasil (Costa Richter, De Toledo y Moraes-Santos, 2009, Aguilera et al., 2017); en el Mioceno Temprano de las Formaciones Castillo y Cantaure, y la península de Paraguaná, en Venezuela (Aguilera y Rodrigues de Aguilera, 2004, Aguilera, 2010). También, en la Formación Uitpa en el Mioceno Temprano de Colombia (Carrillo-Briceño et al., 2016). En el Caribe dicha especie ha sido descrita en el Mioceno Inferior de Cuba (Iturralde-Vinent Hubbell y Rojas, 1996). 1996). En América Central ha sido recientemente registrado en el Mioceno Inferior de Panamá, en la Formación Culebra por Pimiento et al., (2013a) y el Mioceno Inferior de Costa Rica en la localidad de Tres Equis de Turrialba, en sedimentos de la Formación Río Banano como Megaselachus subauriculatus (Agassiz, 1838) (Laurito et al., 2014).
Paleoecología
De acuerdo con la distribución paleogeográfica registrada para Otodus (Megaselachus) chubutensis (Ameghino, 1901 a 1902) y los sedimentos en que sus restos han sido hallados, podemos deducir que fue un potente nadador nectónico y epipelágico, habitante primordial de las plataformas continentales e insulares, tanto en aguas someras sublitorales como en ambientes oceánicos de las regiones tropicales y subtropicales. Sus dientes de relativo gran tamaño y fuertemente aserrados, sugiere la posibilidad de que se alimentara de grandes peces, reptiles y mamíferos marinos, ya fuera como un cazador activo o como un carroñero oportunista, comparable a lo observado para el nicho ocupado en la actualidad por Carcharodon carcharias (cf. Compagno, 2001).
Otodus (Megaselachus) megalodon (Agassiz, 1835)
Mioceno Medio - Plioceno temprano (15.9-3.6 Ma), Boessenecker et al., 2019 a Plioceno tardío (2,6 Ma) cf. Pimiento y Clements, 2014.
Sinonimia: A continuación se indican algunas de las sinonimias más comunes utilizadas tradicionalmente. Para una exhaustiva revisión se recomienda consultar a Cappetta (1970), Antunes y Jonet (1970), Laurito (1999) y Mikuž Šoster, Stare y Sukič Prekmurski, 2015.
- Carcharocles megalodon (Agassiz, 1843) en Müller, 1969.
-Procarcharodon megalodon (Agassiz) en Longbottom, 1979.
-Carcharodon megalodon Agassiz en Bendix-Almgreen, 1983.
-Megaselachus megalodon (Agassiz, 1835) en Schultz, 2013
-Carcharocles megalodon (Agassiz, 1837) en Toscano, 2016.
-Otodus (Megaselachus) megalodon Agassiz 1835 en Trif y Codrea, 2017.
-Otodus megalodon (Agassiz, 1835) en Maisch, Becker y Chamberlain., 2018.
Glikman (1964) consideró la pérdida de las cúspides laterales en Otodus megalodon, suficientemente significante como para incluir a esta especie en el género Megaselachus. Sin embargo, los juveniles de Otodus (Megaselachus) megalodon pueden retener las cúspides laterales, lo que es considerado una variación ontogenética (Applegate y Espinoza-Arrubarena, 1996).
Material: 3 restos dentales, el CFM-911, un diente superior lateral próximo a la sínfisis (figura 3a-c); el CFM-5398 correspondiente a una corona de un diente antero lateral inferior figura 4.1) y el CFM-5399 correspondiente a un fragmento distal basal de gran tamaño (figura 4.2).

Fig. 3.CFM-911, diente antero-lateral superior de un juvenil de Otodus (Megaselachus) megalodon (Agassiz, 1835), procedente de la localidad de Malpaís. En vista: a. lingual, b. labial y c. perfil.

Fig. 4 1. CFM-5398, corona de un diente antero lateral inferior de un individuo juvenil y 2. CFM-5399, fragmento distal y basal de un diente antero-lateral de posición incierta, ambos de la especie Otodus (Megaselachus) megalodon (Agassiz, 1835), procedentes de la localidad de Malpaís. En vista: a. labial y b. lingual.
Descripción: El ejemplar CFM-911 es un diente superior lateral próximo a la sínfisis y de lateralidad incierta de 81,79 mm de altura y 60,97 mm de ancho, de aspecto triangular ligeramente inflado, la corona es elevada con una altura 59,14 mm, su cara labial es cóncava pero poco pronunciada, mientras que la cara lingual es ligeramente convexa. La raíz es alta y su cara lingual presenta una protuberancia lingual amplia pero no sobresaliente, y limita a la corona por un prominente cuello en forma de “chevron” amplio, mientras que la cara labial limita a la corona por un delgado cíngulo de color más oscuro que se ensancha ligeramente hacia los extremos radiculares. La corona en su cara labial presenta una hendidura poco profunda en su porción mediana que se vuelve indefinida hacia su mitad superior y que divide al diente de manera simétrica. Los filos de la corona son finamente aserrados y uniformes.
El ejemplar CFM-5398 corresponde a una corona casi completa de 75,63 mm de alto y un ancho de 20,95 mm en su base, la cara lingual es ligeramente convexa en tanto que la cara lingual es cóncava e inflada y se observa un “chevron” angosto y alto.
El ejemplar CFM-5399 corresponde a un fragmento lateral basal de un diente aparentemente inferior, en el que se preservó el lóbulo radicular y una porción de la corona. El lóbulo radicular preservado tiene una altura de 47,32 mm y un ancho de 36,27 mm, y el fragmento de corona tiene una altura de 42,27 mm. En la cara lingual se observa parte del “chevron” que marca el límite entre la corona y la raíz.
Distribución paleobiogeográfica y edad de los registros
Otodus (Megaselachus) megalodon (Agassiz, 1835) (Agassiz, 1835) se caracteriza por tener una distribución cosmopolita, pantropical y pan-subtropical, por lo que el registro de sus hallazgos resulta ser un hecho frecuente en las secuencias marino someras del Neógeno a nivel planetario (Antunes y Jonet, 1970 y Capetta, 2012).
En el Caribe se ha descrito para el Mioceno Medio y Superior, y Plioceno de Cuba (Sánchez Roig, 1920, Iturralde-Vinent et al., 1996) y La Española (Leriche 1938, Kruckow y Thies, 1990); Mioceno de la isla de Barbados (Ray, 1964); Mioceno Medio de Puerto Rico (Nieves-Rivera, Ruíz-Yantín y Gottfried, 2003), Mioceno Medio del Caribe de Venezuela en la Formación Cantaure, Plioceno Temprano de la Formación Paraguaná (Aguilera, García y Cozzuol, 2008, Carrillo-Briceño et al., 2016) .
En América Central ha sido descrito para el Mioceno Tardío de las Formaciones Gatún (White, 1955, Gillette, 1984, Pimiento, Ehret, MacFadden y Hubbell, 2010, Pimiento et al., 2013b), Chagres (Carrillo-Briceño et al., 2015) y Chucunaque en la localidad de Lago Bayano (Perez et al., 2017) en Panamá.
En Costa Rica Carcharocles megalodon (Agassiz, 1835) fue inicialmente descrito para la Formación Pacacua (Crosby, 1945) y luego para la Formación Roca Carballo (Kruckow y Thies, 1990) ambas de edad Mioceno Medio, para el Mioceno Superior-Plioceno Inferior de la Formación Uscari (Laurito, 1999). Los presentes hallazgos confirman el registro fósil de Carcharocles megalodon (Agassiz, 1835) en el sur de la Península de Nicoya, procedente muy probablemente de los sedimentos más tardíos de la Formación Santa Teresa de edad Mioceno Superior-Plioceno Inferior.
Paleoecología
Otodus (Megaselachus) megalodon (Agassiz, 1835) fue un potente nadador oceánico de allí su distribución cosmopolita; habitante de ambientes neríticos tanto continentales como insulares, ocupó como tiburón la categoría de depredador máximo en la cadena alimentaria (Laurito, 1999) debido a su gran tamaño, con una media de 17,9 m en el Mioceno Medio y 10,2 m en el Mioceno tardío (sensuPimiento y Balk, 2015). Su dieta se basó principalmente de cetáceos (delfines y ballenas de mediano y pequeño tamaño); además de pinnípedos, sirenios, tortugas y grandes peces (Cigala Fulgosi, 1990, Aguilera y Rodrigues de Aguilera, 2004, Aguilera et al., 2008, Collareta et al., 2017, Godfrey y Altman, 2005, Godfrey, Ellwood y Verdin, 2018).
Discusión y conclusiones
El ejemplar CFM-1500, fue ingresado al Museo Nacional de Costa Rica, como parte de un decomiso realizado en el Aeropuerto Internacional Daniel Oduber Quirós a turistas que visitaron el sur de la Península de Nicoya. El diente en sí, presenta una preservación, una mineralización y color propios de los dientes de Otodus (Megaselachus) chubutensis procedentes de la Formación Santa Teresa, en la Provincia de Puntarenas y de las secuencias sedimentarias con rango de edad Mioceno Inferior a Medio. Al no haber en la península ninguna otra formación con ese rango de edad y registro de dientes de tiburón de esta especie, asumimos que el ejemplar procede de la Fm. Santa Teresa (ver los ejemplares ilustrados y analizados por Laurito, 2015 donde coinciden el tipo de preservación y coloración).
El CFM-911 es un ejemplar de Otodus (Megaselachus) megalodon con 81,79 mm de alto y 60,97 mm de ancho, estas dimensiones caen dentro del rango de los dientes laterales de juveniles observados para la Formación Bone Valley de edad Plioceno Temprano (ver figura 3 de Pimiento et al., 2010), de allí que se puede concluir que el ejemplar es un juvenil lateral anterior.
El ejemplar CFM-911 fue recuperado por el Sr. William Granados R., en las coordenadas lambert 175,75 N y 410,8 W, en el lecho de una quebrada intermitente luego de una lluvia intensa en Punta Cuevas. En esa localidad, los únicos sedimentos aflorantes de los cuales pudo provenir el diente fósil son los depósitos de playa Qpl (sensu Denyer, Aguilar y Montero, 2013), pero la edad de estos sedimentos discrepa del rango de distribución conocido para Otodus (Megaselachus) megalodon, por lo que es posible que se trate de material fósil retrabajado, procedente de las secuencias más tardías de la Formación Santa Teresa de edad Mioceno Superior-Plioceno Inferior, aflorantes tierra adentro a unos cuantos cientos de metros de la playa.
El espécimen CFM-5399 a pesar de ser un fragmento distal, permite extrapolar sus dimensiones y hacer un cálculo conservador de su ancho radicular, el cual pudo haber tenido un aproximado de 125 mm y una altura de 130 a 140 mm. Este fragmento puede corresponder al diente de Otodus (Megaselachus) megalodon (Agassiz, 1835) más grande, hasta ahora hallado en el país.
Los ejemplares CFM-5398 y CFM-5399 fueron recuperados por el Sr. Daniel Gutiérrez Solano y vecino de playa Carmen, sobre la plataforma de abrasión marina que allí aflora, misma que está constituida por estratos sedimentarios de la Fm. Santa Teresa, en el año 2019, en las coordenadas 9 37'03''N y 85 08'48,5'' W. Su ubicación sugiere que ambos ejemplares provienen de dicha plataforma, producto de la erosión provocada por el oleaje durante la marea alta.
Por último, en el país se han registrado 3 especies de tiburones Otodontidae gigantes hasta el momento, Otodus (Carcharocles) auriculatus procedente de la Unidad Cajón de la Formación Fila de Cal de edad Eoceno Superior cf. Henningsen, 1966 y Kruckow y Thies, 1990; Otodus (Megaselachus) chubutensis para las formaciones Santa Teresa y San Miguel del Mioceno Inferior y Otodus (Megaselachus) megalodon para el Mioceno Medio de Roca Carballo (Kruckow y Thies, 1990), Mioceno Superior - Plioceno Inferior de la Formación Uscari (Laurito, 1999) y los presentes hallazgos que confirman su registro en sedimentos marinos sublitorales a litorales asociados a la Fm. Santa Teresa.

