











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroduction
The Republic of Guatemala has an area of almost 109,000 km2, and, with a population of about 16 million people, it is the most populous country in Central America. Today, Guatemala is also a country remarkable for its biodiversity (at least 1200 living species of animals and 9000 plant species) and its ecological and topographic diversity, which encompasses seacoasts, mangrove forests, wetlands, forest, lakes, 100 rivers and mountains as high as 4211 m.
The complex geology of Guatemala has been the focus of much recent study, especially because of its tectonic position relative to plate boundaries (Fig. 1). However, relatively little study of the sedimentary rocks and fossils of Guatemala has been undertaken ( Bundschuh & Alvarado, 2007).
Indeed, the vertebrate paleontology of Guatemala has only been studied and published upon sporadically, since the first published scientific report of a vertebrate fossil from Guatemala in 1926. Until recently, few fossil vertebrates were documented from Guatemala (cf. Lucas et al., 2007), but that has begun to change during the last decade (Mead et al., 2012; McDonald & Dávila A., 2017; Dávila et al., 2019). Here, we review the vertebrate paleontology of Guatemala based on published records and fossils in museum collections. Our goal is to place the vertebrate-fossil record of Guatemala in its Central American context and to evaluate its biogeographic significance.
Institutional Abbreviations
The following institutional abbreviations are used in this paper: AMNH = American Museum of Natural History, New York, New York, USA; ME = Museo de Estanzuela, Estanzuela, Guatemala; NMMNH = New Mexico Museum of Natural History, Albuquerque, New Mexico, USA; USAC = Museo de Historia Natural, Universidad de San Carlos de Guatemala, Ciudad de Guatemala, Guatemala.
Geological Timescale
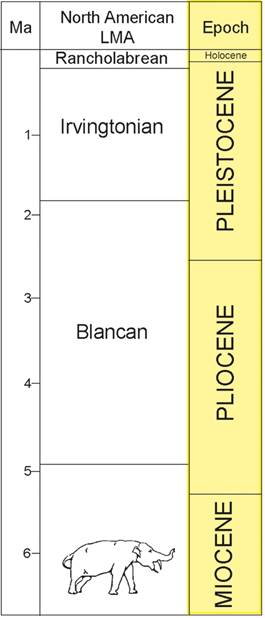
We assign ages to vertebrate fossils from Guatemala based primarily on the North American land-mammal “ages” (NALMAs), as characterized and correlated to the late Cenozoic epochs by Tedford et al. (2004) for the Miocene and by Bell et al. (2004) for the Pliocene-Recent. We use the redefinition of the Pleistocene, which lowers the Pleistocene base to about 2.6 Ma. Thus, the NALMAs relevant to the Guatemalan vertebrate fossil record are: (1) Hemphillian: Late Miocene-Early Pliocene, ~ 4.7-9.0 Ma; (2) Blancan: Early Pliocene-Early Pleistocene, ~1.8-4.7 Ma; (3) Irvingtonian: Early-Middle Pleistocene, ~ 200,000 years-1.8 Ma; and (4) Rancholabrean: Late Pleistocene, ~ 10,000-200,000 years (Fig. 2).
Geological Context
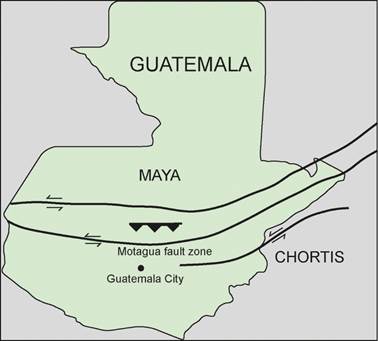
Guatemala is at the center of a region of actively converging plates and transform faults (e.g., Weyl, 1980; Dengo & Case, 1990; Nagle et al., 1994; Stanford University, 2004-2005; Bartole et al., 2019) (Fig. 1). Thus, the southwestern coast of Guatemala adjoins the Middle American trench where the Cocos plate is being actively subducted beneath the North American and Caribbean plates. The boundary between the Caribbean and North American plates cuts through central Guatemala along what has variously been called the Motagua valley fault zone (Weyl, 1980), the Motagua suture zone (Dengo & Case, 1990), or the Polochic and Motagua fault system (Bartole et al., 2019). It divides two tectonic blocks-the Maya block to the north and the Chortís block to the south (Fig. 1)
Guatemala is thus composed of parts of three tectonic plates: North American, Caribbean and Cocos. Northern Guatemala is on the North American plate, and the exposed bedrock is mostly folded sedimentary rocks. The Cocos plate plunges into the mantle off of the Pacific coast in a subduction zone, the Middle American trench. This led to the deposition of the Cenozoic (largely Quaternary) sedimentary and volcanic rocks that cover most of southern Guatemala. The northwestern edge of the Caribbean plate is being pinched from the south by the Cocos plate and truncated to the north by the left lateral strike-slip boundary with the North American plate (Fig. 1).
Guatemala is typically divided into four geological provinces (Weyl, 1980; Nagle et al., 1994; Stanford University, 2004-2005; Bundschuh & Alvarado, 2007; Cosillo, 2008): (1) Quaternary sediments along the Pacific coastal plain; (2) a volcanic province inboard of and essentially parallel to that plain that was created by Cenozoic subduction of the Cocos plate beneath the Caribbean plate; (3) an east-west metamorphic belt and the sutural zone of the Motagua Valley fault zone across the central portion of the country, which is the boundary of the North American and Caribbean plates (and the Maya and Chortis blocks); and (4) sedimentary deposits of variable thickness and extent on the Maya block in the northern part of the country (e.g., Millan, 1985; Alvarado et al., 2007).
The Maya block has an extensive sedimentary cover of Mesozoic rocks that include substantial beds of marine limestone and evaporites. The erosion and dissolution of these strata accounts for the karst topography of parts of northern Guatemala. The Petén lowland of northern Guatemala is continuous with the Mesozoic-Cenozoic deposits of the Campeche-Yucatán basin of Mexico (Alvarado et al., 2007).
The Chortís block is the northernmost part of the Caribbean plate. Active tectonism on the block has produced various late Cenozoic grabens, some with more than 2 km of sedimentary fill, as well as the large basin of northwestern Petén. Deposits in one of the late Cenozoic grabens yield the oldest Guatemalan vertebrate fossils, of late Miocene age. But, the vast majority of Guatemalan vertebrate fossils are from late Pleistocene sedimentary deposits, some volcaniclastic, in the southern half of the country (Dávila et al., 2019) (Fig. 3).
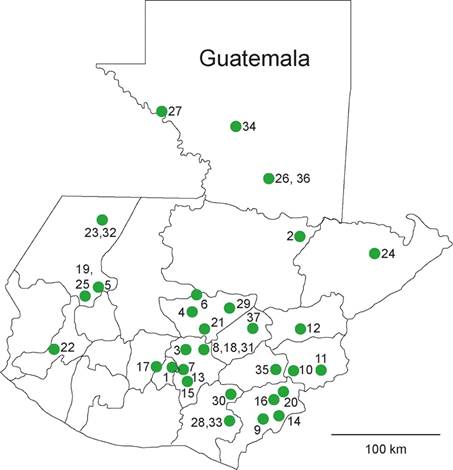
Fig. 3 Vertebrate fossil localities in Guatemala discussed in the text. Localities are: 1 = Cerro de la Virgen; 2 = Chahal; 3 = Chinautla; 4 = Chiticoy, Rabinal; 5 = Chivacabé; 6 = Chixoy; 7 = Ciudad Real; 8 = Colonia Tecun Umán; 9 = El Adelanto; 10 = El Rosario-Ipala; 11 = Área de El Trifinio; 12 = Estanzuela; 13 = Finca San Ramón; 14 = Hacienda El Peñol; 15 = Hacienda San Carlos; 16 = Jutiapa; 17 = Kilómetro 31, Carretera CA-1 Occidente; 18 = Los Vados; 19 =Malacatancito; 20 = Paso Calandrias; 21 = Quebrada Pacoy; 22 = Quetzaltenango, Tívoli; 23 = Río Canbalam; 24 = Río Carboneras; 25 = Río Chinacá; 26 = Río La Pasión; 27 = Río Usumancita; 28 = Roca Madariaga; 29 = Salamá; 30 = San Rafaelito; 31 = Sanguayabá; 32 = Santa Cruz Barillas; 33 = Santa Rosa; 34 = Sayaxché; 35 = Songotongo; 36 = Treasure Island, Petén; 37 = Tulumaje y Tulumajillo.
Unfortunately, most of the records of fossil vertebrates reported from Guatemala lack geological context and stratigraphic data, or data on their preservation (taphonomy). Such data have been acquired in Costa Rica and Nicaragua (Lucas & Alvarado, 2017, and references therein), for example, through site visits, talking to the people that discovered the fossils, and post-discovery reconstruction of the geological context, even though it may have needed to be made many years after the original discovery. We could not recover most of the geological, stratigraphic or preservational context of Guatemala’s fossil vertebrates for the present paper except based on observations and analyses available in published sources (e.g., Mead et al., 2012; Dávila et al., 2019).
Some History
Colón (2015, p. 86-103) provided a very detailed and extensive review of the history of discoveries and studies of the fossil vertebrates of Guatemala, so our historical review is brief. He divided that history into three phases, 1690-1945, 1946-1977 and 1978-2010. However, we identify two phases divided by the date 1926, when the first scientific publication appeared on a fossil vertebrate from Guatemala. Thus, although the first published reference to vertebrate fossils from Guatemala was very long ago, 1690, scientific description of fossil vertebrates from in Guatemala began in the 1920s, when Osborn (1926) presented the first published report of a fossil vertebrate from Guatemala, a proboscidean molar, that was part of a purchased collection (see below).
Three famous students of fossil vertebrates from the United States collected in Guatemala at various times between the late 1940s and early 1970s: Bryan Patterson (1909-1979) of Harvard University, Barnum Brown (1872-1963) of the American Museum of Natural History and C. Lewis Gazin (1904-1995) of the Smithsonian Institution. Brown worked along the Río La Pasión during the late 1940s and early 1950s. According to Mook (1959), Brown worked there from 1948 through 1953, but Dávila et al. (2019) state that he worked there in 1940-1945 (see Brown, 1956, for a colorful account of some of this fieldwork), and he conducted the first extensive excavation of Pleistocene fossil mammal from the vicinity of Estanzuela in 1950. However, according to Museo de Estanzuela records and local news media, the German geologist Karl Sapper (1866-1945) first brought the fossil mammals found in the vicinity of Estanzuela to scientific attention. Although in Guatemala’s local media it is said that Sapper discovered the fossil remains at Estanzuela in the 1940s, the reality is that he was already retired in Germany by then, putting his large number of documents in order and did not return to Central America (Alvarado & Denyer, 2012). Possibly in Estanzuela, the fossil locality is referred to with Sapper’s name as a recognition of his extensive research and contributions to the geology of Guatemala. Many years later, Gazin visited Guatemala in 1962 to examine various sites, including Estanzuela, but he apparently did little or no collecting (Gazin, 1965). And, Bryan Patterson worked with Roberto Woolfolk at Estanzuela in 1971-1972.
Roberto Woolfolk Saravia (1928-2000) was born in Honduras to an American father and Guatemalan mother. He was trained as an engineer and had a military career. His interests in archaeology and paleontology induced him to excavate Pleistocene mammals in the Estanzuela area. This led him to found the museum at Estanzuela in 1974, which is now named the Museo de Paleontología y Arqueología Roberto Woolfolk Saravia in his honor (Fajardo, 2004) (Fig. 4). The museum in Estanzuela is, at present, the most outstanding paleontological museum in Central America.

Jorge A. Ibarra (1920- ) was another Guatemalan scientist who made important contributions to vertebrate paleontology. Born in Quetzaltenango, Ibarra came from a family of naturalists and was also trained in taxidermy. In 1950, he founded the Museo Nacional de Historia Natural in Guatemala City and served as its director from 1950 till 1996. In 1988, the Guatemalan government named the museum the Museo Nacional de Historia Natural “Jorge A. Ibarra” in his honor. Ibarra published 14 books on natural history topics, including his 1980 volume that listed all the Guatemalan vertebrate fossil localities and fossils known to him.
However, collecting and study of Guatemalan fossil vertebrates during the latter half of the 20th century was sporadic and mostly undertaken by archaeologists (Vinson, 1962; Woodburne, 1969; Thurmond, 1973; Cano & Schuster, 1995; García & Ericastilla, 1995).
Work in this millennium has added substantially to our knowledge of Guatemalan fossil vertebrates. This is evident in the important publications of Mead et al. (2012), McDonald & Dávila (2017) and Dávila et al. (2019), and we add to these reports this review paper.
Fossil Vertebrate Localities
The oldest sedimentary rocks in Guatemala are of Silurian age, and the oldest fossils are of Carboniferous-Permian age (Schuster & Bonis, 2008). These are marine fossils of the typical late Paleozoic shelly benthos, mostly brachiopods, crinoids and bryozoans. There are also Jurassic, Cretaceous and Paleogene marine fossils known from Guatemala, particularly microfossils of foraminiferans and radiolarians (Schuster & Bonis, 2008). The oldest vertebrate fossils from Guatemala are of Miocene age.
Here, we review all of the fossil vertebrate localities known to us in Guatemala (Fig. 3). Most of the vertebrate fossils from Guatemala come from a diversity of Pleistocene deposits about which little is known in terms of their sedimentary history (e. g., García & Ericastilla, 1995; Schuster & Bonis, 2008; Colón 2015; Dávila et al., 2019). Many of these localities also lack adequate documentation (descriptions, illustrations) of the fossil vertebrates that have been reported from them, and the whereabouts of specimens to voucher many of these reports is unknown. Therefore, although included here for the sake of completeness, we stress that most reports of fossil vertebrates from Guatemala need documentation before they are considered in scientific analysis. Only six localities of late Pleistocene age in Guatemala yield a documented diversity of about 4-10 mammal genera (Table 1).
Table 1 Six localities of late Pleistocene age yield a documented diversity of about 4-10 mammal genera, and those localities and genera are listed here, with their North or South American origin indicated.
| Genus | Chivacabé | Ciudad Real | El Rosario | Estanzuela/ Zacapa | Río La Pasión | San Rafaelito | North/South America |
| Canis | - | - | - | - | X | - | north |
| Cuvieronius | X | X | X | X | X | X | north |
| Equus | X | X | - | X | X | - | north |
| Eremotherium | - | X | X | X | X | X | south |
| Glyptotherium | X | - | X | X | X | X | south |
| Mammuthus | - | - | - | X | X | - | north |
| Mixotoxodon | - | X | X | X | X | X | south |
| Neochoerus | - | - | - | X | X | - | south |
| Odocoileus | X | X | - | X | X | X | north |
| Paramylodon | - | - | - | - | X | - | south |
Colón (2015) presented the most detailed locality data available on these sites, and he also listed (p. 79-82) 24 more vertebrate fossil occurrences “sin lugar exacto” (without exact location) not listed here. Dávila et al. (2019) presented map coordinates and some geological context for some of the most prolific fossil-vertebrate sites in Guatemala.
In our review, the localities, some of which encompass multiple localities in a limited geographical area, are listed in alphabetical order. Many of the reports are of “mastodon(s),” and these are likely records of the gomphothere Cuvieronius.
Cerro de la Virgen
Located in the Municipio de Mixco, this locality yielded a “mastodon” fossil according to Colón (2015).
Chahal
Lucas et al. (2016) described and illustrated a lower jaw of a proboscidean from Pleistocene deposits near Chahal (Fig. 5). This lower jaw lacks lower tusks, has a short symphysis that is not downturned, its molars have relatively simple crowns, and its m3s have 4.5 lophids. Therefore, it displays features diagnostic of Cuvieronius hyodon, to which it was assigned.

Chinautla
Chinautla encompasses multiple localities, one of Miocene age and the others of late Pleistocene age. Colón (2015) noted that the first discoveries of vertebrate fossils at Chinautla were of a giant sloth, “mastodont” and horse reported in 1876 by Pedro Ibarra (an ancestor of Jorge Ibarra) of the “Museo Nacional de la Sociedad Económica.” Osborn (1926, p. 12, fig. 10; also see Osborn 1936, p. 432) named the new species Serridentinus guatemalensis for an incomplete M3 of a gomphothere (Fig. 6) from Chinautla (Departamento de Guatemala). According to Osborn, this fossil was purchased in 1910 as part of the Emil Mosonyi collection, but he provided no more detailed provenance. This tooth has a width of 92 mm, cuspidate and anteriorly canted lophs and at least one pillar between the lophs. It clearly belongs to Gomphotherium (Lucas et al., 2007; Lucas & Alvarado, 2010), and S. guatemalensis is likely a synonym of G. hondurensis (sensuLucas & Morgan, 2008; Carranza-Castañeda, 2018). However, strictly speaking, this incomplete molar is not diagnostic of a species of Gomphotherium, so we consider G. guatemalensis to be a nomen dubium (nomen vanum). This is presumably a late Miocene record.
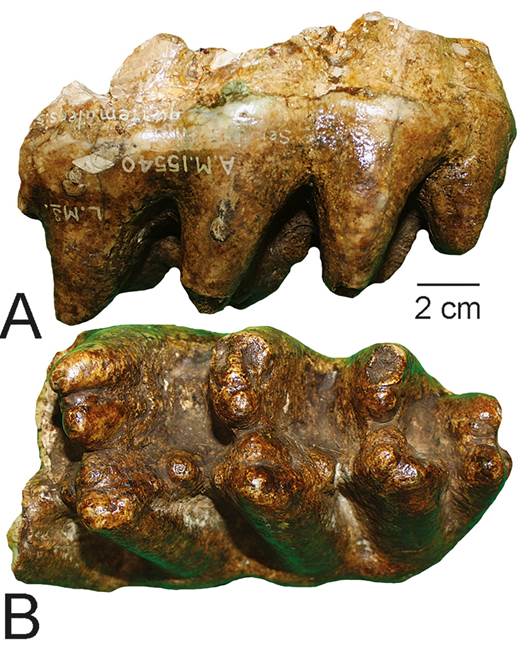
Fig. 6 Holotype of Serridentinus guatemalensis (= Gomphotherium hondurensis) from Chinautla, Guatemala, incomplete left M3, in labial (A) and occlusal (B) views. The specimen is catalogued as AMNH 15540.
Gazin (1965) reported Eremotherium and a mandible of a “mastodonte” from Chinautla. Colón (2015) states that Gazin attributed the mandible to Cuvieronius hyodon or to “Haplomastodon waringi”, Ibarra’s (1980) listing of Cuvieronius or Haplomastodon from Chinautla is likely a reference to this fossil.
From the old city of Chinautla, McDonald & Dávila A (2017, fig. 6) documented an incomplete lower molar of Mammuthus columbi (Fig. 7A-B) that they say was found with fossils of Equus sp. and cf. Euceratherium sp. At Senahu II, in the vicinity of Chinautla, García & Ericastilla (1995) reported (without documentation) fossils of Eremotherium rusconii (= E. laurillardi: Cartelle & De Iuliis, 1995) and Cuvieronius hyodon. These are all late Pleistocene (Rancholabrean) records if the taxonomic identifications are correct. Mead et al. (2012) listed only a “gomphothere” as a valid record from Chinautla. Dávila et al. (2019) document specimens of Equus and Euceratherium from Chinautla. A late Pleistocene genus-level listing from Chinautla thus includes Cuvieronius, Equus, Eremotherium, Euceratherium and Mammuthus, though most of these taxa need further documentation and all are not from a single locality, so Chinautla is not listed in Table 1.

Chiticoy, Rabinal y Las Vegas Chayan
From the Quebrada Pacux near the village of Chiticoy, Rabinal y Las Vegas (Departamento de Baja Verapaz), García & Ericastilla (1995, p. 115) mention a proboscidean fossil, “probably Holocene.” However, that age assignment seems unlikely given that there are no documented Holocene records of proboscideans in Central America (e.g., Lucas & Alvarado, 2010; Prado et al., 2015). Colón (2015) calls the Chiticoy locality “Quebrada Pacoy,” and notes that similar proboscidean material was found nearby at Las Vegas Chayan. Mead et al. (2012, table 2) list only a “proboscidean” from Chiticoy.
Chivacabé
This site was first excavated by archaeologists in the 1970s. Ibarra (1980) listed mastodont and horse from this site, which he called the “Sitio Villatoro.” García & Ericastilla (1995) list glyptodont, giant armadillo, mastodont, Tayassu tajacu and Odocoileus from this locality.
Mead et al. (2012) described the fossil mammal assemblage from this site (also see an earlier report by Cano & Schuster, 1995), which is in the western highlands of Guatemala and is bracketed by radiocarbon dates of 12,920 and 15,700 calendar years BP. Taxa documented are Cuvieronius hyodon, Equus, Glyptotherium, and Odocoileus cf. O. virginianus (Table 1). Dávila et al. (2019, p. 278) state that the fossils here are in a “lahar of reworked ash,” and Yelacic (2010) and Lohse et al. (2012) provide more details on the geology.
Chixoy
In Alta Verapaz at Chixoy, García & Ericastilla (1995) report that a glyptodont was discovered and destroyed during construction of a hydroelectric dam. A dentary fragment with a m3 of Cuvieronius hyodon from Chixoy was illustrated in a newspaper article (Dominical Prensa Libre, 13 February 2005, p. 23).
Ciudad Real
Colón (2015) states that this locality, which he calls “Quebradas del Frutal,” was discovered in the early 1970s. Thurmond (1973), in an abstract, first reported on this locality and listed the occurrence of Cuvieronius, Equus, Eremotherium, Mixotoxodon (“Toxodon”) and Odocoileus. García & Ericastilla (1995) also mention fossils of sloth, toxodont, mastodont, horse and deer from this locality. Dávila et al. (2019) call this site Barranco del Ciudad Real and list in their appendix specimens of Cuvieronius, Equus, Eremotherium, Euceratherium, Mixotoxodon, and Odocoileus (Figs. 8-10; Table 1). They state (p. 279-280) that the fossils here came from a “fine sandy lahar matrix (reworked volcanic sediments) of basaltic-andesitic composition.”

Fig. 8 Eremotherium laurillardi and Odocoileus? sp. from Ciudad Real, Guatemala. A, Eremotherium laurillardi, molariform tooth in lateral view, USAC 199/72. B-D, Ungual phalanx of E. laurillardi, USAC 345/72, in dorsal (B), lateral (C) and ventral (D) views. E-F, Odocoileus? sp., USAC 264/72, proximal part of antler in two views.

Fig. 9 Mixotoxon larensis and Euceratherium sp. from Ciudad Real, Guatemala. A-B, M. larensis, molariform tooth in two viewa, USAC 533/72. C-E, Euceratherium sp., horn core in three views, USAC 1063.

Colonia Tecún Umán
This site is located in the Municipio San Pedro Ayampuc. Colón (2015, p. 62) states a “mandíbula de mastodonte con molares” was found here.
El Adelanto
Colón (2015) calls this locality “El Sitio,” and it is located in the Municipio El Adelanto. He states that it yielded a “mastodon” fossil.
El Rosario-Ipala
Gazin (1965) mentioned the presence of the ground sloth Eremotherium at Chiquimula in eastern Guatemala. The fossil site, which they called El Rosario-Ipala in Chiquimula, was described in some detail by García & Ericastilla (1995). They reported “mastodonte,” “perezosos gigantes,” “megaterios,” a “cervido” and a “roedor.” Although there are evidently numerous fossil bones at the site (cf. García & Ericastilla, 1995, fig. 5), no specimens were illustrated or otherwise documented, though these likely represent at least records of Cuvieronius and Eremotherium. Schuster & Bonis (2008) also mention this site, as does Colón (2015), who refers to it as “Tontol” and notes a possible molar of Cuvieronius from there.
Mead et al. (2012, table 2) list megathere, gomphothere and deer from this locality. Dávila et al. (2019) list specimens of Cuvieronius, Eremotherium, Glyptotherium and Mixotoxodon.
(Área de) El Trifinio
Colón (2015) states that fossils of a turtle and of Eremotherium were found at this locality.
Estanzuela
Colón (2015) notes that Navarro (1856) first mentioned Zacapa as a vertebrate-fossil locality, claiming that fossil skeletons of “mastodontes” were collected there. Estanzuela, in the Departamento de Zacapa, is the richest and most diverse late Pleistocene mammal locality in Guatemala and the location of a museum with many Pleistocene mammal fossils on display (e.g. Gazin, 1965; Ibarra, 1980; Dávila et al., 2019; Table 1; Figs. 11-12). Barnum Brown first excavated at this late Pleistocene mammal locality with Jorge Ibarra in 1950. It was later collected by Brown and Gazin in 1955, Roberto Woolfolk and Bryan Patterson in 1971-1972, and subsequently by Woolfolk.
Dávila et al. (2019, p. 278) note that the site they call La Estanzuela is “different approximately coeval sites along the Motagua River, which today are all located in the urban area of Estanzuela.” They went on to describe the sites as from a single stratigraphic unit, several meters thick, of “fluvially transported pumitic (sic) sand and silt of volcanic origin (lahar).” Other nearby localities are Calle del Comercio in the city of Zacapa where Dávila et al. (2019) record Equus; Providencia (a glyptodont); Los Yajes (a mastodon) (Colón, 2015); and the banks of the Motagua River near Teculutlán, from which McDonald & Dávila A (2017, fig. 4) documented a lower molar of Mammuthus columbi.
Mead et al. (2012, table 2) listed the fauna of Estanzuela as glyptodont, gomphothere, megathere, tapir, horse and camel. McDonald and Dávila A (2017) list the following taxa from Estanzuela: Cuvieronius hyodon, Equus sp., Eremotherium laurillardi, Glyptotherium cf. G. floridanus, Mixotoxodon larensis, Mammuthus columbi, and Neochoerus sp. (Table 1; Figs. 7C-D, 11, 12, 13, 14).

Fig. 11 The main gallery in the Museo de Estanzuela. The mounted skeleton of Cuvieronius is from the Estanzuela area (and is a composite), but the mounted skeleton of the ground sloth Eremotherium is from Finca San Ramón in zone 6 of Guatemala City.

Fig. 12 Closer view of the mounted skeleton of the ground sloth Eremotherium from Finca San Ramón on display at the Museo de Estanzuela.
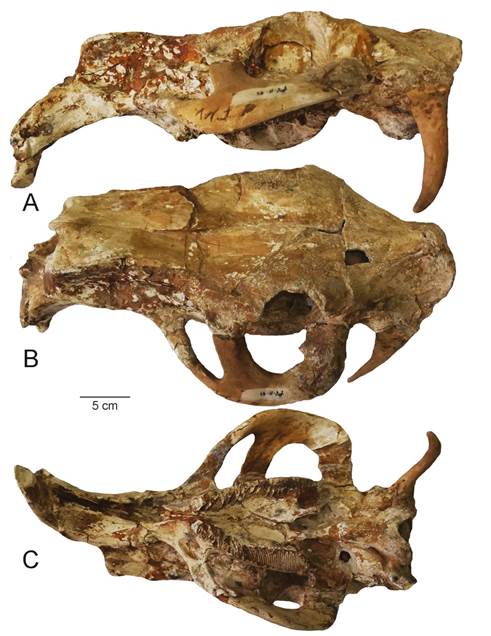
Fig. 13 Skull of Neocheorus from Estanzuela, Guatemala, in lateral (A), dorsal (B) and ventral (C) views. From locality #59, María del Transito Suarez Pinto, Estanzuela area.

Fig. 14 Cuvieronius hyodon lower jaw fragments from Zacapa, Guatemala. A-B, right dentary fragment with part m2 and m3 in occlusal (A) and labial (B) views, Zacapa. C-D, left dentary fragment with m3 in labial (C) and occlusal (D) views. From Sitio no. 2, María Navas Vida de Cabrera, Estanzuela.
McDonald & Dávila A (2017, figs. 1-3) documented three molars of Mammuthus columbi from Estanzuela. Dávila et al. (2019) listed numerous specimens of Cuvieronius and Mixotoxodon from a locality they call Barrio la Reforma, City of Zacapa. They also listed Glyptotherium.
Finca San Ramón
There are multiple fossil-vertebrate localities in Guatemala City, and we recognize Ciudad Real (see above) and Finca San Ramón as two distinct localities. Located in zone 6 of Ciudad de Guatemala, Colón (2015) states that a fossil of Eremotherium was collected at Finca San Ramón. Ibarra (1980) lists Eremotherium from zona 6 of Guatemala City, as do Schuster & Bonis (2008). Dávila et al. (2019) list material of Cuvieronius and Eremotherium from this locality.
Hacienda El Peñol
Colón (2015) notes that large fossil bones found here (in the Municipio de Atescatempa) in 1690 were thought to be the bones of Biblical giants. This appears to be the first known discovery of vertebrate fossils from Guatemala.
Hacienda San Carlos
Navarro (1856) mentioned mastodon bones from this locality, which is in the Municipio de Villa Nueva (Colón, 2015).
Jutiapa
Ibarra (1980) lists “pieza de mamut” from “unos pocos kilómetros de la cabecera departamental.” Colón (2015) calls this site “El Salitre” and states that it yielded a molar of Mammuthus columbi. He also refers to another nearby site called “El Recuerdo,” which yielded a “perezoso gigante.”
Kilómetro 31, Carretera CA-1 Occidente
Colón (2015) states that highway workers found a jaw fragment of a “mastodon” here in 1962.
Los Vados
This site is in the Municipio de San Pedro Ayampuc. Colón (2015) states that a fossil of a proboscidean or of Eremotherium was collected here.
Malacatancito
García & Ericastilla (1995, p. 115) mention extinct megafauna (“fauna mayor extinta”) from this locality. However, Mead et al. (2012) list only a gomphothere from this locality.
Paso Calandrias
Dávila et al. (2019) list an osteoderm of Glyptotherium from this locality in El Progreso.
Quebrada Pacoy
Located in the Municipio de Rabinal, Colón (2015) states that proboscidean fossils were found at this locality.
Quetzaltenango, Tívoli
García & Ericastilla (1995) mention fossils of glyptodont, elephant and mastodont from this locality. Mead et al. (2012, table 2) call this site Tivoli and list gomphothere, glyptodont, horse and mammoth? Colón (2015) states that in 1975 a glyptodont fossil was collected here. Dávila et al. (2019) list Glyptotherium armor from this site, which they call Xela. They also note the presence here of two glyptodont carapaces “in a layer of volcanic ash” (Dávila et al., 2019, p. 281).
Río Canbalam
Colón (2015) lists large fossil bones from this location, which is in the Municipio de Barillas.
Río Carboneras
Mota-Vidaurre (1989; also see Madden & Mota-Vidaurre, 1989) reported on proboscidean cheek teeth from the Herrería Formation along the Río Carboneras in the Departamento de Izabal (also see Lucas & Alvarado, 1995 for detailed provenance) (Figs. 15-16). Lucas & Alvarado (1995) assigned these teeth (Fig. 16A-C) to “Rhynchotherium blicki”, and they suggested a Hemphillian age by correlation to the Gracias Formation in Honduras. However, these teeth are not diagnostic of Rhynchotherium and are a likely Hemphillian record of Gomphotherium hondurensis (Lucas & Morgan, 2008; Lucas & Alvarado, 2010).
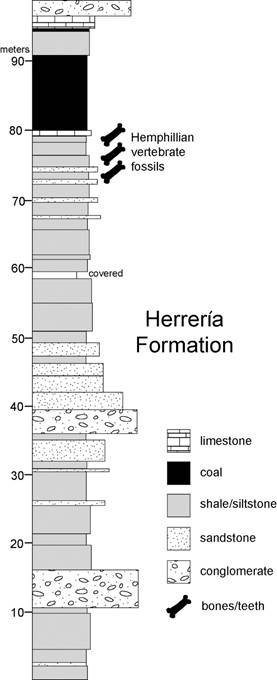
Fig. 15 Stratigraphic column of lower part of Herrería Formation along the Río Carboneras showing stratigraphic interval that produced Hemphillian mammals (redrawn and simplified from Romero, 1992).

Fig. 16 Hemphillian (late Miocene) mammals from the Río Carboneras, Guatemala. A-C, Gomphotherium hondurensis, plaster casts in the NMMNH collection of a right P4 (A), right M2 (B) and right M1 (C) in occlusal view. D-E, Neohipparion sp., m2 in labial (D) and occlusal (E) views. F-G, Aceratheriine rhinoceros, P2 in labial (F) and occlusal (G) views.
Additional fossils from the same locality along the Río Carboneras collected by J. Romero are an upper premolar of a rhinocerotid and a lower molar of a horse now in the collection of the Estanzuela Museum (Fig. 16D-G). The rhinoceros is represented by a right P2 (Fig. 16F-G) with a length = 24.3, width = 24.5 and height of the worn ectoloph = 14.4 mm. It has a thick and blunt parastyle, long and well worn ectoloph, a relatively thin and posteriorly projecting metastyle, a thicker and wider protoloph than metoloph and no antecrochet. It is clearly an upper premolar of an aceratheriine rhinoceros and is identical to a tooth illustrated by Webb and Perrigo (1984, fig. 6) from the late Miocene of Honduras. It is small for Aphelops (cf. Prothero 2005, table 4.7), and closer in size to a small species of Teleoceras, particularly the species T. guymonense (Prothero, 2005, tables 4.9-4.11; Lucas, 2008b). However, this isolated premolar is not diagnostic of a genus, so we identify it simply as Aceratheriinae.
The horse is represented by a right m2 (Fig. 16D-E) with a length = 22 and width = 13.2. It is high crowned, with a prominent parastylid and mesostylid, a shallow ectoflexid, a prominent isthmus and well developed pli caballinid and eurystylid. It compares well with molars of the hipparionine Neohipparion from the Hemphillian (MacFadden, 1984; Lucas, 2008a, fig. 6), so we identify it as Neohipparion sp. One reviewer of this paper suggested that this tooth may be assigned to N. affine or to N. cf. N. affine, and therefore the vertebrate faunal horzion in the Herrería Formation may be of late Clarendonian age. However, we are reluctant to assign a species name to an isolated equid molar, though the possibility of a late Clarendonian age should be considered.
These Hemphillian mammal fossils are from an interval of the lower part of the Herrería Formation about 5 m thick (Fig. 15). They co-occur with fossils of fishes, turtles and crocodiles (Romero, 1992), and this important late Miocene locality merits further collecting.
Río La Pasión
Woodburne (1969) documented a right maxilla of the peccary Dicotyles tajacu as part of an extensive late Pleistocene mammal assemblage from playas of the Río La Pasión and adjacent Río Santa Amelia (Sayaxché region, Departamento de Petén) near the town of Santa Amelia (2 km below the mouth of the Río Santa Amelia). Barnum Brown collected these fossils for the AMNH in the late 1940s and early 1950s. Woodburne (1969) listed (but did not document) the following taxa as part of this late Pleistocene assemblage: Dasypus, Pampatherium, Glyptodon, Megalonyx, Megatherium, Mylodon, Paramylodon, Hydrochoerus, Felis?, Cuvieronius?, toxodont, equid, Tapirus, Camelus, Mazama, and a bovid?
As noted by McDonald & Dávila A (2017), Brown’s collections are from multiple sites near Santa Amelia. They stated that Río de la Pasion or Santa Amelia refers to “numerous individual localities along the river and should not be treated as a single associated collection” (McDonald & Dávila A, 2017, p. 125). Indeed, Brown (1956) makes it clear that there were diverse fossil sites collected along the Río La Pasión, including some that were submerged in the riverbed. However, like Dávila et al. (2019) we treat the fossils from the Río La Pasión/Santa Amelia area as one assemblage (Table 1), pending data to separate the fossils into their different localities, which, however, are all likely of approximately the same age.
McDonald & Dávila A (2017, fig. 5) documented a lower molar of Mammuthus columbi from a site near Brown’s localities. Dávila et al. (2019, fig. 3, appendix) updated the list of taxa published by Woodburne (1969) to recognize Cuvieronius, Eremotherium, Megalonyx, Paramylodon, Glyptotherium, Pachyarmatherium, Holmesina, Neochoerus, Canis, Mixotoxodon, Tapirus, Dicotyles, Hemiauchenia and Palaeolama from this locality (Table 1), which they also called Santa Amelia.
Río Usumancita
Vinson (1962, p. 455) mentions the presence of Cuvieronius (“lower milk molar”), as well as bones of “Megatherium” (likely Eremotherium) in Pleistocene deposits in the vicinity of the Río Usumancita on the Petén-Chiapas frontier (also see Millan, 1985, p. 115).
Roca Madariaga
Colón (2015, p. 73) notes that a bone of a “mamífero gigante” was found at this site, which is in the Municipio de Oratorio.
Salamá
Gazin (1965) states that 5 km south of Salamá in northern Guatemala he observed fossils of Eremotherium. Ibarra (1980) lists Eremotherium from this locality, likely based on Gazin’s report. Colón (2015) refers to this site as “Escuela Forestal Centroamericana.” Mead et al. (2012, table 2) list only a megathere from this locality. Davila et al. (2019) list an Eremotherium maxilla with eight molariform teeth from this site.
San Rafaelito
This site is in the Municipio de San Rafael. A “mastodonte” molar and rib were collected here according to Colón (2015). Dávila et al. (2019) call this site “San Rafael las Flores” and list/document the following taxa: Cuvieronius, Eremotherium, Glyptotherium, Mixotoxodon and Odocoileus.
Sanguayabá
Located in the Municipio de Palencia, this locality yielded a “mastodon” bone (Colón, 2015).
Santa Cruz Barillas
Mead et al. (2012, Table 2) list this as a proboscidean locality, and Dávila et al. (2019) indicate a skeleton of Cuvieronius was collected here. Dávila et al. (2019) also list various specimens of Cuvieronius and Glyptotherium from this locality.
Santa Rosa
García & Ericastilla (1995) report fossils of Equus from this locality. Dávila et al. (2019) report Cuvieronius and Equus from this locality.
Songotongo
García & Ericastilla (1995) report but do not identify bones from this location near Chiquimula. Colón (2015, p. 71) states that “huesos largos y fragmentos de costillas” were found here. Dávila et al. (2019) identify a rib fragment from this locality as Eremotherium.
Treasure Island, Petén
Mook (1959, p. 2) documented a left maxillary of a crocodilian (Fig. 17) collected by Barnum Brown from a “probably middle Pleistocene” horizon on “Treasure Island.” This is not a place name in Guatemala but instead likely an informal name for a fossil locality in the Río La Pasión area where Brown collected extensively. Mook named the crocodile fossil a new subspecies, Crocodilus moreletii barnumbrowni, diagnosing the subspecies by its more angular external border of the maxilla and a maxillary plate between the palatine fenestra and the tooth row of variable width. Nevertheless, we are skeptical of the validity of this subspecies pending further documentation of the variablility of the maxillary in Crocodilus moreletii.

Tulumaje y Tulumajillo
From this locality, Gazin (1965) lists an “enorme perezoso” (likely Eremotherium). Ibarra (1980) lists Eremotherium and a proboscidean from this location, which is in the Valle del Río Motagua near San Agustín Acasaguastlán, Departamento de El Progreso. García & Ericastilla (1995) list a long bone of a proboscidean and a tibia of Eremotherium from this location (also see Schuster & Bonis, 2008). Mead et al. (2012, table 2) regard this as two localities-a megathere from Tulmaje and a proboscidean from Tulumajillo.
Late Miocene Fossil Vertebrates
The oldest fossil vertebrates from Guatemala are a handful of large mammals known from the Herrería Formation along the Río Carboneras. These are Gomphotherium hondurensis, an aceratheriine rhinoceros and the horse Neohipparion. The Gomphotherium molar described by Osborn (1926) from Chinautla is likely of the same age as this assemblage, but it is an isolated record, and its detailed provenance is not known.
These mammals compare well with the well known early Hemphillian (~ 7-8 Ma) vertebrate assemblage from the Gracias Formation in Honduras described by Olson and McGrew (1941), Corinto in northeastern El Salvador (Webb and Perrigo, 1984; Cisneros, 2008) and San Gerardo de Limoncito in Costa Rica (Laurito & Valerio, 2010), though the Costa Rican locality may be younger than these other sites (Rincón et al., 2020). Indeed, Rincón et al. (2020) published a radioisotopic age (87Sr/86Sr) of 5.80 Ma + 0.60/-0.80 for the Costa Rican locality. Furthermore, Gomphotherium has also been reported from the Miocene of panama (MacFadden et al., 2015) and from Chiapas, Mexico, near the Guatemalan border (Gómez-Pérez & Carbot-Chanona, 2018).
The Miocene mammals from Guatemala are few in number and diversity but are all North American forms, as are those from the correlative assemblages in El Salvador and Honduras. They thus support the conclusion that an extensive interchange of North and South American mammals across Central America had not taken place by about 8 million years ago, despite some recent claims to the contrary.
Late Pleistocene Fossil Vertebrates
Most of the vertebrate fossils from Guatemala are of Pleistocene age, as is the case in the other Central American countries (e. g., Lucas et al., 1997, 2007, 2008; Laurito et al., 2005; Cisneros, 2005, 2008; Aguilar & Laurito, 2009; Pérez, 2013; Lucas, 2014a; Lucas & Alvarado, 2017). The collective Guatemalan Pleistocene mammal fauna consists of the pampathere Holmesina, the dasypodid Pachyarmatherium, the glyptodont Glyptotherium, the sloths Megalonyx, Eremotherium and Paramylodon, the capybara Neochoerus, the cat Felis, the dog Canis, the toxodont Mixotoxodon, the gomphothere Cuvieronius, the mammoth Mammuthus, the tapir Tapirus, the horse Equus, the peccary Dicotyles, the camel Palaeolama, the deer Mazama and Odocoileus and the large bovid Euceratherium.
Most of the late Pleistocene sites in Guatemala yield Equus, Eremotherium or Mixotoxodon, as well as Cuvieronius. Similar assemblages are well documented throughout Central America and include Yeroconte in Honduras (Lucas, 2008c), El Hormiguero in El Salvador (Webb & Perrigo, 1984; Cisneros, 2005), Bajo Barrantes in Costa Rica (Lucas et al., 1997) and El Hatillo in Panama (Lucas, 2014b). These assemblages are dominated by the fossils of large edentates and ungulates and are a nearly equal mixture of South American (glyptodonts, sloths, toxodonts) and North American (horses, peccaries, deer, proboscideans) immigrants. Lucas (2014b) called these EME assemblages, referring to the association of Eremotherium, Mixotoxodon and Equus. But, given the common occurrence of the gomphothere Cuvieronius, we modify this to CEME assemblage. It is fair to say that the late Pleistocene mammal assemblages of Guatemala are very much like other age equivalent assemblages throughout Central America and present no unique or endemic records.
Discussion
We list two late Miocene and 36 fossil vertebrate localities in Guatemala, though, as noted above, some are composites of multiple localities. However, most of the late Pleistocene localities are just reports for which no fossils apparently reside in collections or have been described/illustrated. Of the Rancholabrean localities, six yield at least four genera that are documented (Table 1), so they are the foundation of our knowledge of Guatemala’s late Pleistocene mammal fauna (Dávila et al., 2019).
Taphonomic megabias
The Central American fossil record of Pleistocene mammals, like all records of fossil vertebrates, is biased by factors of taphonomy (e.g., Noto, 2011). However, we see an extensive and consistent taphonomic bias in this record, which can properly be called a taphonomic megabias, which is “a large-scale bias caused by differential preservation of organic remains representing particular organisms or ecosystems” (Behrensmeyer, 2021, p. 13).
This taphonomic megabias of the Central American vertebrate-fossil record is seen in a record dominated by relatively large and durable fossils of large mammals-large edentates and ungulates. Perhaps most notable are the cheek teeth of Cuvieronius, very dense and relatively large objects that are among the most common fossils of Central American vertebrates. The taphonomic megabias reflects preservation, discovery and collection of such relatively large mammal fossils, particularly of their most durable body parts, which are most easily preserved and discovered. The primary signal here is thus taphonomic, not biologic.
The Guatemalan vertebrate fossil record is very much a product of this megabias. Thus, it is almost devoid of lower vertebrates--only an undocumented report of a turtle from El Trifinio, the crocodile fossil published by Mook (1959) and rare fishes from the Río Carboneras (Romero, 1992). This important late Miocene locality merits further collecting. The Guatemalan vertebrate fossil record also lacks micromammals, as does most of the Central American vertebrate fossil record. Given that most of the diversity (and biomass) in late Cenozoic terrestrial vertebrate communities is in micromammals, this is a huge limitation of the Central American vertebrate fossil record,
This megabias thus skews the information in a systematic way so that we know little beyond the presence of large mammals in Central America. It greatly limits the biological interpretation of the Central American (and Guatemalan) record of fossil vertebrates.
Stratigraphy, Chronology and Correlation
As noted above, most of the records of fossil vertebrates reported from Guatemala unfortunately lack geological context and stratigraphic data, or data on their preservation (taphonomy). In a country with several active volcanoes with well know large explosive events (marker layers include pyroclastic flows and tephra falls) with known geochronological age control and a preliminary Quaternary geology (alluvial terraces and lacustrine deposits) established, it should be possible to reconstruct and correlate some paleontological localities based on the Quaternary stratigraphy of each site. Such data have been acquired in Costa Rica and Nicaragua (Lucas et al., 1997, 2008), for example, through site visits, talking to the people still alive that discovered the fossils or following the oral local tradition, or consulting newspapers, to arrive at a post-discovery reconstruction of the geological context, even though it may be several decades since the discovery. We could not recover most of the geological, stratigraphic or preservational context of Guatemala’s fossil vertebrates for the present paper except based on observations and analyses available in published sources (e.g., Romero, 1992; Mead et al., 2012; Dávila et al., 2019). Thus, a strong recommendation for the present and future generations of young Guatemalan geologists is to try to visit, recover and rebuild the geological context of the known vertebrate fossil sites in Guatemala.
Assignments of geologic age to Guatemala’s fossil vertebrates are primarily based on mammalian biochronology within the framework of the NALMAs (see above and Fig. 2). This biochronology assigns Hemphillian and Rancholabrean ages to the Guatemalan fossil vertebrates. However, more precise age assignments, particularly of the Rancholabrean vertebrates, are difficult to impossible. Indeed, the possibility that some of the Pleistocene mammal fossils from Guatemala are of Early or Middle Pleistocene age (older than Rancholabrean) needs to be considered, though current evidence does not identify older ages in the Pleistocene for the Guatemalan fossils.
Dávila et al. (2019, Table 1) reported 18 radiocarbon ages on bone bioapatite and two on charcoal from the localities Chinautla, Ciudad Real, Estanzuela, San Rafaelito and Xela. They drew attention to Holocene ages for a Glyptotherium from Xela and a Cuvieronius from Estanzuela and even suggested “that a local population of Cuvieronius may have survived into the Early Holocene” (p. 289). Dávila et al. (2019) also concluded that because nine of the radiocarbon dates are between 19,910 and 29,350 14CyBP, they indicate that most or all of Guatemala’s late Pleistocene vertebrates date to the last glacial maximum.
However, as Dávila et al. (2019) stressed, the radiocarbon ages they report are from bioapatite, and the possibility of C exchange with the environment may have compromised their accuracy, particularly in the hot and humid environments of Guatemala (e.g., Cherkinsky, 2009; Zazzo & Saliège, 2011; Wright, 2017). Furthermore, we note the wide ranges of their radiocarbon ages from particular sites. Thus, for example, seven ages from Estanzuela range from 9,539 to 22,400 14CyBP, and one charcoal age from Estanzuela is 39,670 14CyBP. Only the site San Rafaelito yields relatively consistent ages between 25,590 and 27,100 14CyBP.
Given the method and these inconsistencies, we are skeptical of the accuracy of the radiocarbon ages reported by Dávila et al. (2019). Most of these are late Pleistocene ages, and that is consistent with the Rancholabrean age the mammals indicate and the presence of the fossils in relatively superficial deposits. However, we attribute no more significance to these radiocarbon ages, especially the few Holocene ages. As Dávila et al. (2019) note, a more extensive and more rigorous analysis of the radiocarbon ages of Guatemala’s late Pleistocene vertebrates is needed.
Proboscideans
Lucas & Alvarado (2010) identified four immigrations of proboscideans from temperate North America to Central America: (1) late Miocene (early Hemphillian) immigration of Gomphotherium; (2) early Pleistocene (Blancan) immigration of Cuvieronius; (3) middle Pleistocene (Irvingtonian) immigration of Mammuthus; and (4) late Pleistocene (Rancholabrean) immigration of Mammut. The Guatemalan record of fossil proboscideans is consistent with three of these immigrations, of Gomphotherium, Cuvieronius and Mammuthus.
Mammoth fossils in Central America are found from Guatemala to Costa Rica but are relatively few and scattered-they are mostly single specimens (molars) without associated fauna (Arroyo-Cabrales et al., 2007; Lucas & Alvarado 2010; McDonald & Dávila A, 2017). Guatemala and Nicaragua have the largest numbers of mammoth localities, four each, and, as McDonald & Dávila A (2017) noted, most Central American mammoth localities are near the Pacific Coast. Those farthest away from the Pacific Coast are in Guatemala, particularly the record at Río La Pasión (Fig. 3).
Mammoths are well known to have been grazers whose presence implies grasslands/savannah habitats (e.g., Agenbroad, 1984). Such habitats must have existed in Central America to support mammoths, and they arrived in Central America during the Irvingtonian LMA, not long after their arrival in North America (Cisneros, 2005; Lucas & Alvarado, 2010; Lucas et al., 2017).
As McDonald and Dávila A (2017, p. 126) concluded, the distribution of mammoth fossils in Central America “likely reflects the lack of extensive collecting in these regions rather than any indication of the relative abundance of mammoths in Central America.” However, they did note that mammoths in Central America are much less common than gomphotheres and that this “is suggestive of a greater extent of forest compared to open country” (McDonald & Davila A, 2017, p. 127). These observations underscore our conclusion that the distribution of Mammuthus in Central America (and in Guatemala) is too limited to support anything other than very broad conclusions-Mammuthus immigrated to Central America, and there were at least local grasslands.
Dávila et al. (2019, fig. 8B) suggested the possibility that mammoths moved southward from Mexico into Petén in northern Guatemala and then followed northwest-southeast-oriented lowland corridors across the Motagua fault zone to reach southeastern Guatemala and then move southward into the rest of Central America. While this is a possibility, it is very speculative given the limited record of Mammuthus in Central America. One mammoth fossil from the Pacific side of Guatemala would refute the proposal of this immigration route. Indeed, if we examine the geographical distribution of fossil megamammal finds in Central America, most of them are concentrated in the most densely inhabited regions, which usually correspond to the Pacific coast and central area of each country.
Human association
Although claims of the association of Paleoindian artifacts and extinct Pleistocene mammals and of human alteration of the bones of extinct mammals have been made in Guatemala (e.g., Coe, 1960; Gruhn et al., 1977; Brown, 1980; Hayden, 1980), none have stood up to critical scrutiny. The evidence for Paleoindians in Guatemala is based on fluted, lanceolate projectile points. But, there is no direct/demonstrable association of artifacts with the bones of late Pleistocene mammals. For example, the claim of an association at Chivacabé (Hayden, 1980) is not borne out, as the points are found in loose colluvium, not in association with excavated, in situ bones (Yelacic, 2010; Lohse et al., 2012; Mead et al., 2012).
One petroglyph relevant to a supposed human-extinct megamammal association in Guatemala has not been published but instead has been presented online by Guatemalan archaeologist Sergio Ericastilla de Godoy (see remarq.ning.com/profiles/blogs/el-proboscidio-de-chuacs). This is the “proboscídio de Chuacus” in Baja Verapaz, a supposed petroglyph of a proboscidean. However, archaeologists Luis Hurtado de Mendoza, Phillippe Costa and Priscila Molina (personal communication, 2020) tell us that Paleoindian art is not known in Central America, and what has been attributed are hand prints of questionable antiquity. Therefore, a petroglyph of a proboscidean would be unique in Central America and not consistent with the record as currently understood. Although the “proboscídio de Chuacus” may be suggestive of a proboscidean in one view, if you turn it around, it resembles a flower or two spirals. The interpretation of rock art must be undertaken not only with engravings, but with knowledge of the sites, their correlation and their possible relative ages. Thus, we regard the “proboscídio de Chuacus” as unconvincing evidence that a Paleoindian registered an image of an extinct proboscidean.
Biogeography
The extant vertebrate fauna of Guatemala is a mixture of endemics that evolved in Guatemala and of descendants of immigrants from both North and South America (e.g., Iturralde-Vinent, 2006; Schuster & Bonis, 2008). However, the fossil record of Guatemalan vertebrates is a record of immigrants, not of any demonstrably endemic forms. There must have been some endemic evolution of vertebrates in Central America during the Pleistocene (Woodburne et al., 2006; Woodburne, 2010), but a fossil record to document that remains elusive. Perhaps the best candidates for endemic evolution are two ground sloth species known only from El Salvador (Webb & Perrigo, 1985). However, there is no evidence of endemism among Guatemala’s late Pleistocene record of fossil mammals.
Dávila et al. (2019, p. 289) noted that the composition of Guatemalan Pleistocene mammal assemblages “is consistent with other associations documented along the Mesoamerican corridor…which also include predominantly Cuvieronius, Eremotherium, Glyptotherium and Mammuthus.” Despite this, they concluded that “the underrepresentation (in Guatemala) of typical South American fauna suggests a more southerly located biogeographic juncture between the two subcontinents (North and South America), at least for the Pleistocene” (Dávila et al., 2019, p. 277). However, we see no evidence in the fossil record of Guatemalan fossil vertebrates to support such a conclusion. Guatemalan late Miocene vertebrates are of North American affinity, predating the Great American Biotic Interchange. In contrast, the late Pleistocene vertebrates of Guatemala are a mixture of North American and South American immigrants like that seen in the fossil record of the rest of Central America.

