











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroduction
The genus Yaberinella is relatively rare in the geological record. Few species have been reported from the Eocene of Jamaica (Hottinger, 1969; Lehmann, 1961; Robinson, 1993; Robinson and Wright, 1993; Vaughan, 1928, 1929), Panama (Cole, 1952), and Nicaragua (Robinson, 1993), making it an endemic genus restricted to the Caribbean area. Other unpublished or “hard-to-find” documents report the presence of Yaberinella in Jamaica and Nicaragua (Eva, 1976; Robinson, 1988 - See Robinson, 1993). Additionally, Butterlin, in an unpublished work, mentioned the presence of Yaberinella jamaicensis Vaughan, the genus type species, in Costa Rica (Adams et al., 1986). This report comes from the species recognition of Yaberinella by Butterlin in a sample collected in the southwest of the Nicoya Peninsula (Mora, 1985). More recently, Bolz and Calvo (2020), who also cite this first Costa Rican report, confirmed the presence of this species in two additional samples from the Nicoya Peninsula. Also, Yaberinella jamaicensis has apparently been evidenced in western Oman (Adams and Racey, 1992). This finding, contradicting the Caribbean area endemism of the genus, has recently been qualified as inconceivable and considered as being the product of man-made confusion (BouDagher-Fadel, 2018). In the same book, BouDagher-Fadel cites a personal communication of Hottinger saying the following: “I have been at the point indicated near Muscat: No trace nor even a possibility of their occurrence there considering the local geology” (BouDagher-Fadel, 2018, p. 517). The latter affirmations being said without scientific proof, we may use them with precaution. The most recent revision of Jamaican and Nicaraguan material mentions four distinct species, reordering the older systematic, which reported only two (Robinson, 1993): Yaberinella hottingeri, Y. jamaicensis, Y. jamaicensis trelawniensis, Y. panamensis. These four species are specific to the middle/late Eocene deposits. Y. hottingeri seems to mark the Lutetian, excepting the higher part of the stage (Robinson, 2004). Y. jamaicensis trelawniensis would mark the late Lutetian to the earliest Bartonian (Robinson, 2004). Y. jamaicensis apparently appears during the end of the Lutetian and disappears at the end of the Bartonian in the central Caribbean region (Jamaica and Nicaragua Rise) (Robinson, 2004), while in southern Central America its composite range comprises both stages, the Bartonian and the Priabonian (Bolz and Calvo, 2020; Cole, 1952). Y. panamensis, however, extends only in the late Bartonian of Jamaica (Robinson, 2004), but reaches the Priabonian in the type locality of Panama (after Cole, 1952).
In the present contribution, we report the occurrence of the genus Yaberinella in early-late Eocene (Bartonian) carbonate deposits of southwestern Costa Rica (Fig. 1-A). A systematic intent to relate the observed specimens with two of the previously described species is made, and a short comment about their presence is proposed.

Fig. 1: A. Localization of the studied samples (yellow star) in southwestern Costa Rica (black box). Top and left grid is in Costa Rican coordinates (CRTM05). Bottom and right grid is in international coordinates (WGS84). B. Photograph of the outcrop where the Yaberinella-bearing sample has been picked-up (black arrow). Note the fractures preventing any dip measurements. The visible part of the outcrop measures about 10 metres from bottom to top (image taken by Sergio Vega). C. Outcrop situated about 37 meters east of the Yaberinella-bearing bed from which sample CPII4 comes (black arrow). The deposit is tectonically disrupted, exhibiting beds (dm) dipping to the northeast. Person on the left for scale.
Material and methods
The material examined comprises two different limestone samples labelled CPII3 and CPII4 collected from outcrops of the Fila de Cal Formation exposed in the Piedras Blancas River, southern Costa Rica (Fig. 1A). Both samples are bioclastic larger foraminiferal-calcareous algal microfacies. The study includes a systematic approach to Yaberinella specimens found in sample CPII3 and an examination of the larger foraminiferal assemblages contained in both samples, CPII3 and CPII4, to establish their biostratigraphic ages. Sample CPII3 exhibits abundant tests as well as test fragments of Yaberinella specimens, taxonomically examined in detail. Four thin sections from this sample, named CPII3-1 to CPII3-4, were prepared for recognition and measurements. The samples being limestones, difficulties were encountered to obtain well-oriented sections of Yaberinella. Descriptions are based on direct observation of thin sections under the stereomicroscope, as well as on photomicrographs and scanned images. Identification of foraminiferal tests has been done on axial and/or equatorial cuts. Biostratigraphic age determination is based on comparison with assemblages of the Gulf of Mexico, Central America and northern South America (e.g., Bolz and Calvo, 2018, 2019, 2020; Bolz et al., 2019; Butterlin, 1981; Cole, 1952, 1953; Eames et al., 1968; Pécheux, 1984). Measurements of whorls on axial and slightly oblique sections of thirteen specimens permit to see the relative evolution of coiling (Plate 1, figs. 1-4; Plate 2, figs. 1-5; Plate 3, figs. 1-4). Axial sections prohibiting the view of full whorls, these lasts are divided in half parts (Fig. 2). In contrast, the other larger benthic foraminiferal determinations have exclusively been made on high-resolution scanned images and photomicrographs. Regarding the palaeontological approach, the absence of oriented equatorial sections of Yaberinella tests could eventually have compromised the data. Both samples were collected in 2018 by a group of geology students during field practice of the undergraduate course Geología de Campo II given at the Central American School of Geology of the University of Costa Rica. The two rock samples, including their thin sections, are stored in the School’s paleontological collection at the University of Costa Rica. The sampling coordinates of CPII3 are: -83.2219523587; 8.79118703169 (WGS84); that of CPII4 are: -83.220259651; 8.79218715886 (WGS84).
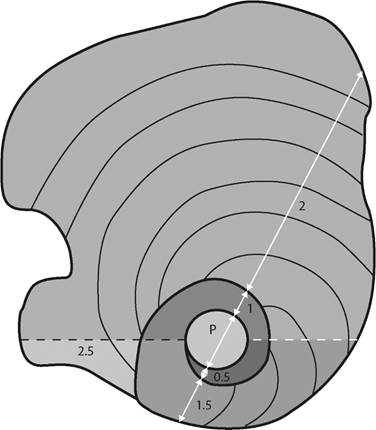
Fig. 2: Schematic equatorial section of a Yaberinella. The horizontal black dashed line indicates the boundary of whorls. The white horizontal dashed line indicates the boundary of half whorls. The white arrows show the length measured for every cut “half whorl” on a hypothetic axial section (see tables 1 and 2).
Sample CPII3 comes from a massive and fractured bed pertaining to the Fila de Cal Formation (Fig. 1-B). These fractures, which are due to a nearby important N-S fault, do not allow to truthfully calculate the thickness of the outcrop. However, the nearest obtained dip direction (N26-65° - WGS84: -83.221616017; 8.79120442047; Fig. 1-C-), 37 metres east of the former sampled outcrop, is like most measurements of the Fila Costeña in other publications. This suggests that the stratigraphic section should continue vertically for at least tens of metres in thickness. Thus, the upper outcrop from which sample CPII4 comes stratigraphically overlies the sampled Yaberinella-bearing bed. It consists of decimetric to metric limestone beds exhibiting planar to undulated contacts (Fig. 1-D), also pertaining to the Fila de Cal Formation. A waterfall and large fallen blocks did hinder to explore the canyon more than 80 metres to the northeast of the latter GPS point. In this area, the dip direction changes because of the same N-S fault (N155-35°).
Systematic Palaeontology
Phylum Foraminifera d’Orbigny, 1826
Class Tubothalamea Pawlowski, Holzman and Tyszka, 2013
Order Miliolida Delage and Hérouard, 1896
Family Soritidae Ehrenberg, 1839
Genus Yaberinella Vaughan, 1928
Type species. Yaberinella jamaicensisVaughan, 1928
Remarks: Different terms have been used to describe the internal structure of Yaberinella. Vaughan (1928) originally installed the term criss-cross pattern, talking about sets of crossing light lines and dark plates. The lines cross at an angle of 90 to 135°. Lehman (1961) described the endoskeleton in three dimensions, using the terms ramps (=plates of Vaughan), stolons (=lines of Vaughan), and attics (=lines of Vaughan). Adams and Racey (1992) agree with the general description but prefer using the terms partitions (=plates/ramps) and tubes (=lines/stolons and attics). For a better understanding and for the reasons cited by Adams and Racey (1992), we may use the latter terms for our descriptions.
Yaberinella panamensisRobinson, 1993
(Plate 1, figs. 1-4; Plate 2, figs. 1-5; Plate 3, figs. 1-4)
Yaberinella jamaicensis Vaughan, Cole, 1952, p. 8, pl. 6, figs. 1, 2, 4-6.
Yaberinella panamensisRobinson, Robinson, 1993, p. 55, pl. 8, fig. 2.
Description: The general form is elongated, discoid and biconvex. Some small specimens exhibit an ovoid shape, while other are more ellipsoidal. Because of cut orientation, larger specimens (15 and 27.6 mm) (Plate 4, fig. 1) general form is not described. On this large specimen, observed on an almost equatorial cut, sets of tubes cross at an angle of about 90° to 132°, presenting a typical criss-cross pattern (Plate 4, figs. 1-3) (Vaughan, 1928). On axial sections, tubes and partitions generally appear as wavy or in zigzag. Measurements of proloculus diameters of thirteen specimens yielded a mean value of 0.545 mm (range from 0.255 to 0.715 mm) (Table 1). Although most of the examined specimens show a single spherical proloculus, some individuals exhibit a bilocular or irregular embryonic chamber (Plate 1, fig. 3; Plate 2, fig. 5; Plate 3, fig. 2). Mean total diameter measured on twelve specimens is 3.76 mm (range from 1.29 to 7.81 mm) (Table 1). The mean maximum thickness (central bulge) calculated for ten specimens is 0.741 mm (range from 0.475 to 1.308 mm). The mean size of the first half of whorl one is 0.428 mm, while the mean size of the second half is 0.932 mm (Table 1), a bit more than two times bigger. The mean size of whorl one and a half is 1.562 mm, about three times bigger than the first half of whorl one. Very few specimens reach more than 1.5 whorl, giving mean values that cannot be used truthfully.
Table 1 : Measures on axial sections of thirteen specimens of Yaberinella panamensis.
| Thin section | - | - | - | CPII3-1 | - | - | - | CPII3-2 | - | - | CPII3-3 | - | CPII3-4 | - |
| Specimens | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | Mean values |
| Proloculus diameter (mm) | 0.490 | 0.405 | 0.605 | 0.700 | 0.555 | 0.465 | 0.450 | 0.685 | 0.595 | 0.715 | 0.695 | 0.475 | 0.255 | 0.545 |
| Whorl 0.5 (mm) | 0.380 | 0.310 | 0.250 | 0.180 | 0.330 | 0.340 | 0.320 | 0.330 | 0.790 | 0.500 | 1.230 | 0.540 | 0.070 | 0.428 |
| Whorl 1 (mm) | 0.830 | 1.100 | 0.630 | 0.600 | 0.850 | 0.590 | 0.520 | 0.850 | 1.370 | 0.600 | 2.650 | 1.390 | 0.140 | 0.932 |
| Whorl 1.5 (mm) | 3.870 | 1.430 | 1.680 | 0.780 | 0.280 | 0.260 | 0.420 | 4.560 | 0.780 | 1.562 | ||||
| Whorl 2 (mm) | 1.660 | 2.130 | 0.370 | 0.990 | 1.288 | |||||||||
| Whorl 2.5 (mm) | 2.430 | 2.430 | ||||||||||||
| Max. thickness (mm) | 0.808 | 0.657 | 0.812 | 0.809 | 0.648 | 0.680 | 0.578 | 1.308 | 0.638 | 0.475 | 0.741 | |||
| Total diameter (mm) | 7.23 | 7.81 | 3.17 | 2.63 | 2.02 | 1.66 | 1.29 | 3.28 | 2.76 | 6.38 | 4.58 | 2.41 | 3.76 | |
| Number of whorls | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 | 1 | 2.5 | 1 | 1.5 | 1.5 | 1.5 | 1.5 |
Remarks: The mean proloculus size is slightly smaller to that reported for Panamanian specimens (640-680 µm: Cole, 1952), and much smaller than that of Y. jamaicensis from other localities [0.9-1 mm: Vaughan (1928); 1 mm: Lehman (1961); 0.5-1 mm: Hottinger (1969); 07-1.11 mm: Adams and Racey (1992); 0.5-1 mm: Robinson (1993), Robinson and Wright (1993)]. The proloculus size of specimens found on the Nicoya Peninsula of Costa Rica is within the same range (Bolz and Calvo, 2020). Although for these authors the proloculus size is not a reliable criterion for distinguishing among the species of the genus. However, the large range of sizes observed is consistent with report made from the Panamanian specimens. The results on whorls calculations agree with the description made for the type species (Robinson, 1993) and with pictures of the first observation (Cole, 1952). It indicates that the operculine stage of coiling is missing, and that the flaring stage apparently and generally starts at about whorl one and a half. This flaring stage rarely expands enough to make the illusion of a finishing second whorl on axial sections. The mean extent of the first chamber has not been measured for a lack of good-oriented section. Nonetheless, it can be supposed that it occupies more than one whorl (Plate 1, fig. 1), as described for Y. jamaicensis (Vaughan, 1928).
Age-distribution: Bartonian of Costa Rica, Fila de Cal Formation. This age is in accordance with ranges given in Robinson (2004) for Jamaica and the Nicaragua Rise. However, it is older than the Priabonian of the Panamanian specimens originally described by Cole (1952). The outcrop is situated on the southwest Pacific side of the country, in the Piedras Blancas River, about 2.1 km northeast of the bridge in the town of Piedras Blancas (WGS84: -83.2219523587; 8.79118703169) (Fig. 1-A). Yaberinella panamensis occurs together with the following other foraminifers which permitted a better age determination: Lepidocyclina (Neolepidina) macdonaldi Cushman, L. (Neo.) pustulosa pustulosa H. Douvillé, L. (Nephrolepidina) chaperi Lemoine and R. Douvillé, Nummulites floridensis Heilprin, Fabiania cassis (Oppenheim) var. cubensis (Cushman and Bermúdez), Gypsina vesicularis (Parker and Jones), Pellatispirella matleyi (Vaughan), Yaberinella hottingeri Robinson, Helicostegina sp., Amphistegina sp. A sample labelled CPII4, collected a few tens of meters northeast of the Yaberinella-bearing bed, gave a slightly younger age (Priabonian). The following larger foraminifers were recognized in CPII4: Lepidocyclina tobleri Douvillé panamensis (Cushman), L. (Neolepidina) macdonaldi Cushman, Asterocyclina minima (Cushman), Nummulites floridensis Heilprin, Helicostegina sp.
Yaberinella hottingeriRobinson, 1993 (Plate 2, fig. 6)
Yaberinella trelawniensis Vaughan, Hottinger, 1969, p. 746, pl. 2, fig. 3; pl. 5, figs. 1-6.
Yaberinella hottingeri Robinson, Robinson, 1993, pl. 2, fig. 5; pl. 4, fig. 6 ; pl. 5, figs. 5-7.
Yaberinella hottingeri Robinson, Robinson and Wright, 1993, fig. 15.11.
Description: The described specimen appears in a axial section. Its form is ovoid-elongated and biconvex. The proloculus diameter is 0.12 mm. Test diameter is 2.84 mm. The thickness (central bulge) reaches 0.8 mm. Size detail of each whorl is given in table 2. The internal structure of endoskeleton is irregular, not showing the typical “criss-cross” pattern of other Yaberinella species.
Table 2: Measures on axial sections of one specimen of Yaberinella hottingeri.
| Thin section | CPII3-3 |
| Proloculus diameter (mm) | 0.120 |
| Whorl 0.5 (mm) | 0.200 |
| Whorl 1 (mm) | 0.270 |
| Whorl 1.5 (mm) | 0.200 |
| Whorl 2 (mm) | 0.270 |
| Whorl 2.5 (mm) | 0.230 |
| Whorl 3 (mm) | 0.400 |
| Whorl 3.5 (mm) | 0.530 |
| Whorl 4 (mm) | 0.620 |
| Max. thickness (mm) | 0.800 |
| Total diameter (mm) | 2.84 |
Remarks: The general form, absence of a final flaring stage, and number of whorls (4) are like those described by Robinson (1993) and Robinson and Wright (1993). Additionally, the poorly ordered internal chamberlets, not showing the typical “criss-cross” pattern of other species, is also like the initial description. However, this data could be biased because of a lack of equatorial sections observation. The proloculus diameter is, however, smaller than that of the type species. The smaller size of proloculus, together with the absence of the final flaring stage, are the principal differences permitting to separate this form from Y. jamaicensis.
Age-distribution: Bartonian of Costa Rica, Fila de Cal Formation. This age disagrees with ranges given in Robinson (2004). Nonetheless, the unique found specimen could eventually be reworked from older Lutetian rocks. The outcrop is situated on the southwest Pacific side of the country, in the Piedras Blancas River, about 2.1 km northeast of the bridge in the town of Piedras Blancas (WGS84: -83.2219523587; 8.79118703169) (Fig. 1-A). Yaberinella hottingeri occurs together with the following other foraminifers which permitted a better age determination: Lepidocyclina (Neolepidina) macdonaldi Cushman, L. (Neo.) pustulosa pustulosa H. Douvillé, L. (Nephrolepidina) chaperi Lemoine and R. Douvillé, Nummulites floridensis Heilprin, Fabiania cassis (Oppenheim) var. cubensis (Cushman and Bermúdez), Gypsina vesicularis (Parker and Jones), Pellatispirella matleyi (Vaughan), Yaberinella panamensis Robinson, Helicostegina sp., Amphistegina sp. A sample labelled CPII4, collected a few tens of meters northeast of the Yaberinella-bearing bed, gave a slightly younger age (Priabonian). The following larger foraminifers were recognized in CPII4: Lepidocyclina tobleri Douvillé panamensis (Cushman), L. (Neolepidina) macdonaldi Cushman, Asterocyclina minima (Cushman), Nummulites floridensis Heilprin, Helicostegina sp.
Discussion
Yaberinella is considered as a Caribbean endemic genus, occurring between a narrow latitudinal range (9° to 18°). The southernmost observation was made in the deposits of the Gatuncillo Formation of the Panama Canal Zone (Cole, 1952). The northernmost observations come from the Yellow Limestone of Jamaica and the Nicaragua rise (Hottinger, 1969; Lehmann, 1961; Robinson, 1993; Robinson and Wright, 1993; Vaughan, 1928, 1929). The new find made from Costa Rica slightly extends the latitudinal range up to a few minutes to the south. At the exception of the Nicoya Peninsula records (Bolz and Calvo, 2020; Mora, 1985) and the controverted evidence from Oman and its provincialism implications (Adams and Racey, 1992), older records of Yaberinella are mostly from the Caribbean side of Central America. The actual location of the Costa Rican record, on the southwest Pacific side of the country, confirms its occurrence outside the Caribbean area. Considering the mixing of water masses between the Pacific and the Atlantic oceans in Central America during the Eocene, it is not, and has never been worthy to separate their respective marine faunas. Naturally, genera and species of larger foraminifers described from both sides have always been consistent. However, the rarity of Yaberinella in the Pacific area of Costa Rica (four records in 35 years and hundreds of samples), implies a probable complex marine current mechanism or a strict water chemistry necessary in enhancing its dispersal. A combination of these two factors is most probable. The Bartonian record in southwestern Costa Rica of a likely older form attributed to Yaberinella hottingeri alongside Y. panamensis shows the extension of the species habitat to the Pacific side of southern Central America. This could argue against Adams and Racey’s (1992) view of the alleged migration of Yaberinella from the Caribbean to Oman through Africa. On the other hand, even if faunistic correlations exist between the Caribbean area and the Middle East (Schlagintweit and Consorti, 2020), the peculiarity of this form (Y. hottingeri) compared to other species of the genus supports the idea that Caribbean area conditions played an important and restricted role in the development and evolution of Yaberinella. Previous finds on the Nicoya Peninsula (e.g., Bolz and Calvo, 2020) and the newly record in the Fila de Cal range, both located in the Costa Rican Pacific area, actually mark the westernmost occurrence of the genus in Central America. Besides, the fact that Priabonian beds (sample CPII4) rest over Bartonian Yaberinella-bearing deposit (sample CPII3) agrees with the time slice given for Yaberinella panamensis in Jamaica and Nicaragua Rise (late Bartonian: Robinson, 2004). It does, however, not agree with the younger age of Yaberinella panamensis in the type locality, in Panama (Priabonian: after Cole, 1952).
Additionally, Yaberinella hottingeri has previously been recorded only from early Lutetian, and uniquely in Jamaica-East Nicaragua. Its observation in Bartonian of southwestern Costa Rica implicates a probable larger, or different time slice extent, at least in this area - although eventually reworking from older deposits cannot be ruled out in this bioclastic microfacies -.

