











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
En el presente trabajo se analizan trazas fósiles que se encuentran en las areniscas cuarzosas de la Formación Coris. Las trazas no fueron observadas en el afloramiento, se observaron ex situ, en rocas ornamentales, cortadas para ser utilizadas como enchape, procedentes de las facies de areniscas cuarzosas típicas de la unidad (Obando, Bottazzi y Alvarado (1991).
Estas trazas se desarrollaron en ambientes marinos someros marginales (plataforma), los cuales son frecuentes en los depósitos del Mioceno y Plioceno de Costa Rica. Las principales características de estos ambientes es que son áreas de alta energía por olas, tormentas, corrientes oceánicas y mareas, aunque en los estuarios predominan las condiciones de aguas tranquilas. Los depósitos en sedimentos marinos marginales son principalmente arenas de cuarzo bien seleccionadas, lodos y gravas que constituyen deltas, playas, islas de barrera, estuarios, lagunas y llanuras de marea (Boggs, 2009, Nichols, 2009). Esta variabilidad de ambientes depende de factores locales como el rango mareal, la energía de las olas y las tormentas y factores regionales como la subsidencia de la cuenca, variaciones en la tasa de sedimentación y los cambios eustáticos del nivel del mar (Mcbride y Moslow, 1991).
Los sedimentos estudiados corresponden con ambientes de playa, los cuales pueden ser divididos en tres zonas (Boggs, 2009):
- Backshore (trasplaya), la cual se extiende hacia el continente desde la berma de playa sobre el nivel de marea alta.
- Foreshore (playa) llamada zona litoral o zona intermareal.
- Shoreface (nearshore) (frente de playa) que se extiende desde el nivel base de las olas de buen tiempo hasta una profundidad de 10-15 m, pero puede profundizarse durante las tormentas.
El transporte de sedimentos en la playa y el shoreface está dominado por las olas y olas inducidas por corrientes, aunque las corrientes de marea pueden ser localmente importantes cerca de los canales de marea y los estuarios (Boggs, 2009).
Las trazas de D. ottoi estudiadas, se desarrollaron en el shoreface, específicamente en el upper shoreface (Obando et al., 1991), donde los depósitos son muy variables debido a las complejas condiciones hidráulicas de la zona de rompiente, lo que genera una secuencia compleja de estructuras sedimentarias multidireccionales entre ellas estratificación cruzada de bajo ángulo tipo hummocky y swaley cross stratification (Mial, 2000; Nichols, 2009) y sedimentos de textura variable (Boggs, 2009).
Estas condiciones favorecen el desarrollo de trazas epi y endobentónicas, perforaciones inclinadas en forma de U muchas veces con spreites protrusivas, tubos simples o ramificados, algunos tubos verticales. Los organismos productores pueden incluir carnívoros móviles y diversos suspensivoros y comedores de detrito. Los sedimentos se encuentran de moderada a intensamente bioturbados, por lo que muchas veces se pierden las estructuras por homogenización. Estas condiciones caracterizan la icnofacies Cruziana (Bromley, 1996). La bioturbación es más intensa y frecuente en sedimentos arenosos, debido a que las corrientes que transportan y depositan arena pueden traer nutrientes para los organismos bentónicos que las habitan (Nichols, 2009).
Se analizaron once individuos de D. ottoi y gran cantidad de cortes transversales. Las rosetas individuales varían en tamaño y preservación y debido a que presentan algún grado de inclinación con respecto a la estratificación original, los cortes son diversos.
Este reporte se hace debido a que los icnofósiles pertenecientes al género Dactyloidites son poco comunes en el registro sedimentario (Lazo et. al., 2008), los ejemplares estudiados están muy bien conservados y por no haber sido reportados antes en dicha unidad ni en otras unidades del país. Además, por ser depósitos de aguas someras donde la escasez de fósiles es notable y las posibilidades de conservación son bajas, por las condiciones hidrodinámicas tan variables y las severas y estresantes condiciones ecológicas (Nichols, 2009).
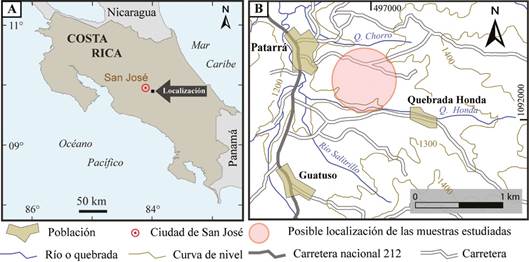
Las muestras probablemente provienen de una cantera ubicada 1,5 km al sureste de la población de Patarrá (Fig. 1), uno de los sitios más característicos donde afloran las ortocuarcitas de la Formación Coris.
Contexto geológico
Los icnogéneros estudiados se encuentran en rocas sedimentarias provenientes del Sur del Valle Central. Corresponden con facies silicoclásticas, asociadas a ambientes someros de la Formación Coris, definida por Castillo (1969) y constituida principalmente por ortocuarcitas o areniscas cuarzosas, areniscas volcaniclásticas, conglomerados, tobas, lutitas carbonosas y lignito (Denyer y Arias, 1991). Fischer y Franco (1979); realizan un estudio paleontológico detallado de la formación, describiendo los fósiles e icnofósiles e indicando que la unidad se depositó en un ambiente litoral a terrestre, reconociendo tres facies (facies marina del sublitoral, facies intermedia de aguas someras y facies terrestre), que ocurren durante el Mioceno Medio tardío- Mioceno Superior tardío. Estos autores mencionan la presencia de estructuras biogénicas, entre ellas tubos en U con spreites y galerías o estructuras de minería organizada. Los restos analizados provienen de la facies intermedia de aguas someras, que describen como carente de fósiles excepto trazas de actividad de organismos en algunos niveles. Además, mencionan la presencia de laminación paralela, angular o cruzada, por ondulitas, placeres de minerales pesados, capas de gradación simple y normal y paraconglomerados intercalados. Hay capas de areniscas cuarzosas puras con buena selección, redondez y madurez textural, que presentan una estratificación masiva. Obando et al., (1991) mencionan que la secuencia está compuesta de ciclos progradantes que se relacionan con depósitos de strand plain (llanuras costeras), reconociendo cuatro facies (facies de plataforma silicoclástica, facies de plataforma abierta, facies de frente de barra y facies de barras arenosas costeras). Las trazas estudiadas se encuentran en las facies de barras arenosas costeras (upper shoreface), en lo que corresponde a la subfacies cordón de playa, caracterizada por niveles cuarzo arenosos bien seleccionados y limpios, de grano medio a grueso, con laminaciones paralelas e inclinadas de bajo ángulo tipo hummocky cross stratification (HCS), que indican un ambiente de depositación de alta energía (Obando et al., 1991), afectados por tormentas.
Resultados
Ichnogenero DACTYLOIDITES Hall, 1886
Dactyloidites ottoi (GEINITZ, 1849)
Descripción
Las trazas se encuentran bien preservadas, pero se observan en cortes donde solo se ven los elementos radiales y no hay indicios del conducto central. La ausencia de un conducto central se puede explicar como resultado de los procesos tafonómicos, ya que este conducto tiene un potencial de fosilización menor que los elementos radiales (Uchman y Pervesler, 2007).
La traza está conformada por galerías superpuestas con varios elementos alargados y subhorizontales, dispuestos radialmente en forma de abanico o de roseta y que salen de un conducto central (no reconocible en nuestros ejemplares, debido a la preservación y a que presentan algún grado de inclinación con respecto a la superficie del fondo). Estos abanicos tienen un rango entre 120 y 180 grados y se pueden reconocer de 3 a 6 elementos principales, los cuales presentan una o varias ramificaciones, llegando a tener entre 10 y 13 ramas en la parte terminal (Fig. 2 A, B). Los elementos principales generalmente presentan una dicotomía o bifurcación primaria o isótoma cerca del tubo o conducto central. Los elementos son cercanos entre sí, el ángulo entre elementos contiguos varía entre 20-30 grados (Fig. 2 A, B). A veces también presentan una bifurcación secundaria o heterótoma, que se produce hacia un solo lado de la estructura, generando ramas más cortas, dando como resultado un patrón de ramificación asimétrico (Fig. 2 A, B). Los elementos principales tienen un largo más o menos uniforme en cada roseta, variando entre 3 cm y 6 cm en los diferentes individuos y un ancho variable entre 0,5 a 0,7 cm en el mismo individuo (Fig. 2 A). En la sección transversal el corte no es circular, más bien en la parte superior es convexo y en la parte inferior tiende a ser plano o irregular, como lo señalan Agirrezabala, y Gilbert, 2004 (Fig. 3 A, B). Los elementos radiales presentan un relleno de coloración desigual (generalmente más claro (Fig. 2 B,) o de color diferente (Fig. 4 A) que la roca circundante, pero en los cortes no se notan diferencias texturales entre el relleno y la roca que los rodea, por lo que se puede inferir que el productor mientras se alimentaba de la materia orgánica, generaba una selección, remoción y transporte de las partículas apenas visible. Los cortes no permiten ver claramente los spreites, solo se pueden apreciar algunas diferencias en el relleno y las terminaciones de las estructuras que son redondeadas o romas, producto de los spreites protrusivos (Fig. 2 B, 3 C, 4 A).
Los abanicos aparecen separados en la roca, no se nota solapamiento de estructuras, posiblemente por ser estructuras de corta duración. Se notan muy bien diversas galerías superpuestas o escalonadas en los cortes transversales (Fig. 3 A, B).
Paleoecologia/Icnocoenosis
Las trazas de D. ottoi, se encuentran en areniscas cuarzosas de grano medio, color amarillo a pardo rojizo, con laminaciones paralelas planas y curvas, perturbadas por el retrabajo intenso de organismos bentónicos. Estos sedimentos se acumularon en un ambiente marino somero afectado por alternancia de eventos erosivos de alta energía, tormentas (storm event suite) y depositación de baja energía (fair weather suite) (Mayoral, Ledesma-Vazquez, Baarli, Santos, Ramalho, Cachão, da Silva y Johnson, 2013), en el upper shoreface donde las condiciones ambientales son muy variables, con muchas fluctuaciones (Boggs, 2009; Nichols, 2009). Las trazas posiblemente fueron producidas en los periodos más estables, con predominio de baja energía que sucedieron a los eventos de alta energía (Fursich, Uchman, Alberti y Pandey, 2018). En los cortes no se ha observado una relación directa de las trazas con las estructuras típicas de eventos de alta energía como la estratificación cruzada HCS, aunque si se aprecian cortes de D. ottoi asociados a laminación paralela planar y curva (Fig. 4 B, C,D). Además, se observan relictos de laminación cruzada en niveles muy retrabajados que no permiten definir con precisión la morfología de las trazas (Fig. 4 B,D).
D. ottoi aparece en conjunto con otras trazas de la Formación Coris, principalmente tubos de Ophiomorpha nodosa, que se preservan como cortes transversales y longitudinales con diámetros que varían entre 1 cm y 2,5 cm y longitud hasta de 8 cm (Fig. 3 D, E), producto igualmente de organismos oportunistas que colonizaron el sustrato arenoso blando, bien oxigenado, en periodos de estabilidad y con disponibilidad de materia orgánica que favorecieron la colonización ( Lazo, Palma y Piethe, 2008, Mayoral et al., 2013).
En los ejemplares estudiados el conducto o tubo central no se preserva, debido a que generalmente está vacío o se rellena de forma pasiva, por lo que colapsa muy fácilmente cuando es abandonado, a diferencia de los elementos radiales que contienen sustancias orgánicas producto de la descomposición de los restos fecales, que los hace más resistentes (Gilbert et al. ,1995). Otra explicación para la no conservación del conducto central puede ser las diferencias de inclinación de las estructuras radiales o rosetas con respecto al fondo o al plano de estratificación (López y Ardévol, 1999), que implican diferencias en la preservación, por lo que los más inclinados solo conservan sus terminaciones (Fig. 3 A), mientras que los menos inclinados tienden a preservar una mayor proporción de la longitud de las ramas (Fig. 2 B, 3 C). En estos casos la traza resultante tiene forma de abanico y el conducto vertical no se preserva (Lazo et al., 2008).
Fürsich y Bromley (1985) indican que el hecho de que las rosetas individuales nunca sean completamente circulares se debe a una buena utilización de los recursos por parte de los organismos productores, por lo que no necesitan completar la circunferencia.

Fig. 2: A) Esquema que presenta la conformación de una roseta o abanico con las ramificaciones primarias y secundarias (inspirado en Lazo et a., 2008). B) Abanicos o rosetas de D. ottoi, donde se observan los elementos radiales elongados y las ramificaciones primarias (isótomas) secundarias (heterótomas).
Las trazas forman parte de la Icnofacies Cruziana que se da en la zona sublitoral (por debajo de las olas de buen tiempo y por encima del nivel base de las olas de tormenta (MacEachern, Pemberton, Gingras y Bann, 2007; Boggs, 2009), lo cual determina la disponibilidad de nutrientes. Esta icnofacies esta caracterizada por la presencia de diferentes trazas entre ellas tubos verticales, tubos en U inclinados, estructuras horizontales y trazas de organismos que se mueven dentro o sobre el sustrato (Mayoral et al., 2013).

Fig. 3: A) Cortes en diversos ángulos de los elementos elongados, debido a la inclinación de la traza con respecto al fondo. B) Cortes transversales de las galerías superpuestas que conforman las rosetas. C) Fragmentos de varias rosetas donde se observan los elementos radiales y sus ramificaciones. D) E).Cortes transversales de Ophiomorpha nodosa en facies de areniscas medias bien seleccionadas de la Formación Coris.
Las condiciones en que se conservan los restos de D. ottoi estudiados, coinciden con otros reportes de Dactyloidites en diferentes lugares del mundo, que se encuentran en areniscas limpias con HCS, en ambientes marinos someros (shoreface) (Uchman y Pervesler, 2007), depositados bajo el nivel base de las olas de buen tiempo y sobre el nivel base de olas de tormenta (Boyd y Mcllroy, 2016), condiciones similares han sido reportadas por (Lazo et al., 2008; López, y Ardévol, 1999), evidenciado también por la presencia de ondulitas y areniscas cuarzosas muy bien seleccionadas que sugieren un intenso retrabajo en un ambiente somero de alta energía afectado por tormentas ( Myrow y Southard, 1991). Típicamente la traza presenta una distribución de facies restringida y una amplia distribución espacial y temporal (López, y Ardévol, 1999).

Fig. 4 A) Roseta de D. ottoi que muestra los elementos radiales con su terminación roma y algunos spreites protrusivos, la coloración diferente en la parte interna.B).,D). Perdida de la laminación por retrabajo del sedimento. C). Cortes transversales de D. ottoi asociados a sedimentos con laminación paralela curva.
Gilbert et al (1995) reportan especímenes de D. ottoi en areniscas bien seleccionadas con estratificación cruzada que son interpretadas como dunas submarinas producidas en ambientes estuarinos someros. Además, mencionan que tanto la diversidad como la composición paleontológica sugieren un clima cálido en un ambiente litoral a sublitoral somero, coincidiendo con otros hallazgos de la icnoespecie. De igual forma López y Ardévol,1999, consideran que esta icnoespecie ha sido siempre asociada a sedimentos arenosos costeros con estratificación cruzada, tanto en el Cretácico Superior como en el Neógeno de varias localidades del mundo.
En general, todas las localidades que contienen restos de D. ottoi, están constituidas por areniscas limpias y los ejemplos provenientes de localidades del Neógeno se depositaron en ambientes marinos someros con fuerte energía hidrodinámica (Gilbert et al, 1995; Lazo et al, 2008). Estas coincidencias entre los diferentes afloramientos con D. ottoi, reafirman el hecho de que esta traza es un buen indicador de paleoambientes someros, energéticos y cálidos (Gilbert et al, 1995), principalmente en facies de areniscas limpias, afectadas por tormentas, las cuales frecuentemente presentan estratificación cruzada tipo HCS.
Agirrezabala y Gilbert (2004), mediante estudios sedimentológicos y análisis de facies detallados en un delta tipo Gilbert, concluyen que D. ottoi ocurre en estas condiciones a profundidades entre 0 a 3 m, en sustratos arenosos siliciclásticos ricos en materia orgánica, con tasas de sedimentación altas y discontinuas y algunas veces con salinidad reducida.
Productor
Los icnofósiles en forma de roseta o abanico pueden incluir trazas producidas por diferentes organismos o por la diferente conducta o comportamiento de un solo organismo, por ejemplo, reposo, habitación o vivienda y diferentes tipos de trazas de alimentación, tanto endogénicas como epigénicas (Gilbert et al., 1995), por lo que han sido interpretados de diversas maneras.
Originalmente fue considerada como un fósil de una esponja (López y Ardévol, 1999), como restos de medusas o de plantas (López y Ardévol, 1999) algas, trazas epigenéticas, poliquetos (Uchman y Pervesler, 2007), como trazas endogenéticas (Gilbert et al., 1995), pistas de alimentación de invertebrados tipo Fodicnia (López y Ardévol, 1999).
Fürsich y Bromley (1985) sugiere para D. ottoi un productor semejante a un gusano, que posiblemente posee una probocis para retrabajar el sedimento. La traza en conjunto es del tipo Fodicnia (Fürsich y Bromley, 1985) y muestra un patrón de conducta complejo de alimentación endógena. Además, el hecho de que las ramas estén muy cercanas y dispuestas casi paralelas y que casi nunca se crucen entre sí, sugieren una conducta fobotáctica (Agirrezabala y De Gilbert, 2004), que genera una explotación altamente eficiente del sustrato disponible, que no muestra perturbación del sedimento entre las galerías superpuestas (Fürsich y Bromley (1985). Las rosetas exactamente superpuestas permiten explotar el sedimento completamente en dirección vertical, y gracias a abarcar sólo dos tercios de círculo, las rosetas adyacentes pueden cubrir el espacio en la horizontal casi en su totalidad (Fürsich y Bromley (1985); López y Ardévol, 1999).
Lazo et al (2008), mencionan que las trazas con forma de roseta o abanico reflejan una búsqueda de alimento más estacionaria y proponen que pudieron ser formadas por poliquetos o bivalvos depositívoros sifonados como lo había propuesto Bromley et al., (2003). Gilbert et al, 1995, mencionan que el gusano poliqueto Arenicola marina vive en ambientes marinos someros comparables con los que contienen los icnofósiles D. ottoi, por lo que se puede asumir que el productor de la traza puede ser un vermiforme parecido a Arenicola y con requerimientos ecológicos y comportamiento similares. El carácter gregario de D. ottoi, es consistente también con un productor semejante a Arenicola.
Aunque no se conoce con seguridad el organismo productor, la mayoría de los autores suponen que sería un organismo vermiforme, que podría tener una probóscis extensible y parte del animal se mantendría en el eje central (Pickerril, Donovan y Dixon, 1993; López y Ardévol, 1999). D. ottoi es una traza tipo fodicnia producida por un gusano depositívoro en areniscas silicoclásticas (Pickerill et al., 1993)
Conclusiones
Se describe por primera vez la traza D. ottoi, para la Formación Coris del Mioceno y para la estratigrafía de Costa Rica en general. Esta traza está constituida por rosetas o abanicos, formadas por elementos elongados dispuestos radialmente, que conforman galerías superpuestas y que salen de un conducto central.
D. ottoi es un indicador de ambiente marino somero, principalmente upper shoreface, de alta a mediana energía, caracterizado por facies de areniscas limpias, afectadas por tormentas con laminaciones paralelas planas y curvas e inclinadas de bajo ángulo (HCS).
D. ottoi está restringido a la facies y solo se desarrollan en sustratos arenosos someros e inestables, con materia orgánica, altas tasas de sedimentación, depositación episódica que permite cortos períodos de colonización y retrabajo.
Los icnofósiles D. ottoi son raros y ocurren en conjuntos de baja diversidad, junto a otras trazas como Ophiomorpha nodosa. Son estructuras de alimentación complejas tipo fodicnia utilizadas para buscar eficientemente el alimento dentro del sedimento, producidas posiblemente por un organismo vermiforme, forma parte de la Icnofacies Cruziana.

