











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La familia Tayassuidae es un grupo de artiodáctilos cuya morfología recuerda a los cerdo o jabalíes del Viejo Mundo, pero se diferencian de estos últimos, en términos generales, por poseer cráneos más bajos y cortos con la porción trasera característicamente plana; sus hocicos son angostos y sus caninos son notablemente desarrollados para un mamífero no carnívoro, tanto los incisivos inferiores como los superiores, se orientan dorso ventralmente y no se tuercen hacia arriba, atrás o lateralmente como en los cerdos (Kurtén y Anderson, 1980[1]; Scott, 1913[2]; Sowls, 1997[3]; Wright, 1991[4], 1998[5]).
Los tayassuidos en la actualidad se distribuyen desde el suroeste de Estados Unidos hasta la parte norte central de Argentina, propiamente la región del Chaco, abarcando hábitats tan variados como los bosques lluviosos tropicales y subtropicales, bosques nubosos y secos, desiertos, bosques de mezquite, bosques de galería, matorrales, algarrobales, sabanas y hábitats alterados (Carrillo, Wong y Sáenz, 1999[6]; Eisenberg y Redford, 1999[7]; Sowls, 1997[3]).
Los tayassuidos están representados por 3 especies vivientes, a saber: Tayassu pecari, Pecari tajaçu y Catagonus wagneri, este último género previamente conocido únicamente a partir del registro fósil de América del Sur (Ameghino, 1908[8]; Dutra, Casali, Missagia, Gasparini, Perini & Cozzuol, 2016[9]; Eisenberg y Redford, 1999[7]; Prothero, 2009[10], 2015[11]; Reig, 1952[12]; Wetzel, Dubos, Martin & Myers, 1975[13]). Cabe señalar que una cuarta especie de Tayassuidae el Pecari maximus fue descrita por van Roosmalen et al., 2007[14] para la cuenca amazónica, sin embargo, su validéz taxonómica ha sido cuestionada por Gongora et al. (2011)[15], quienes consideran que en realidad se trata de individuos notablemente grandes de la especie Pecari tajaçu.
El registro fósil de la familia Tayassuidae en América del Norte inició en el Eoceno tardío a partir de un ancestro euroasiático que atravesó Beringia en esa época (Hulbert, 2001[16]; Romer, 1966[17], Wright, 1998[5]), llegando a desarrollar una gran diversidad taxonómica en dicho subcontinente; se sabe que alcanzó América Central meridional en el Mioceno superior o Hemingfordiano temprano (MacFadden et al., 2010[18], 2014[19]) y arribó a América del Sur en el Plioceno Medio, hace 3,3 a 4 Ma en el Chapadmalalano tardío (Prevosti, Gasparini & Bond, 2006[20]) al inicio del Gran Intercambio Biológico entre las Américas (Marshal et al., 1984[21], Gasparini, Kerber & Oliveira, 2009[22]). Adicionalmente, Frailey y Campbell (2012)[23] describen 2 nuevos géneros y especies de Tayassuidae para el Mioceno Superior de la Amazonia peruana, asignándoles una edad Chasicoano–Huayqueriano, aproximadamente unos 9.0 Ma por correlación geológica, sin embargo, dicha edad es cuestionada y aún debe ser verificada (Dutra et al., 2016)[9].
La subfamilia Tayassuinae que es la única subfamilia con representantes actuales, comprende 35 especies (Doughty, 2016)[24], incluidas las 3 existentes, y se caracteriza por su alta diversidad morfológica y de especies y su amplísima distribución geográfica y temporal, Barstoviano al Reciente de Norteamerica y Chapalmalalano al Reciente de América del Sur (Wright, 1991[4], 1998[5]).
Los objetivos de la presente publicación es dar a conocer el primer registro fósil de pecaríes para Costa Rica y describir el material dental, craneal y post-craneal de Tayassuinae colectado en la localidad de San Gerardo de Limoncito, ubicada 14 km al oeste de la ciudad de San Vito en el cantón de Coto Brus, provincia de Puntarenas en las coordenadas 8º51’19.6’’333 N y 83º04’51.9’’ W (Fig. 1), cuya edad ha sido asignada al Hemphilliano Temprano tardío (Mioceno Tardío).
Metodología
Las medidas biométricas lineales y dentales en milímetros se realizaron con calipper digital de acuerdo con la metodología propuesta por Von den Driesch (1976)[25], Doughty (2016)[24] y Doughty et al. (2018)[26], ver cuadro 1 y cuadro 2.
La descripción de los dientes se basó en la nomenclatura dental propuesta por Doughty et al., 2018 quienes a su vez la modificaron de Wright y Webb (1984)[27] y Wright (1991)[4].
Los datos obtenidos se registraron y graficaron en Microsoft Excel, a ellos se añadieron datos biométricos dentales compilados y utilizados por Doughty (2016)[24], Doughty et al. (2018)[26], Prothero y Grenader (2012)[28], Prothero y Sheets (2013)[29], Wright (1989)[30] y Wright y Webb (1984)[27].
La base de datos incluyó únicamente aquellos géneros y especies de Tayassuinae restrictos al Hemphilliano tardío Hh 3 y Hh 4 de América del Norte, a saber: Protherohyus brachydontus Mooser y Dalquest (1980)[31], Platygonus pollenae Prothero y Grenader (2012)[28], Mylohyus elmorei White (1942) y Prosthennops serus (Cope, 1877)[32] Gidley (1904)[33]; expresados en varios gráficos, como se muestra en lafigura 2.
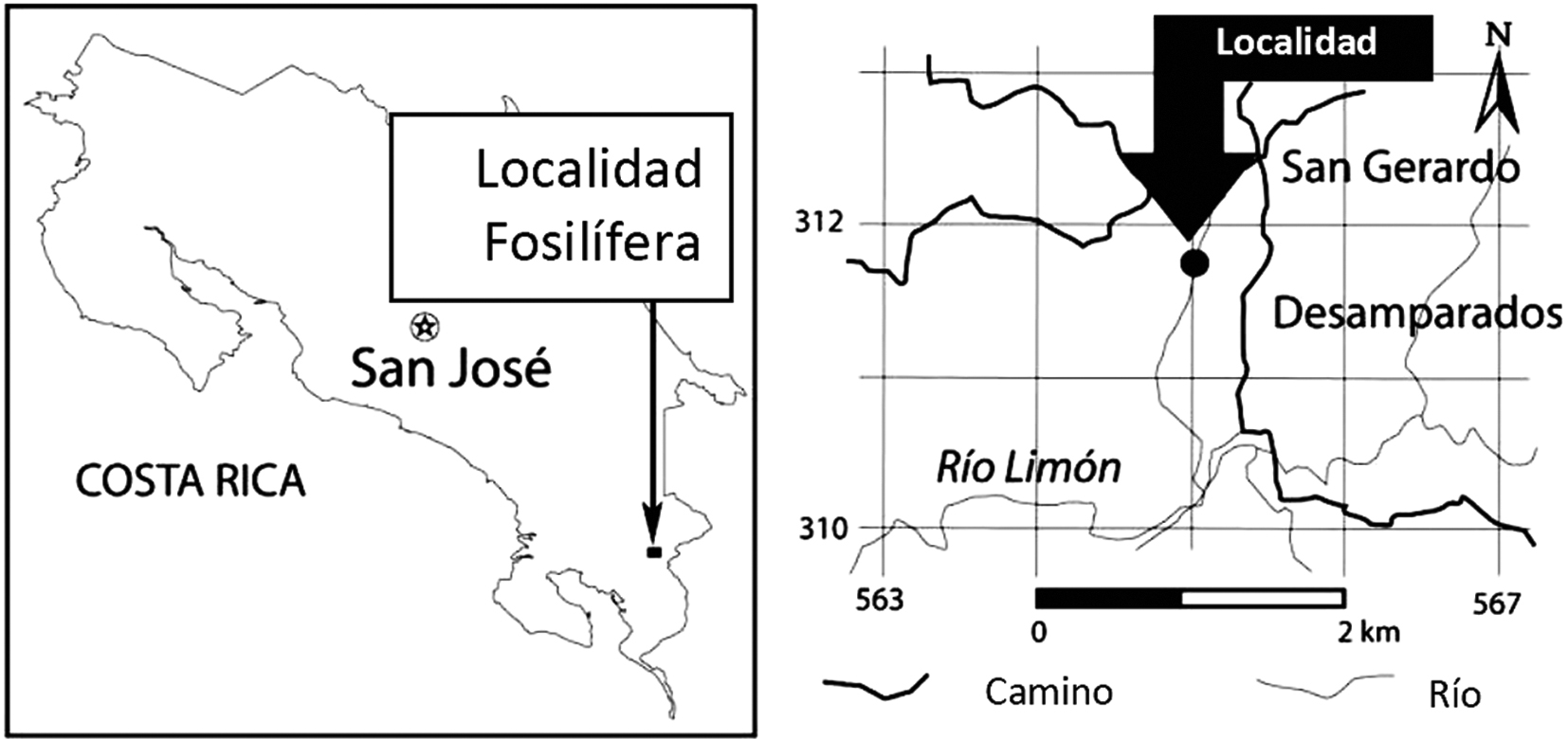
Figure 1 Mapa de ubicación de la localidad fosilífera de San Gerardo de Limoncito, cantón de Coto Brus, provincia de Puntarenas.
Cuadro 1 Datos biométricos dentales de los Tayassuidae fósiles de San Gerardo de Limoncito.
Cuadro 2 Datos biométricos del material post-craneal de los Tayassuidae fósiles de San Gerardo de Limoncito.
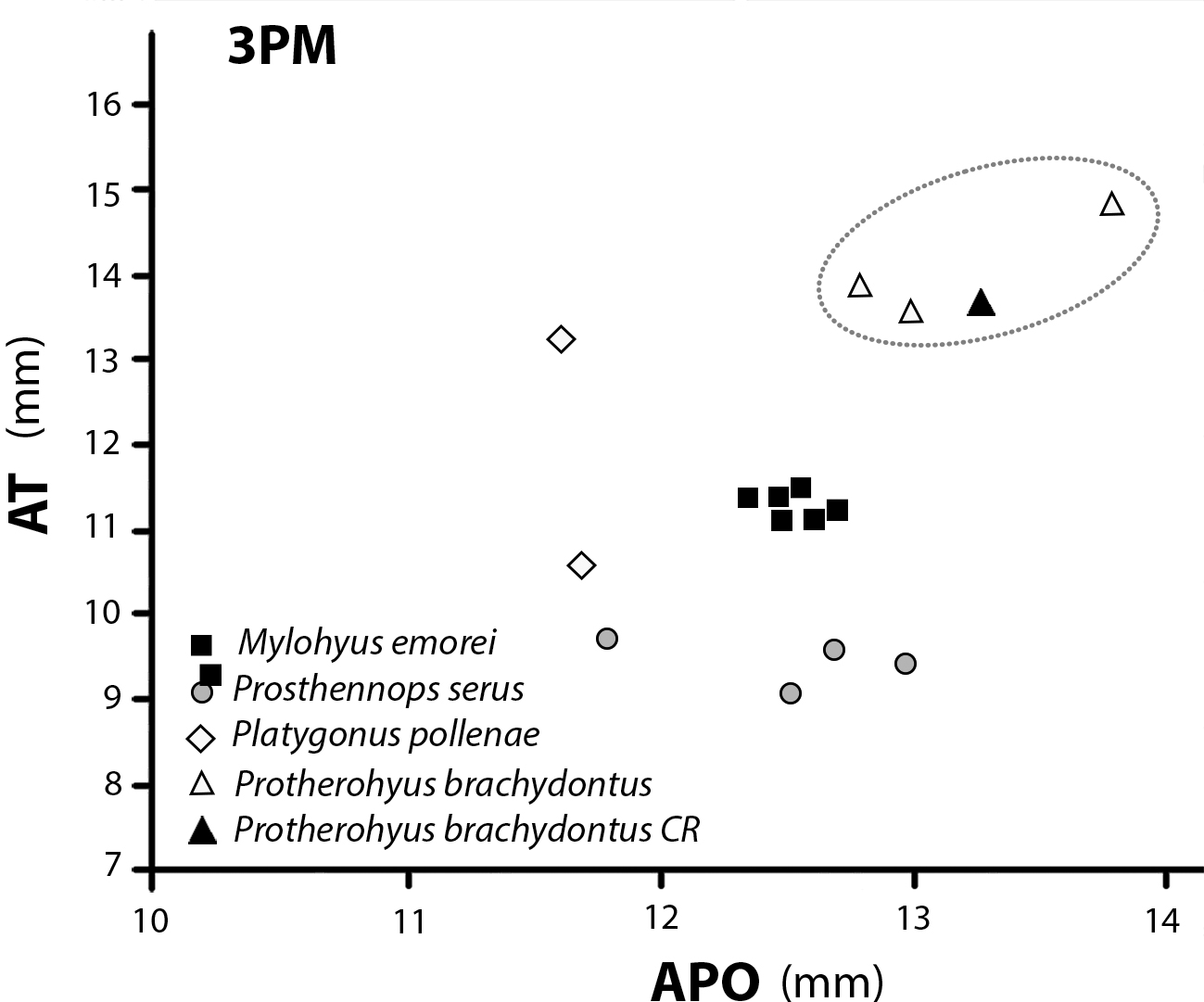
Fig. 2 Gráfico de dispersión de APO, longitud anteposterior en mm vs. AT, ancho transversal de los terceros premolares superiores de 4 especies de Tayassuidae del Hemphilliano de Norte América, comparadas con el ejemplar CFM-2195, representado por un triángulo negro que coincide con el ámbito de tendencia y dimensiones de Protherohyus brachydontus Dalquest y Mooser (1980).
Materiales
Ejemplar CFM-2915, es un fragmento de ramo mandibular derecho con pm4-m3 bien preservados el m1 esta roto y solo se preservaron las cúpides anteriores (Fig. 3). El CFM-1861 corresponde a un fragmento mandibular derecho con el m2 muy bien preservado (figure 4. 1a c). El CFM-1862 corresponde a la corona de un 2m (Fig. 4.2a-c) y el CFM-1499 a un m2 (Fig. 4.3a-c), ambos bien preservados.Ejemplar CFM-2195 corresponde a una corona de un premolar superior izquierdo 3PM con avanzado desgaste (Fig. 4.4a-b).
Ejemplares CFM-1652 (Fig. 5 1a-d) y CFM-1651 (Fig. 5 2b) corresponden a 2 caninos superiores, el ejemplar CFM-1867 correspondiente a un tercer incisivo superior (fig.5 5a-b) y dos fragmentos sinfisiales correspondientes a los ejemplares CFM-1651 y CFM-1652 (Figs. 5.2 y 5.1).
Fragmentos post-craneanos
Escasos fragmentos del post-cráneo de tayassuidos fueron recuperados en la localidad de San Gerardo de Limoncito. El CFM-3102 es una falange proximal (Fig. 9) y el CFM-3056 y el CFM 1260 corresponden a 2 fragmentos de húmero izquierdo (Fig. 10.1 y 10.2), el CFM-3842 es un fragmento radio ulnar (Figs. 10a-c).Todos los materiales se encuentran depositados en la Colección de Fósiles de la Sección de Geología del Departamento de Historia Natural del Museo Nacional de Costa Rica, bajo el acrónimo CFM.
Palentología
Clase MAMMALIA Linnaeus (1758)
Orden ARTIODACTYLA Owen (1848)
Suborden SUIFORMES Jaeckel (1911)
Infraorden SUOIDEA Gray (1821)
Familia TAYASSUIDAE Palmer (1897)Subfamilia TAYASSUINAE Palmer (1897)
Tribu PLATYGONINI Perini et al. (2016)
Género Protherohyus Perini et al. (2016)
Protherohyus brachydontus Dalquest y Mooser (1980) Perini et al. (2016)Hemphilliano Tardío Hh3-Hh4
Sinonimia
- Desmathyus brachydontus n. especie Dalquest y Mooser (1980, pp. 15-17)[31].
- Catagonus brachydontus (Dalquest y Mooser, 1980)[31], Wright (1989, p. 291)[30].
Descripción
3PM: premolar no molariforme trirradiculado, con corona muy desgastada y de contorno más o menos redondeado, la cara lingual es notablemente redondeada en tanto que la cara labial es ligeramente recta, presenta un conspícuo cíngulo anterior. Las cúspides presentan una condición seudomolariforme, es decir con cuatro cúspides, con un protocono muy amplio de contorno redondeado y contiene una amplia foseta cicular; el hipocono es alargado en sentido linguo-vestibular y angosto en sentido mesio-distal con un contorno triangular y presenta una pequeña foseta; el paracono posee un contorno rectangular con una foseta angosta, está roto, pero sus dimensiones son similares al protocono; el metacono tiene un contorno triangular, es relativamente pequeño y a pesar de que el diente está muy desgatado el metacono porta la única cúspide observable con una foseta diminuta (Figs. 4. 4a y 4b).

Fig. 3 CFM-2915, Prosthennops serus (Cope, 1877), fragmento de ramo mandibular derecho en a vista oclusal, b vista lingual y c vista vestibular. Escala visual 1 cm.

Fig. 4 Prosthennops serus (Cope, 1877)[32] 1. CFM-1861 corresponde a un fragmento de mandíbula derecha con un m2 preservado; 2. CFM-1862 corresponde a la corona de un 2m y 3. CFM-1499 corresponde a un m2, en todos los casos las vistas son: a oclusal, b lingual y c labial. Protherohyus brachydontus Dalquest y Mooser (1980)[31] 4. el CFM 2195 corresponde a un 3PM a. vista oclusal y b. detalle de la raíz, ant anterior, pos posterior, li lingual y la labial o vestibular.

Fig. 5 Prosthennops serus (Cope, 1877)[32], fragmentos sinfisiales el 1. CFM 1652, en vistas a. dorsal, b. ventral, c. y d. laterales, 2. CFM-1651, en vistas a. dorsal y b. lateral. 3. y 4. Fragmentos de caninos superiores en vistas a. labial, b. lingual y c. comisural. 5. Primer incisivo superior derecho en vista a. lingual y b. comisural.
Discusión
Los premolares superiores de Protherohyus brachydontus Dalquest y Mooser (1980)[31] se destacan por no ser molariformes, incluso los cuartos premolares.
El ejemplar CFM-2195 es un 3PM que, aunque muy desgastado, se caracteriza por su talla comparativamente más grande que los restantes molares de Tayassuidae colectados en San Gerardo de Limoncito, el contorno redondeado de su corona que, a pesar de estar desgastada, muestra una base notablemente ensanchada y desplazada lingualmente, y la presencia de un amplio cíngulo anterior. Todas estas características son propias de los premolares superiores terceros de Protherohyus brachydontus Dalquest y Mooser (1980)[31] y no se diferencian de los ejemplares p.e. de Bone Valley ilustrados por Wright (1989)[30] en las figuras 2a y 2b, Pipe Creek Sinkhole, Indiana ilustrado por Prothero y Sheets (2013) en la Fig. 1 o el especimen de Buis Ranch, ilustrado en la Online Repository of Fossils (UMORF) of The University of Michigan.
Por otra parte, los datos biométricos del ejemplar CFM-2195 coinciden dentro del ambito de dispersión para los terceros premolares superiores de Protherohyus brachydontus Dalquest y Mooser (1980)[31], ver Figure 2.
Distribución paleobiogeográfica
En Estados Unidos la especie Protherohyus brachydontus Dalquest y Mooser (1980)[31] se ha registrado para el Mioceno Superior, en el Hemphilliano Hh3 de Coffee Ranch en Texas, Hh4 de Buis Ranch en Texas y Palmetto en La Florida (Webb et al., 2008[34], 1998), Hh3-Hh4 de Bone Valley en La Florida y Hh4 Pipe Creek Sinkhole Local Fauna, Indiana (Prothero y Sheets, 2013)[29].
La especie Protherohyus brachydontus Dalquest y Mooser (1980)[31] se ha descrito para el Mioceno Superior de México, en el Hemphilliano tardío de la Cuenca de San Miguel Allende en las localidades Hh4 de Rancho El Ocote, Rinconada, Arroyo Tepalcates y Hh3 de Coecillos; en la Cuenca de Tepeji del Río en la localidad Hh4 de Plegaria en el Estado de Hidalgo, en las localidades de Hh3 Santa María y Hh4 La Hacienda en la Cuenca de Tecolotán en el Estado de Jalisco y la localidad de Hh3 Chincua en el Estado de Michoacán (sensu Carranza-Castañeda et al., 2013[35]; Dalquest y Mooser, 1980[31]).
En América Central el presente hallazgo de la localidad de San Gerardo de Limoncito con una edad confirmada Hh3 constituye el registro más austral de la especie.
Tribu TAYASSUINI Perini et al. (2016)Género Prosthennops Gidley (1904)
Prosthennops serus (Cope, 1877) Gidley (1904)Clarendoniano Tardío – Hemphilliano Tardío temprano Cl3 – Hh3
Sinonimia
- Dicotyles serus en Cope (1877, p. 224 – 225)[32]
- Prosthennops serus Cope (1878, en Matthew y Thomson, 1924, p. 179)[36]
- Prosthennops (Prosthennops) serus Cope (1878, en Skinner et al., 1977, p. 359)[37]
- Prosthennops graffhami Schultz y Martin (1975, pp. 35-36)[38]
- Prosthennops serus Cope (1878, en Wright (1998, p. 397)[5]
Descripción
i2i-i3i: fragmentos de las coronas de estos incisivos están presentes en la sínfisis del ejemplar CFM-1652 (Fig. 5. 1b).1c: se preserva parte de la corona del canino inferior izquierdo en el ejemplar CFM- 1652, la cual es ligeramente abultada y no tan comprimida en sentido linguo-vestibular (Fig. 5. 1a-d).
c1: se preserva parte de la corona del canino inferior derecho en la cavidad alveolar de la sínfisis del ejemplar CFM- 1651 (Fig. 5. 2b).
I1: el ejemplar CFM-1867 (Fig. 5. 5a-b) se halla roto en la porción distal de la raíz, es ovoide y muestra poco desgaste en su superficie oclusal.
C1: se preservan 2 fragmentos de caninos superiores derechos con la cara lingual de la corona ligeramente más plana que la cara labial ligeramente que es más inflada. El CFM-2637, aunque roto se caracteriza por su gran tamaño, ser comprimido en sentido linguo-vestibular y presentar filos agudos y lisos en los bordes anterior y posterior, y el CFM-5397, corresponde a un fragmento anterior de corona con filo liso (Figure 4, Figure 5).
pm4: premolar molariforme, cuadrirradiculado con corona rectangular y cúspides medianamente desgastadas. El protocónido y el metaconido presentan un contorno cuasicircular, siendo el protocónido ligeramente más elevado; el hipocónido y el entocónido son bastante más pequeños, ambos de contorno subcircular; el hipoconulido tiene un contorno triangular y el entoconulido tiene un contorno circular. Presenta un conspicuo cíngulo anterior (Fig. 3).
m1: molar roto del cual solo se preservó el lofo distal con el protocónido y el metacónido medianamente desgastados, siendo el primero ligeramente más elevado (Fig. 3).
2m2: los segundos molares inferiores presentan un contorno general rectangular con el eje de los lofos anteriores y posteriores paralelos entre si y perpendiculares con respecto al eje anteroposterior del diente. De las cúspides labiales el protocónido es la cuspide más alta, seguida por el hipocónido y ambas son en general más elevadas que las cúspides linguales siendo el metacónido ligeramente más alto que el entocónido, ello es particularmente notable en los ejemplares CFM-1861 y CFM-1862 (Figs. 4.1 y 4.2) con desgaste incipiente de la corona. Tanto el hipoconulido que tiene un contorno triangular y el entoconulido de contorno elíptico son muy reducidos y poco prominentes, observables únicamente en estados de desgaste avanzado como en el ejemplar CFM-2915 (Fig. 3) y desaparecen en estados de desgaste muy avanzados como en el ejemplar CFM-1499 (Fig. 4.3), donde todas las cúspides principales presentan figuras de desgaste o fosetas triangulares a manera de tréboles simples. Estos molares no presentan cíngulo labial o vestibular, pero si presentan cíngulo mesial y distal, siendo más evidente el mesial. Las coronas en sus bases pueden presentar crenulaciones cortas muy sutiles y poco profundas en el esmalte.
Discusión
La morfología de la corona de los molares inferiores, incluyendo el cuarto premolar, de Prosthennops serus es la menos bunodonta y notablemente bilofodonta (Fig. 4.1 y 4.2) en comparación con la observada en las especies Mylohyus elmorei, Platygonus pollenae y Protherohyus brachydontus. Los cíngulos en los molares inferiores de Prosthennops serus solo están presentes en las caras mesial y distal, en esta última formando un pequeño talónido, además son muy rectos y no se proyectan lingual o labialmente como en el caso de Platygonus pollenae, Mylohyus elmorei y Protherohyus brachydontus, siendo en estas dos últimas especies particularmentes curvos lo que le imprime a los molares un contorno más redondeado, en tanto que el contorno de los dientes inferiores desde el 2p2 al 2m2 son notablemente rectangulares y marcadamente lofodontes, p.e. ver los ejemplares con coronas con poco desgaste como CFM-1861 y CFM-1862 (Fig. 4.1 y 4.2).
El pm4 presente en la hemimandíbula derecha (CFM-2915) presenta dimensiones AT y APO que lo ubican fuera del rango de Mylohyus elmorei y Platygonus pollenae y lo ubican dentro de la nube de Protherohyus brachydontus y Prosthennops serus (Fig. 6 gráfico superior). En el caso del m1, aunque roto, se estimó su longitud APO y se midió su longitud AT, permitiendo ubicarlo en el rango de tendencia de Prosthenops serus (Fig. 6 gráfico inferior), dejándolo fuera del rango de las restantes especies hemphillianas tardías de tayassuidos.
Los segundos molares inferiores están representados por 4 especímenes y presentan dimensiones anteroposteriores y transversales que permiten diferenciarles facilmente de otras especies de tayassuidos del Hemphilliano tardío como por ejemplo de Protherohyus brachydontus Dalquest y Mooser (1980)[31], Platygonus pollenae Prothero y Grenader, 2012[28] y Mylohyus elmorei White (1942) (Fig. 7). A pesar de que los molares inferiores presentan dimensiones ligeramente por debajo de sus similares descritos en América del Norte para la especie Prosthennops serus (Cope, 1878), con excepción del ejemplar CFM-1862, todos caen dentro del rango de tendencia de esta especie. Cabe señalar que el ejemplar CFM-1862 es indiferenciable morfológicamente de los restantes segundos molares inferiores de P. serus, aunque es notable por su relativo gran tamaño de un 11,88 a un 12,45% de longitud anteroposterior más grande que los ejemplares más pequeños.
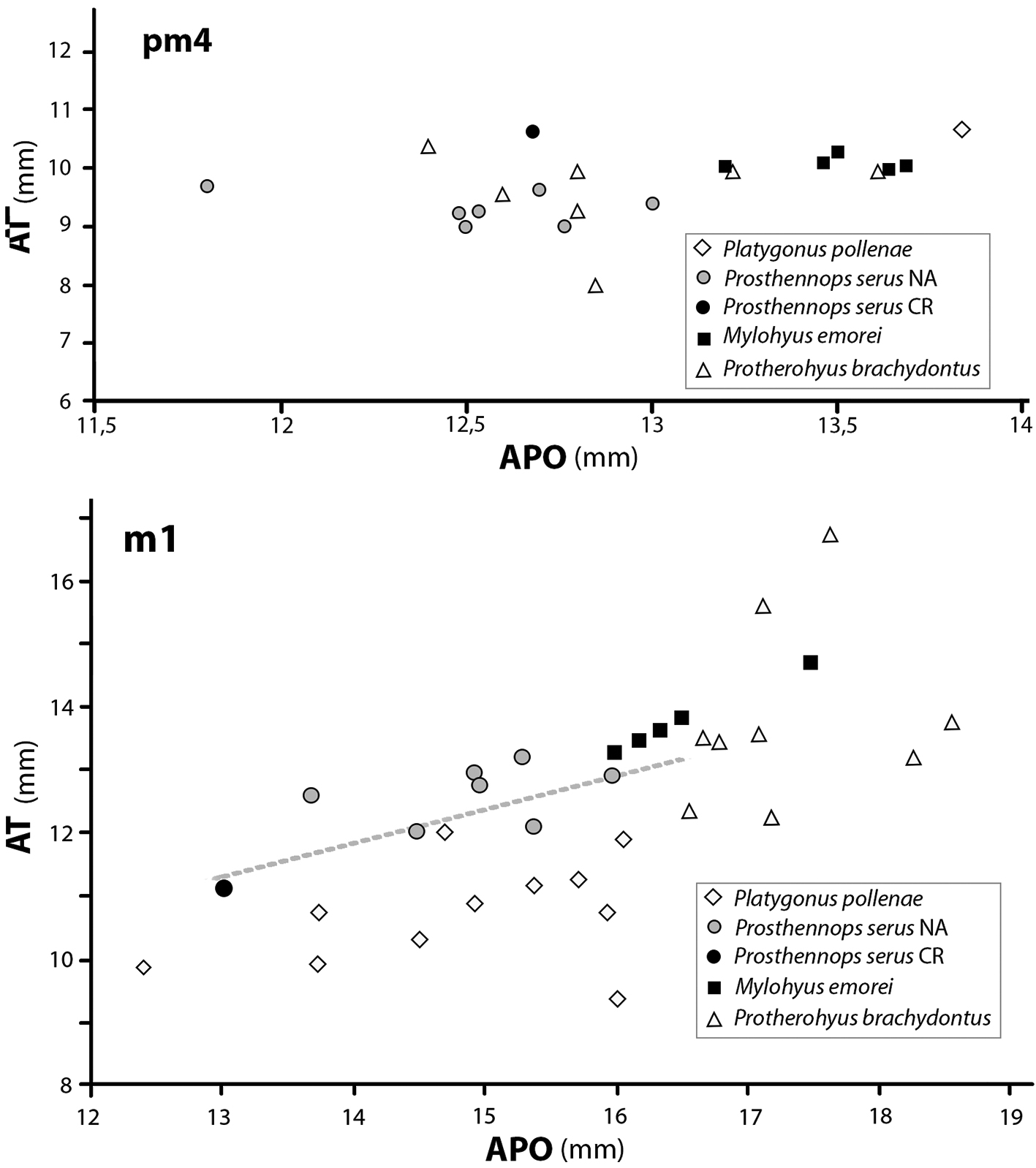
Fig. 6 Gráficos de dispersión de APO longitud anteposterior vs. AT ancho transversal de los cuartos premolares y primeros molares inferiores de 4 especies de Tayassuidae del Hemphilliano de Norte América comparadas con los similares de la muestra CFM-2915.
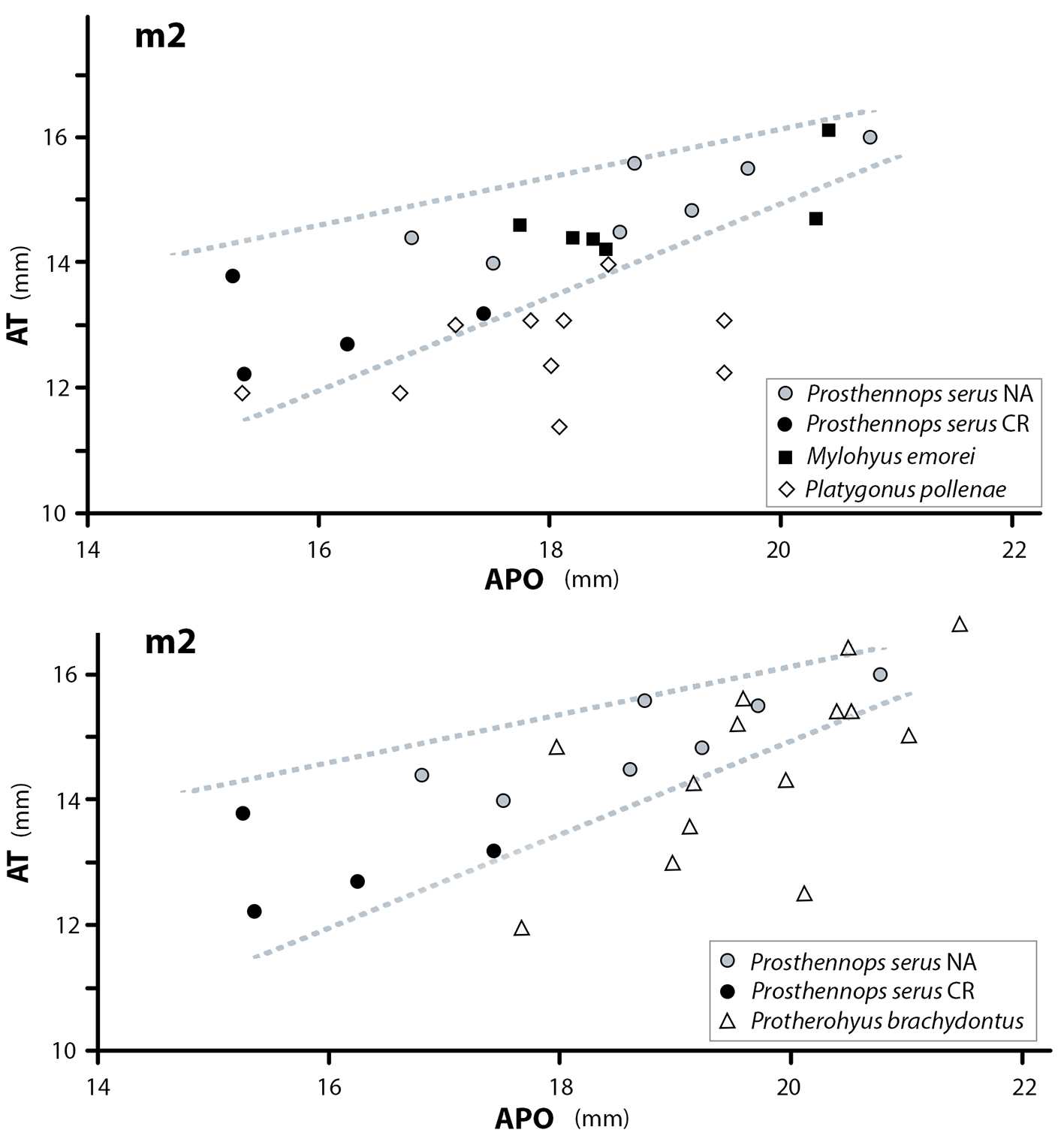
Fig. 7 Gráficos de dispersión de APO longitud anteposterior vs. AT ancho transversal de los segundos molares inferiores de 4 especies de Tayassuidae del Hemphilliano de Norte América comparadas con los ejemplares CFM-1499, CFM-1861, CFM-1862 y CFM-2915.
El m3, por su parte, presenta dimensiones que son compatibles con la línea de tendencia de los 3m3 de Prosthennops serus (Cope, 1878) (Fig. 8, muy próximo al rango más pequeño registrado para individuos de América del Norte, específicamente al ejemplar de la University of Nebraska State Museum, con el código 36 UNSM 76052T cf. Schultz y Martin (1975)[38], compilado por Doughty (2016)[24].
Material postcranial de Tayassuini
Descripción: el CFM-3102 corresponde a una falange proximal completa con la superficie proximal de contorno redondeado más o menos oval (Fig. 9).
Dos fragmentos de húmeros izquierdos, el CFM-1260 corresponde al fragmento de menor talla y el CFM-3056 al de mayor talla, en el caso del primer ejemplar la fosa olecraneana se preservó y presenta un contorno triangular; en ambos ejemplares el epicóndilo medial es angosto y alargado, y el epicóndilo lateral es corto y relativamente angosto, la tróclea es realtivamente ancha latero-medialmente con la superficie patelar en forma de reloj de arena.
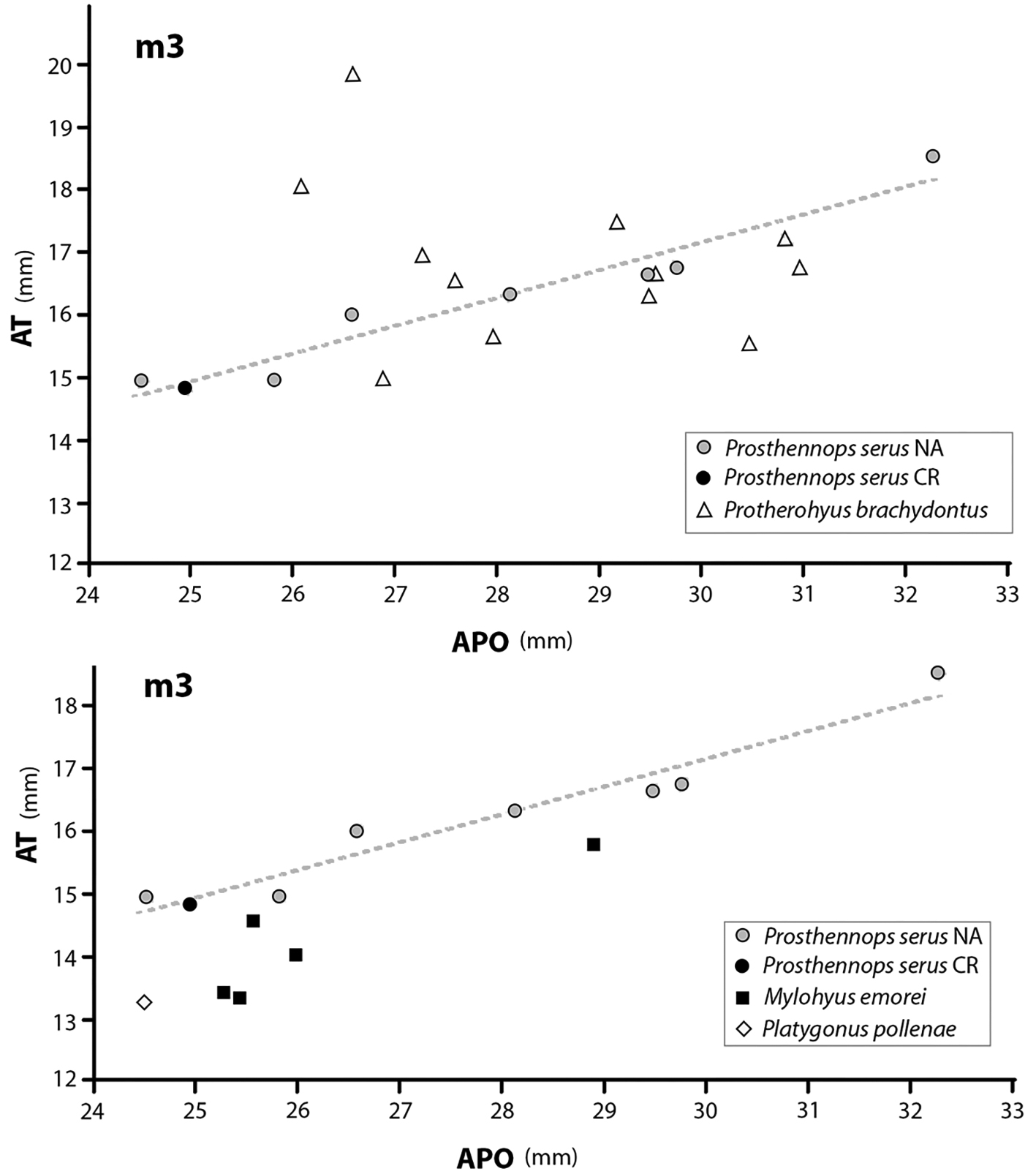
Fig. 8 Gráficos de dispersión de APO longitud anteposterior vs. AT ancho transversal de los terceros molares inferiores de 4 especies de Tayassuidae del Hemphilliano de Norte América comparadas con el ejemplar del fragmento de mandíbula CFM-2915.
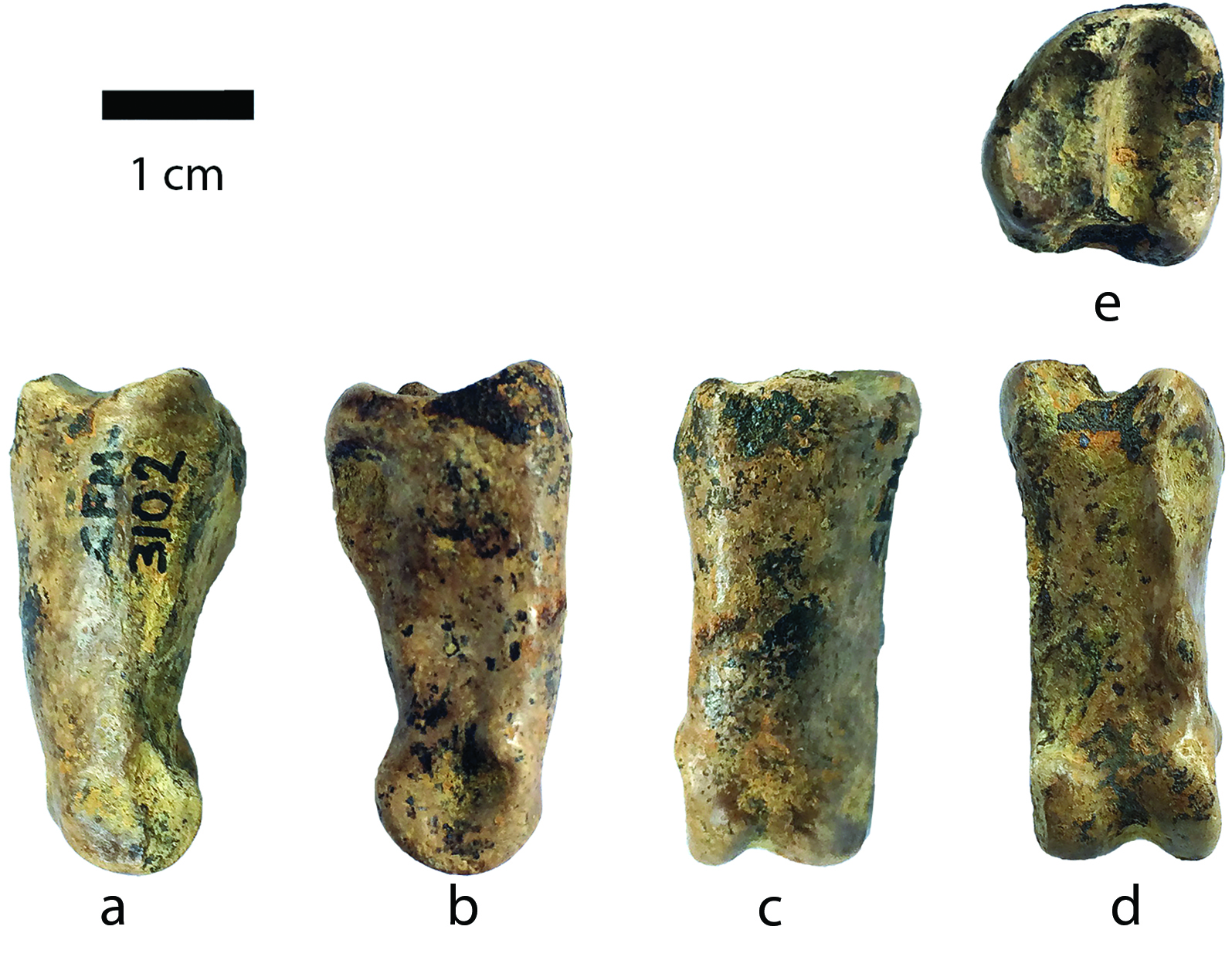
Fig. 9 Prosthennops serus CFM-3102 falange proximal I del dígito III del pie izquierdo en vista a. lateral, b. medial, c. anterior, d. posterior y e. proximal.
El CFM-3842 corresponde a un fragmento proximal de un radio-ulnar en el que se observa parte de la escotadura troclear y la fosa capitular. El especimen es notable por mostrar numerosas marcas de gnatichnia.
Discusión: a nivel de post-cráneo la presencia de una ulna y un radio fusionados es uno de los caracteres típicos de la familia Tayassuidae sensu Scott (1913)[2] y Kurtén y Anderson (1980)[1], pero no podríamos determinar a cuál de las 2 especies pertenece, debido a lo fragmentario del material (Fig. 10. 3a-c).
En el caso de los fragmentos distales de húmeros izquierdos, es posible deducir que se trata de 2 individuos de tallas ligeramente diferentes, el ejemplar CFM-1260 (Figs. 10.2a-d)Figure 10 es en general más pequeño, pero su tróclea es ligeramente más robusta que la que presenta el ejemplar CFM-3056 cuya talla general es ligeramente mayor (Figs. 10. 1a-d)Figure 10.
Distribución paleobiogeográfica
El primer registro fósil conocido para América Central de un Tayassuidae corresponde a Woodburne (1969)[39] quien registró la especie Pecari tajaçu (= Dicotyles tajaçu) (Linaeus, 1758), el cual fue colectado por Barnum Brawn en 1949, en una barra de punto en el lecho del Río de La Pasión a 2 km al sur de la desembocadura del Río Santa Amelia en el departamento de Petén, en sedimentos del Pleistoceno tardío.

Fig. 10 Fragmentos distales de húmeros izquierdos, ejemplares 1. CFM-3056 y 2. CFM-1260 en vista a. posterior, b. distal, c. medial y e. lateral. 3. CFM-3842, fragmento proximal de radio-ulnar izquierdo en vistas a. craneal y b. lateral, c. detalle del recuadro de la superficie donde se observan marcas de gnatichnia.
El segundo registro de un Tayassuidae fósil corresponde a Webb y Perrigo (1984)[40], quienes describieron un M3 de Prosthennops cf. P. serus, procedente de la localidad Las Culebras de la Formación Gracias de edad Hemphilliano temprano Hh1.
El tercer registro de un Tayassuidae fósil en la región corresponde a la especie “Cynorca” occidentale Woodburne, 1969[39], el material fue colectado en la localidad de Centenario, en la Formación Cucaracha y descrito por MacFadden et al., 2010[18]. La fauna de Centenario se le ha asignado una edad NALMA Hemingfordiano temprano (He 1), 18,8 a 19,04 Ma (cf. MacFadden et al., 2014).
La especie Prosthennops serus (Cope, 1877)[32] de acuerdo a Doughty et al. (2018)[26] ha sido registrada en el Mioceno Superior de América del Norte para el Clarendoniano temprano de “Loup Fork beds” en Sappa Creek, Republican River en el noroeste de Kansas (Matthew et al., 1918[41], Colbert, 1935[42]) Mioceno Superior de la Formación Ogallala cf. Wright (1998)[5], el Hemphilliano temprano de Malheur County, Oregon (Colbert, 1938)[43], el Hemphilliano temprano a tardío temprano de Nebraska (Hesse, 1935[44]; Schultz & Martin, 1975[38]), el Hemphilliano temprano tardío de Mauvilla Local Fauna en Alabama (Hulbert & Whitmore, 2006)[45], en esta localidad se registra Prosthennops cf. serus, fundamentado en un P4 (Hulbert y Whitmore, 2006)[45]. El Hemphilliano temprano de la localidad de Tynner Farm, centro norte de Florida, Hulbert, Morgan y Kerner (2009)[46] y el Hemphilliano tardío – Blancano temprano de Gray Fossil Site en los Apalaches de Tennessee (Doughty et al., 2018)[26] (Fig. 11).
Otras localidades citadas por Wright (1998)[5] portadoras de material comparable a Prosthennops serus son Deer Lodge Basin del Hemphiliano temprano de Montana; Higgins Local Fauna de edad Hemphilliano Temprano tardío de Texas y Wray Fauna del Hemphilliano tardío temprano de Colorados en su Doughty (2016)[24] y Doughty et al., 2018[26].
En América Central cabe destacar el registro fósil previo de un 3M, muy desgastado, de Prosthennops serus recuperado en sedimentos de la Formación Gracias en Honduras y de edad Hemphilliano temprano Hh1 (Webb y Perrigo, 1984)[40] y el presente registro de Prosthennops serus para el sur de Costa Rica, en el Mioceno Superior de la Formación Curré, procedente de la localidad de San Gerardo de Limoncito de edad Hemphilliano tardío temprano Hh3.
Localidades con material descartado, previamente asignado a Prosthennops serus (Cope, 1878), como: Coffee Ranch Fauna en Texas (aproximadamente 6.6 Ma sensu Passey et al., 2002[47]), Doughty et al. (2018)[26] consideran que el material fósil descrito para esta localidad debe ser reasignado a otra especie; los materiales del Hemphilliano tardío – Blancano del estado de Hidalgo y Rancho El Ocote en el estado de Guanajuato (cf. Wright, 1998)[5]; con relación a estos registros es importante agregar que aunque Wright, 1998[5] los asume como P. serus, Carranza Castañeda et al., 2013[35], describen el primer registro como Prosthennops sp.? para la localidad de Zietla, cuenca de Zacualtipán en el estado de Hidalgo y el segundo registro como Protherohyus brachydontus para las dos localidades de Rancho El Ocote, por lo que al parecer no existe un registro de la especie Prosthennops serus en México.
Conclusiones
Los elementos dentales permiten reconocer claramente 2 especies de Tayassuidae en el Hemphilliano tardío del sur de América Central, Prosthennops serus (Cope, 1878) y Protherohyus brachydontus Dalquest y Mooser (1980)[31], recuperados en la localidad de San Gerardo de Limoncito y con una edad radiométrica de 5,8 ± 0,60/-0,80 Ma (ver apéndice). Ello además, sugiere un patrón simpátrico de distribución.
Los especímenes de Prosthennops serus (Cope, 1878) de la localidad de San Gerardo de Limoncito son proporcionalmente los más pequeños de la especie, en contraposición con los ejemplares de Tyner Farm y Gray Fossil Site, que son los que muestran las proporciones más grandes (sensu Doughty et al., 2018)[26]. Este rango de dimensiones confirma la gran variabilidad intraespecífica previamente reconocida por (Doughty, 2016[24], Doughty et al., 2018[26]) para Prosthennops serus (Cope, 1878). Esta relativa “pequeña talla” de los ejemplares costarricenses podría obedecer a factores ecológicos limitantes, como lo sería la extensión y el desarrollo paleogeográfico del arco de islas Costa Rica – Panamá en el Mioceno Superior.
Los hallazgos de Prosthennops serus (Cope, 1878) en América Central Meridional, amplía el rango de distribución paleogeográfica de la especie, desde Oregon y Montana en el noroeste hasta Los Apalaches en el centro sureste, las Grandes Planicies, la Costa del Golfo y La Florida en el sureste de E.E.U.U, hasta Honduras y ahora, el sur de Costa Rica. Además, la localidad de San Gerardo de Limoncito con una edad confirmada Hh3 constituye el registro más austral de la especie Protherohyus brachydontus Dalquest y Mooser (1980) y el único de América Central.
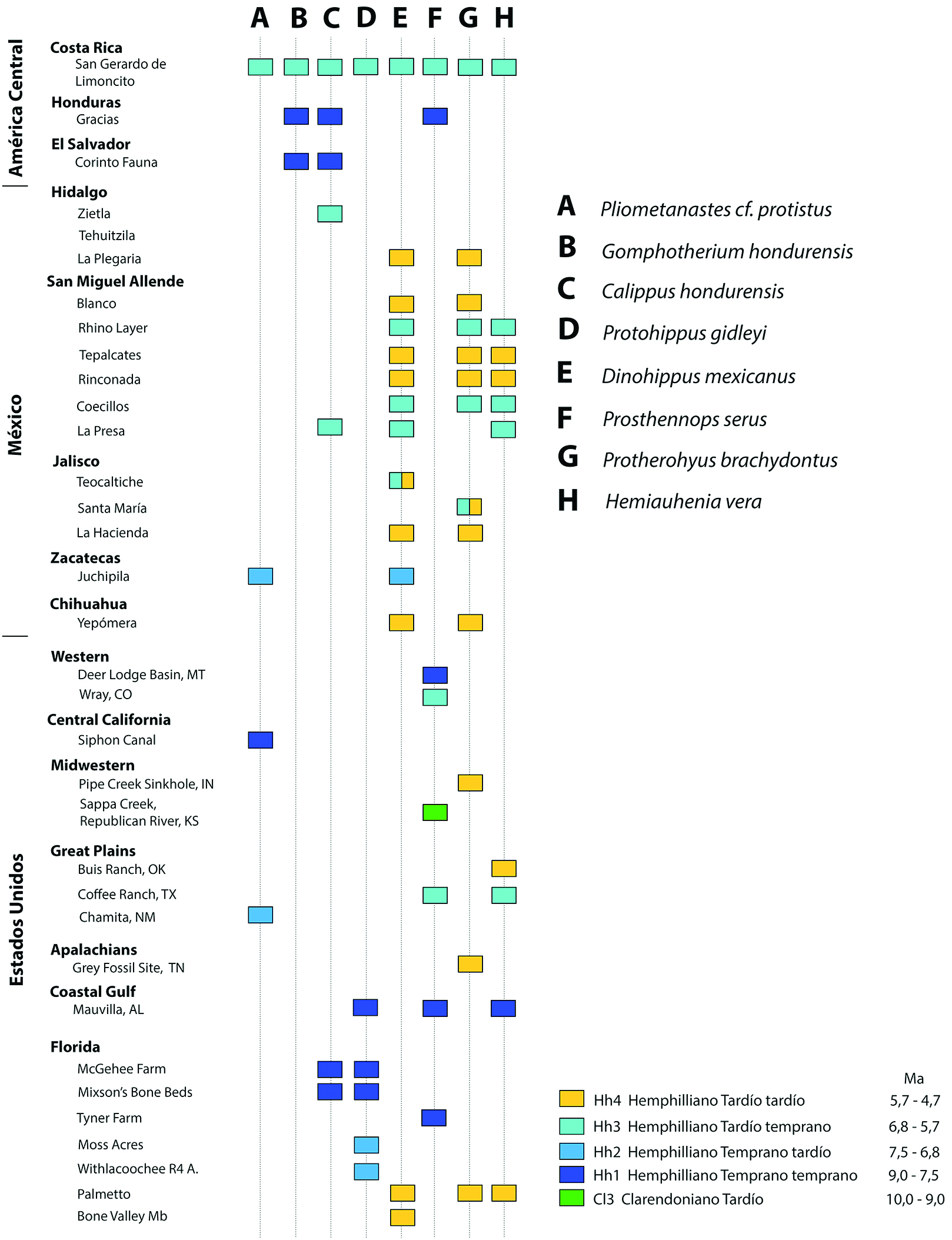
Fig. 11 Cuadro de correlación biocronológica y paleobiogeográfica de los mamíferos de la localidad de San Gerardo de Limoncito con otras localidades fosilíferas de América Central y del Norte.
Las especies Prosthennops serus (Cope, 1878) y Protherohyus brachydontus Dalquest y Mooser (1980)[31] habitaron en un bosque lluvioso tropical mucho más húmedo que los bosques lluviosos actuales del sur de Costa Rica, con presencia abundante de plantas tipo C3 (cf. Pérez Crespo et al., 2018)[48]. Durante el Hemphilliano Tardío temprano Hh3, alrededor de los 5.8 Ma, en esta época al final del Mioceno, estos bosques tropicales se ubicaban al nivel del mar próximos al litoral y estaban asociados a posibles humedales tipo estuario (Valerio, 2010)[49].
Simpatría
El registro de 2 géneros y especies de Tayassuinae en el Hemphilliano tardío Hh3 del sur de América Central, sugiere potencial competición por recursos y territorio, muy similar a lo observado por Doughty, 2016[24] para la fauna de peccaries de Gray Fossil Site del Mioceno Superior de Tennessee.
¿Cómo se dio esta competencia? Pues no es posible determinarlo, los escenarios posibles serían: 1. Compartición del territorio debido a una diferencia en el consumo de frutos y otros alimentos, 2. Comportamiento evasivo de las dos especies y 3. División temporal del hábitat, estos dos últimos casos se darían mediante el uso de glándulas odoríferas y la demarcación de territorios. Todas estas conductas, han sido observadas en regiones donde en la actualidad habitan 2 de las 3, o las 3 especies de pecaries actuales (Bodmer, 1989[50]; Kiltie, 1981[51]; Mayer y Brandt, 1982[52]).
Por último, la localidad de San Gerardo de Limoncito, provee evidencia sólida de la coexistencia de las especies Prosthennops serus (Cope, 1878) y Protherohyus brachydontus Dalquest y Mooser (1980)[31] en el Hemphilliano Tardío temprano (Hh3), algo que Wright, 1998[5] había supuesto, cuando consideró la posibilidad del traslape territorial de ambas especies durante el Hemphilliano Tardío de América del Norte, pero no pudo demostrarlo de manera explícita.

