











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroduction
G.G. Simpson was probably the most influential paleontologist of the XX century. His detailed work and analysis on Cenozoic land mammals was a benchmark to South American paleontologists. Based on his influential work, South America was long considered as an island continent isolated from other landmasses by most of the Cenozoic (approximately 50 millon years). When the Panamá Isthmus arose out of the water some 3 millon years ago, a mass biotic interchange between North and South America started (the Great American Biotic Interchange: GABI; Simpson, 1980; Fig. 1).
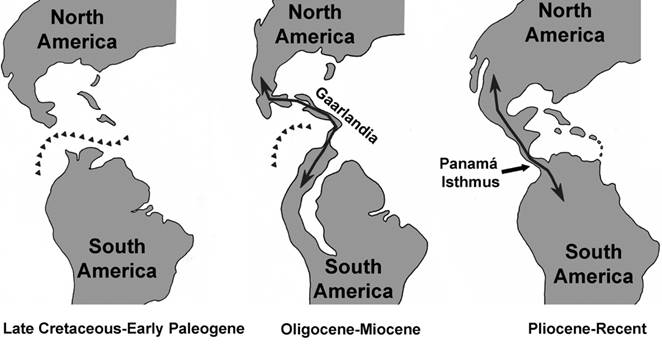
Fig. 1 An alternative model showing early biotic interchange between the Americas. At left, during Latest Cretaceous-early Paleogene North and South America were separated each other. At the middle, Oligocene-Miocene interchange between North America and South America through GAARlandia (GAARBI). At right, post-Pliocene interchange between the Americas through the Panama Isthmus (GABI). Line of triangles indicate presence of deep-sea trough. Reconstructions based on Iturrald-Vinent and Mac-Phee (1999) with modifications.
This biogeographic scheme was followed by most authors until recently (see Patterson and Pascual, 1972; Woodburne et al., 2006; Woodburne, 2010). New evidence coming from geology, paleontology, and molecular biology suggests that the faunistic interchange was older than previously thought and that it was well under progress by the Early Miocene, at least (Farris et al., 2011; Montes et al., 2012, 2015; Carrillo et al., 2015; Bacon et al., 2015; Erkens, 2015; Hoorn and Flantua, 2015; Winston et al., 2016). On this basis, several authors agreed that the splendid isolation of South America should be analyzed in depth (Erkens, 2015) and that migration of biota between this continent and North America should be rethought (Willis and Davis, 2015). Based on biological evidence, Bacon et al. (2015) indicate that the interchange between North and South America faunas was a long and complex process that began as early as the Oligocene-Miocene transition, and probably comprised several distinct migrational events. Further, Winston et al. (2016) cautioned that the GABI was far more complex and that we should depart from the simplistic view of a single migratory event.
This new view of GABI was criticized, among other flaws, because of the lack of empirical geological data sustaining such an old connection between North and South America (Coates and Stallard, 2013; O`Dea et al., 2016). The geology of the Caribbean region is notably complex, involving the Caribbean, Pacific, South American, and North American tectonic plates, and thus, geological data are often contradictory or difficult to analyze.
In spite to the lack of geological data sustaining early connection between the Americas, the paleontological and neontological evidence of a Miocene biotic interchange is strong. Thus, the inconsistency between biological and geological data remains unresolved. The aim of the present contribution is to propose a new hypothesis that may explain the occurrence of Early and Middle Miocene migrants in North and South America.
Discussion
Summary of evidence indicating pre-Pliocene biotic interchange between the Americas
The GABI was a dynamic interchange, including bidirectional migration (Carlini et al., 2008; Castro et al., 2014). The core of the GABI was composed of a series of major migratory waves that begin since 3 Ma. and persisted throughout all Pliocene and Pleistocene ages (Marshall et al., 1979; Webb, 2006; Woodburne et al., 2006; Woodburne, 2010; Cione et al., 2015). The explanation for such faunistic interchange is the complete closure of the Isthmus of Panamá by that time (Simpson, 1980). Recent evidence has prompted some authors to assume that although the bulk of the interchange happened since the Pliocene, this migratory event was probably more gradual than traditionally postulated (Erkens, 2015).
Paleontologists found that a large array of “heralds” such as camels, peccaries, horses, beardogs, monkeys, cricetids, procyonids, mustelids, ground sloths, colubroid and booid snakes, turtles and terror birds passed the landbridge between the continents well before 3 mya (Rusconi, 1933; Hirschfield and Webb, 1968; Webb, 1989; Marshall et al. 1979; Linares 1981; Baskin 1982; Albino and Montalvo, 2006; Czaplewski et al., 2003; Soibelzon and Prevosti 2007, 2012; Morgan, 2008; Verzi and Montalvo 2008; MacFadden et al., 2007; Woodburne 2010; Head et al., 2012; Prevosti and Soibelzon 2012; Laurito and Valerio, 2012; Leigh et al., 2014; Barbiere et al., 2016; O´Dea et al., 2016; Bloch et al., 2016). Most of these heralds were reported by early and middle Pliocene times, namely camelids, equids, mustelids, canids, and possibly felids (see Cione et al., 2015), whereas most taxa arrived in a high migratory pulse since the earliest Pleistocene (e.g., mephitids, ursids, cervids, leporids and man, among others; Cione et al., 2015). The evidence of these late stages of the interchange is abundant, and consensus about the occurrence of this event has been reached by most authors (see details in Cione et al., 2015; O´Dea et al., 2016).
On the other hand, Middle to early Late Miocene taxa that indicate unambiguous faunistic interchange between both Americas include dromomerycine artiodactyls, gomphotheres, peccaries, tapirs, sigmodontine rodents, procyonids, ground sloths, armadillos, bats, boid and colubroid snakes, trionychid turtles and crocodiles (Carranza and Miller, 2004; MacFadden et al., 2007; Vélez-Juarbe et al., 2007; Morgan, 2008; Campbell et al., 2000; 2010; Head et al., 2012; Laurito and Valerio, 2012; Hastings et al. 2013; Fig. 2). Recently, North American migrant mammals were recovered in Late Miocene (or even earlier) strata from the Amazonian basin. These include dromomerycine artiodactyls, gomphotheres, peccaries and tapirs (Campbell et al., 2000; 2010; Frailey and Campbell, 2012; Prothero et al., 2014; but see Lucas, 2013; Antoine et al., 2016). This fauna was dated as being 9.5 Ma. old, although it may be considerably older (Campbell, 2001). It is worthy to mention that Simpson (1980) long ago recognized that during the Late Miocene, procyonids arrived from North America, being the first evidence of some kind of biotic interchange between both landmasses.
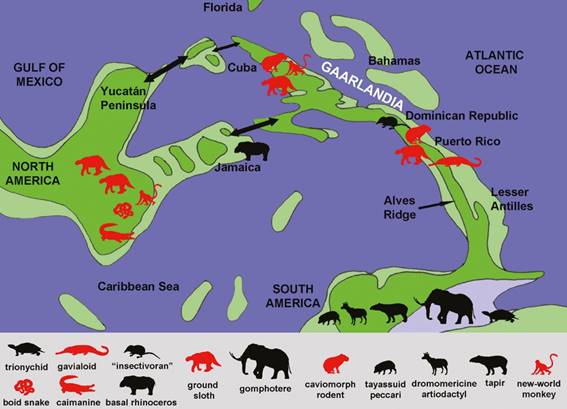
Fig. 2: Palaeobiogeographical map of Central America and Caribbean region by Paleogene (Eocene-Oligocene) times pointing out the occurrence of vertebrates indicating pre-Pliocene faunistic interchange between the Americas. Relevant fossil occurrences and localities are indicated. Double arrows indicate the probably alternative passages of faunas. South American lineages that formed part of the interchange are in red-color, whereas North American taxa are in black. Palaeobiogeographical reconstruction is based on Iturralde-Vinent and MacPhee (1999).
Further, non-paleontological evidence also indicates that migratory events between both landmasses occurred at least since the Early Miocene (23 mya) as suggested by molecular data on plants (Pennington and Dick, 2004; Erkens et al., 2007; Bacon et al., 2013; Willis et al., 2014), insects (Winston et al., 2016) and vertebrates (Koepfli et al., 2007; Weir et al. 2009; Eizirik, 2012).
As follows we attempt to explain these incongruent data on the basis of a new biogeographical hypothesis.
Evidence indicating the existence of a land-bridge connecting North and South America by the Early-Middle Cenozoic
Since the XIX century, biologists proposed several different biogeographical hypotheses with the aim to explain the uniqueness and diversity of Antillean terrestrial vertebrate faunas (Wallace, 1881; Iturralde-Vinent and MacPhee, 1999; Woods et al., 2001). The traditional dispersalist theory proposes that most of the West Indian terrestrial biota arrived by flying or floating, crossing to these islands via a series of apparently uncoordinated dispersal events at various times during the Cenozoic (see Hedges et al., 1992; Hedges, 1996, 2006; see also Ricklefs and Bermingham, 2008). However, a new paleobiogeographical hypothesis is gaining acceptation due to novel evidence coming from paleontology and neontology. Iturralde-Vinent and MacPhee (1999) based on geological evidence find that the Aves Rise was an above water land span between the Greater Antilles and South America by the Late Eocene-Oligocene (Figs. 1-3). This continental bridge was termed GAARlandia by those authors (Greater Antilles+Aves Rise landspan), and was sustained on the basis of abundant geological data (Haq et al., 1987; Miller et al., 1996; Iturralde-Vinent and MacPhee, 1999; MacPhee et al., 2000; Iturralde-Vinent, 2006). The Aves Rise, presently almost completely submerged, was continuous with the Greater Antilles Ridge and other small islands and shallow banks, and have constituted a single entity by the latest Eocene/Early Oligocene times (constituting the GAARlandia landspan). This resulted in most terrestrial vertebrates populating the Antilles across this landspan, as a result of a single migratory event from South America. Iturralde-Vinent and MacPhee (1999) indicate that effects of disruption, combined with marine transgression served to isolate these terraines, by the Early-Middle Miocene. Further, especially high sea-level stands in the Late Miocene, Pliocene and Quaternary submerged this landmass.
Despite the fact that Iturralde-Vinent and MacPhee (1999; see also Iturralde-Vinent, 2006) proposed GAARlandia to be of fleeting existence, restricted to the Eocene-Oligocene boundary (aprox. 35-33 Ma), there is some evidence indicating that this landspan may have persisted at least until the Early Miocene (Fox et al., 1971; Holcombe and Moore, 1977; Donnelly, 1989; Holcombe and Edgar, 1990; Speed et al. 1993). There is evidence of uplift of ridges and islands (Iturralde-Vinent, 2006), existence of shallow-water marine, with terrestrial-derived clastic material (MacPhee et al., 2003), and presence of small islands, sandy keys, and shallows that may be completely subaerial for a short period of time (Iturralde-Vinent, 2006; Fig. 3). Further, a possible dryland connection was also proposed for Miocene times (Duque-Caro, 1990). All this evidence indicates that terrestrial areas were still widespread throughout GAARlandia by the Early Miocene (MacPhee et al. 2003). Further, Bacon et al. (2015) presented extensive evidence indicating that the separation between marine organisms in the Atlantic and Pacific oceans occurred between 23 and 9 Ma., suggesting that a landspan separated both oceans by that time. It appears to be clear that the Aves Rise was flooded by sea level and was deeply submerged by the Late Miocene (Buysse et al., 1985).
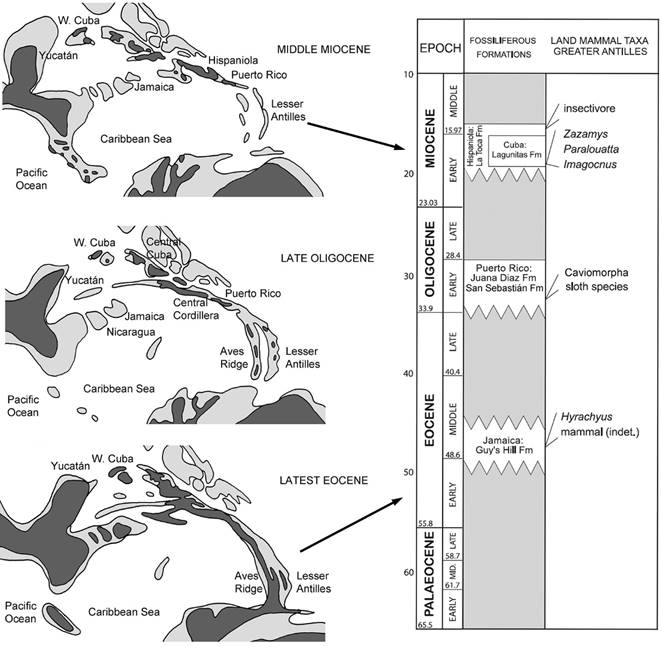
Fig. 3: Paleobiogeographic schematic reconstruction of Caribbean and Central American regions showing main stages discussed in the text (modified from Iturralde-Vinent, 2006). Light grey indicates shallow water, dark grey indicates emerged land. At right, the table shows correlation chart between Cenozoic intervals, and fossiliferous stratigraphic units and Tertiary mammalian records from Greater Antilles (modified from MacPhee, 2005).
As indicated above, the most important implication of the GAARlandia hypothesis is that South American terrestrial taxa dispersed up the landspan into the future Greater Antilles before subsidence occurred (MacPhee et al., 2003). In fact, fossil evidence indicates that for a time period overland dispersal from northern South America into GAARlandia occurred for various elements of the land biota (e.g., caviomorph rodents, ground sloths, monkeys; Borhidi, 1985; MacPhee and Iturralde-Vinent, 1994, 1995, 2000, 2005; Iturralde-Vinent and MacPhee, 1996, 1999; Iturralde-Vinent, 2006; Fig. 2). Some authors sustain that the elements of this biota were of clear South American signature (Borhidi, 1985; MacPhee and Iturralde-Vinent, 1994, 1995, 2000, 2005; Iturralde-Vinent and MacPhee, 1996, 1999; Iturralde-Vinent, 2006; White and MacPhee, 2001). In spite to such claims, several fossils indicate that this pattern was far more complex than thought. For example, Cenozoic arthropods and plants are of distinctly North American aspect, and contain unexpected taxa that are not part of the modern Neotropical fauna and flora (Wilson, 1988; Graham, 1990; MacPhee and Iturralde-Vinent, 1995; Poinar, 2010). The abundant arthropods coming from Miocene Dominican Amber indicate a mixed composition, having strong South American influence, but also North American components (Poinar, 2010). The North American origin of some bats and insectivorans (Dávalos, 2004), and fossils of Eocene rhinocerotans and Miocene insectivorans (MacPhee and Grimaldi, 1996; Domning et al., 1997; Portell et al., 2001) also indicate that the interchange was not uniquely between the Antilles and South America, but included, at least partially, a North American influence (Fig. 2). It appears to be evident that North American faunistic influence in the old Antillean faunas is weaker than the South American influence. It is not improbable that some kind of filter or biotic or geographical barrier existed between North America and the Antilles. Alternatively, this differential influence may be explained by the still notably patchy fossil record of the Antilles.
Paleontological evidence of interchange between the Caribbean Islands and South and North America is still patchy, mainly due to the paucity of the Tertiary fossil record of the Antilles. In spite to that, a large number of mammalian clades were reported from Late Oligocene to Early Miocene beds of several islands. Among these clades are insectivorans (MacPhee and Grimaldi, 1996), three ground sloth clades (MacPhee and Iturralde-Vinent, 1994; MacPhee and Iturralde-Vinent, 2000; MacPhee et al., 2000; White and MacPhee, 2001; Dávalos, 2004), caviomorph rodents (Woods et al., 2001; MacPhee et al., 2003; MacPhee, 2005; Dávalos, 2004), bats (Dávalos, 2004), and primates (Horovitz and MacPhee, 1999; MacPhee et al. 2003; Dávalos, 2004). Insectivores, monkeys, and sloths appeared in the Antilles by the Early Miocene (aproximately 20 Ma; Ali, 2012), whereas rodents have their first record by Oligocene times (Vélez-Juarbe et al. 2014)(Fig. 3).
Molecular evidence sustains that several clades of frogs (Crawford and Smith, 2005; Moen and Wiens, 2009; Alonso et al., 2012), freshwater fishes (Murphy et al., 1999; Perdices et al., 2005; Concheiro Pérez et al., 2007; Hrbek et al., 2007; Hulsey et al., 2011; Rican et al., 2013), spiders (Crews and Gillespie, 2006; Binford et al., 2008), and plants (Fritsch, 2003; Pennington and Dick, 2004; van Ee et al., 2008), may have also dispersed through GAARlandia. Further, detailed analyses by Dávalos (2004) agree in that most non-volant Caribbean mammalian lineages arrived in the West Indies before the Middle Miocene. Further, some authors suggested that the existence of GAARlandia may have resulted in some kind of biotic interchange (Pennington and Dick, 2004; Rican et al., 2013; Hastings et al. 2013; see also Lucas, 1986).
There is some evidence that argues against a biotic interchange between North and South America through GAARlandia. It should be argued that because mastodonts and trionychids are present in Miocene beds of Central America, but are absent in the Antillean fossil record, the entrance of these taxa may have occurred through Central America, rather than by GAARlandia (see Montellano-Ballesteros, 2002; Lucas and Alvarado, 2010; Laurito and Valerio, 2010). However, tryonichid remains are so incompletely preserved that comparissons between South and Central America fossils are not conclusive regarding the phylogenetic ties of available elements (see Laurito et al., 2005). On the other hand, fossil proboscideans from the Miocene of Central America are not phylogenetically close to South American forms (Mothé et al. 2012, 2017), and this fact weakens the possibility that mastodonts that arrived from South America were descendant of these Central American forms.
The GAARlandia model indicates that during the latest Eocene-Oligocene/early Miocene the islands of the Greater Antillean Ridge were in a close-packed array, they either constituted a single large island or a series of islands separated by very narrow water gaps. Thus, dispersal between the Antilles and the mainland may have been easier during the Oligocene and Miocene that it would appear today (Iturralde-Vinent and MacPhee, 1999; Crawford and Smith, 2005; Hastings et al., 2013; Rican et al., 2013).
Based on evidence at hand, we think that the presence of such islands or a narrow landspan between the Americas (GAARlandia) during the Mid-Cenozoic may explain the early dispersal of some taxa between North and South America, as well as the arrival of mammals from both landmasses to the Caribbean islands.
Why the migrants are so poorly represented in the early fossil record
O`Dea et al. (2016) criticized Montes et al. (2015) and Bacon et al. (2015) hypothesis on an early biotic interchange, based on several grounds, including that this interchange begin by Miocene times, why the migrants were so underrepresented in the record of North and South America? In this regard, Morgan (2008) indicates that the Miocene phase of the interchange between Americas hardly qualifies as a major faunal interchange, involving just a few genera. Further, the Antillean Islands apparently lacked representatives of many South American groups that should have made the journey (Simpson, 1956).
Montes et al. (2015; see also Jaramillo et al., 2017) indicate that this scarcity is probably due to the presence of strong ecological barriers (see also Iturralde-Vinent and MacPhee, 1999; MacPhee, 2005). Further, it is possible that sampling bias toward higher latitudes and the still scarce data from tropical faunas may be responsible for such a pattern, as pointed out by Carrillo et al. (2015). In this regard, Campbell et al. (2010, 2011) hypothesized that the faunistic interchange during the Miocene between North and South America was more extensive than previously realized, but is underrepresented because most taxa involved were tropical forest species that did not reach the temperate regions of North or South America, where the fossil record is relatively complete.
We agree with Iturralde-Vinent and MacPhee (1999) that the mechanism that permitted this limited interchange is still obscure and far from being well understood. A full understanding of this interchange will require better knowledge of the GAARlandia landbridge and its environments. Data are also needed on the existence and duration of intermittent transoceanic connections elsewhere.
Conclusions
When and under what conditions North and South American landmasses as well as the Caribbean Islands interchange land-vertebrates continues to be a provoking question.
We here propose that vertebrate interchange between both Americas maybe separated into two different stages, at least: 1) a Oligocene-Miocene GAARlandia Interchange (GAARI) and the 2) Plio-Pleistocene Great American Biotic Interchange (GABI) (Fig. 1).
Evidence at hand indicates that the first migratory episode (GAARI) may have played a similar role to the GABI, but into a narrow time window and involving a lesser number of taxa. The aim of the present contribution is to propose an alternative hypothesis that may explain the anomalous occurrence of Early and Middle Miocene migrants in North and South America. Nevertheless, as sustained by O´Dea et al. (2016), the evidence of such a wholesale interchange is still scarce, and we think that is far from being well understood.
More studies using a proper methodological framework and new fossils are urgently needed to deconstruct the proportion of immigrant taxa, with the GAARlandia hypothesis as opposed to overwater dispersal with a random time distribution. Further, as expressed by MacPhee (2005), the anomalous appearances of taxa of the sort discussed in this paper should be viewed as the key to lead to new ideas, and they perform an invaluable service to historical biology.

