











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
En el estado de Sonora se encuentran afloramientos de rocas sedimentarias del Carbonífero (Misisípico-Pensilvánico) con una abundante y diversa biota conformada por algas, foraminíferos, esponjas, corales, briozoos, braquiópodos, moluscos y crinoideos. Particularmente, con relación a la Sierra Santa Teresa, uno de los primeros estudios fue realizado por King (1939) quien cartografió la geología de la parte norte y centro de Sonora y asignó a las calizas de esta localidad una edad del Pérmico. Peiffer-Rangin (1988) estudió la bioestratigrafía del Paleozoico de varias regiones de Sonora, entre ellas, el sector norte de la Sierra Santa Teresa y citó la presencia de foraminíferos, esponjas, corales y placas articulares de crinoideos del Pensilvánico-Pérmico correspondientes a depósitos de plataforma.
En los trabajos de Stewart y Amaya-Martínez (1993) y Stewart et al. (1997) se dio a conocer la estratigrafía de las rocas sedimentarias con edades del Pensilvánico al Jurásico de la Sierra Santa Teresa y particularmente sobre el Paleozoico superior reportaron la existencia de invertebrados entre ellos, foraminíferos fusulínidos y conodontos. Posteriormente, Cuen-Romero et al. (2016) citaron la presencia de invertebrados del Paleozoico procedentes de diversas localidades de Sonora. Navas-Parejo (2018) realizó la revisión de la bioestratigrafía del Carbonífero de Sonora y Buitrón-Sánchez et al. (2023) publicaron sobre la sistemática de los crinoideos del Carbonífero procedentes de la Sierra Santa Teresa. Estos fósiles tuvieron una distribución cosmopolita en los bordes de plataforma del Océano Reico que servía de enlace entre los océanos de Panthalassa al oeste de Norte América y el Paleo-Tethys al este de Eurasia (Buitrón-Sánchez et al., 2022). La información presentada de los registros nuevos de invertebrados de esta localidad contribuye al conocimiento de la bioestratigrafía, paleoecología y paleogeografía del Paleozoico superior de México.
El objetivo principal de este trabajo es dar a conocer la composición biótica de la Sierra Santa Teresa, contribuir al conocimiento de las características ambientales donde se desarrollaron y su distribución geográfica en el Carbonífero de diferentes regiones del mundo.
Materiales y métodos
Localización geográfica: La Sierra Santa Teresa se localiza a 20 Km al sureste de la ciudad de Hermosillo, Municipio de Bácum, Valle del Yaqui en el estado de Sonora, con coordenadas geográficas 28° 58' 56” N y 110° 51’ 27” W del meridiano de Greenwich (Fig. 1, Fig. 2).
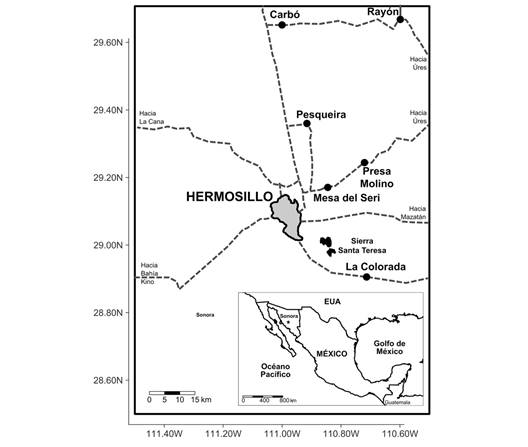
Fig. 1 Mapa de localización de la Sierra Santa Teresa, estado de Sonora, México. / Fig. 1. Location map of the Sierra Santa Teresa, Sonora State, Mexico.

Fig. 2 Vista panorámica de la Sierra Santa Teresa, estado de Sonora, México. / Fig. 2. Panoramic view of the Sierra Santa Teresa, Sonora State, Mexico.
La información geológico-paleontológica de la Sierra Santa Teresa se obtuvo con la consulta de las cartas geológico-mineras y topográficas H12-D41 Hermosillo, H12-D51, Estación Torres/Estación Serdán (Servicio Geológico Méxicano, 2011a; Servicio Geológico Méxicano, 2011b) escala 1:50.000, con artículos de revistas especializadas y tesis. Particularmente, se consultaron los artículos de Peiffer-Rangin (1988), Stewart y Amaya-Martínez (1993), Stewart et al. (1997) y Stewart et al. (1999). La prospección geológico-paleontológica de la secuencia de calizas del Misisípico-Pensilvánico de la Sierra Santa Teresa, se realizó en dos ocasiones en 2021 y 2023. Los fósiles se recolectaron con control estratigráfico y en el Departamento de Geología de la Universidad de Sonora (UNISON) se realizó la preparación de las muestras y la obtención de imágenes fotográficas; en el Instituto de Geología de la Universidad Nacional Autónoma de México (UNAM) se continuó con el trabajo de laboratorio y gabinete. Para el estudio de los invertebrados se utilizó la información de los trabajos de Ausich (1998); Ausich et al. (1999); Ausich et al. (2022); Bassler (1953); Buitrón-Sánchez et al. (2023); Condra y Elias (1944); Hill (1967); Jeffords y Miller (1968); McGhee y McKinney (2002); McKinney y Jackson (1989); Moore y Jeffords (1968); Muir-Wood y Cooper (1960); Rodríguez-García (1984); Stanton et al. (2016); Stukalina (1966); Stukalina (1988). Se asignaron a los ejemplares fósiles números provisionales del Catálogo USONDG-240-256, de la Colección Paleontológica del Departamento de Geología de la Universidad de Sonora.
Marco geológico y contenido fósil de la Sierra Santa Teresa: En la Sierra Santa Teresa se encuentra expuesta una de las secuencias sedimentarias del Paleozoico superior, representadas principalmente por calizas de mares someros. Peiffer-Rangin (1988) midió más de 1 600 m en secciones estratigráficas de caliza y citó la existencia de una abundante y diversa biota del Paleozoico superior. Stewart et al. (1997) publicaron sobre una de las secuencias sedimentarias más completas con edades del Misisípico al Pérmico en la Sierra Santa Teresa, constituida principalmente por caliza, recientemente la edad ha sido confirmada por Buitrón-Sánchez et al. (2023). A continuación, se describen las siete unidades informales propuestas por Stewart et al. (1997) que se complementaron con nuevos registros de invertebrados (Fig. 3, Fig. 4, Fig. 5). Varios de los invertebrados encontrados fueron descritos por Moore & Jeffords, 1968.
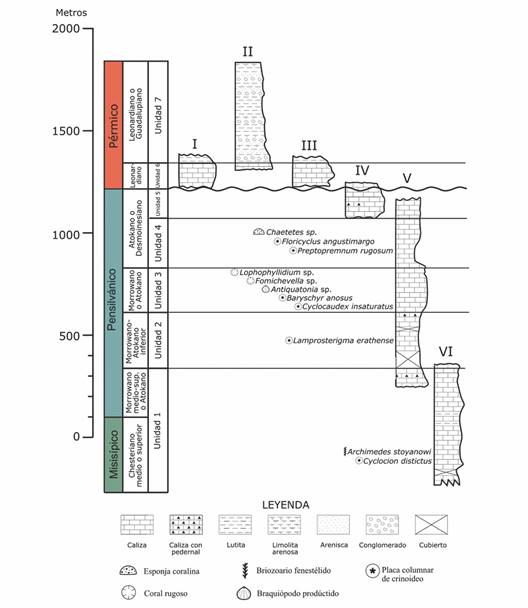
Fig. 3 Representación de las unidades estratigráficas reportadas en Sierra Santa Teresa (Modificada de Stewart et al., 1997). Las unidades 1-6 se desarrollaron en un ambiente marino somero; la unidad 7 corresponde a zonas profundas. Se complementa con nuevos registros de especies de invertebrados (Buitrón-Sánchez et al., 2023). / Fig. 3. Representation of the stratigraphic units reported in Sierra Santa Teresa (Modified from Stewart et al., 1997). Units 1-6 were developed in a shallow marine environment; Unit 7 corresponds to deep areas. Complemented with new records of invertebrate species (Buitrón-Sánchez et al., 2023).

Fig. 4 A-B. Chaetetes sp., ejemplar USONDG-234, A. vista de la superficie de la colonia y B. corte transversal; C. Lophophyllidium sp., vista superior, ejemplar USONDG-235; D-E. Fomichevella sp., ejemplar USONDG-236 D. vista lateral del coral, E. vista superior; F. Archimedes stoyanowi, incrustado en roca caliza, ejemplar USONDG-238; G-H. Antiquatonia sp., valva dorsal, ejemplar USONDG-239. G. vista externa, H. vista interna. / Fig. 4. A-B. Chaetetes sp., specimen USONDG-234, A. view of the colony surface and B. cross section; C. Lophophyllidium sp., top view, specimen USONDG-235; D-E. Fomichevella sp., specimen USONDG-236 D. lateral view of the coral, E. top view; F. Archimedes stoyanowi, embedded in limestone rock, specimen USONDG-238; G-H. Antiquatonia sp., dorsal valve, specimen USONDG-239. G. external view, H. internal view.

Fig. 5 A. Baryschyr anosus, vista de la placa articular, ejemplar USONDG-240; B-D. Cyclocaudex insaturatusMoore & Jeffords, 1968, B. vista de la placa articular con placas cirrales, C. noditaxis con marcas cirrales, ejemplares USONDG-241 y USONDG-242; E-F. Floricyclus angustimargoMoore & Jeffords, 1968, ejemplar USONDG-244. E. vista de la placa articular, F. noditaxis; G-H. Cyclocion distictusMoore & Jeffords, 1968, ejemplar USONDG-248. G. vista de la placa articular, H. noditaxis; I-J. Lamprosterigma erathenseMoore & Jeffords, 1968, ejemplares USONDG-250 y USONDG-251. I. vista de la placa articular. J. noditaxis con un briozoario adherido, K-L. Preptopremnum rugosumMoore & Jeffords, 1968, ejemplar USONDG-255. K. vista de la placa articular, L. noditaxis. / Fig. 5. A. Baryschyr anosus, view of the articular plate, specimen USONDG-240; B-D. Cyclocaudex insaturatusMoore & Jeffords, 1968, B. view of articular plate with cirral plates, C. noditaxis with cirral markings, specimens USONDG-241 and USONDG-242; E-F. Floricyclus angustimargoMoore & Jeffords, 1968, specimen USONDG-244. E. view of the articular plate, F. noditaxis; G-H. Cyclocion distictusMoore & Jeffords, 1968, specimen USONDG-248. G. view of the articular plate, H. noditaxis; I-J. Lamprosterigma erathenseMoore & Jeffords, 1968, specimens USONDG-250 and USONDG-251. I. view of the articular plate. J. noditaxis with an attached bryozoan, K-L. Preptopremnum rugosumMoore & Jeffords, 1968, specimen USONDG-255. K. view of the articular plate, L. noditaxis.
Unidad 1: Es la unidad más antigua expuesta en Sierra Santa Teresa, conformada por estratos delgados de caliza bioclástica con textura mudstone, wackestone y packstone con placas articulares de crinoideos de la morfoespecie Cyclocium distintus Moore & Jeffords. El espesor de la unidad se estima en 579 m. El análisis de conodontos y la presencia de briozoos fenestélidos de la especie Archimedes stoyanowiCondra & Elias (1944) indica que la edad de la base de esta unidad es del Misisípico Tardío (Chesteriano), mientras que la parte superior corresponde al Pensilvánico Medio.
Unidad 2: La unidad tiene aproximadamente 176 m de espesor, la cima está conformada por caliza con textura mudstone y wackestone, con fósiles de corales, braquiópodos y escasas placas columnares de crinoideos de la especie Lamprosterigma erathense Moore & Jeffords. La edad corresponde al Pensilvánico (Morrowano tempano-Atokano temprano).
Unidad 3: La unidad está conformada por caliza con textura mudstone y wackestone recristalizada, con un espesor total de 230 m, con pedernal en algunas zonas y escasas placas columnares de crinoideos. Buitrón-Sánchez et al. (2023) identificaron para esta unidad fósiles de corales rugosos (Lophophyllidium sp., Fomichevella sp.), braquiópodos (Antiquatonia sp) y crinoideos (Baryschyr anosus Moore & Jeffords, Cyclocaudex insaturatus Moore & Jeffords). La edad con base en el estudio de los conodontos fue propuesta por Stewart et al. (1997) para el Pensilvánico Temprano-Medio (Morrowano-Atokano) y complementada con el estudio de crinoideos (Buitrón-Sánchez et al., 2023).
Unidad 4: La unidad tiene 261 m de espesor y está conformada por caliza bioclástica con textura mudstone y wackestone (Stewart et al., 1997). En la parte media de esta unidad se recolectaron esponjas coralinas del género Chaetetes, placas columnares de crinoideos de las morfoespecies Floricyclus angustimargo Moore & Jeffords y Preptopremnum rugosum Moore & Jeffords (Buitrón-Sánchez et al., 2023) y fusulínidos que determinaron la edad para el Pensilvánico Medio (Atokano).
Unidad 5: La unidad está constituida por caliza con textura mudstone con algunos lentes y nódulos pedernal, contiene escasos estratos de caliza con textura wackestone y presenta un espesor aproximado de 164 m. El estudio de los conodontos y fusulínidos indican una edad del Atokano al Desmoinesiano (Pérez-Ramos, 2001; Stewart et al., 1997).
Unidad 6: La unidad tiene 122 m de espesor y está conformada por caliza bioclástica de textura packstone con placas columnares de crinoideos y foraminíferos fusulínidos; también contiene lentes y nódulos de pedernal con horizontes de caliza recristalizada. La edad determinada por los fusulínidos es del Pérmico temprano (Leonardiano). Las unidades 1 a 6 corresponden a sedimentos marinos de agua somera (Stewart et al., 1997).
Unidad 7: Esta unidad corresponde a estratos de caliza limosa-arenosa, lodolita limosa y arenisca limosa con rizaduras o laminación convoluta. Su espesor se ha estimado en 610 m. En la unidad existen facies de calcarenita masiva con Nereites y fusulínidos. El ambiente corresponde a depósitos de agua profunda. La edad se ha establecido a partir de estudios de fusulínidos y se considera del Pérmico temprano (Leonardiano o Guadalupiano) (Stewart et al., 1997).
Resultados
Las rocas sedimentarias de la Sierra Santa Teresa contienen una abundante y diversa biota conformada por algas, foraminíferos, esponjas, corales, briozoos, braquiópodos, moluscos, crinoideos y conodontos (Buitrón-Sánchez et al., 2023; Peiffer-Rangin, 1988; Stewart et al., 1997). Esta biota presenta un intervalo estratigráfico del Misisípico Medio-Superior (Chesteriano) al Pensilvánico Medio (Desmoinesiano). Particularmente, se estudiaron fósiles de briozoos fenestélidos de la especie A. stoyanowi y crinoideos de las especies C. distinctus del Misisípico Tardío (Chesteriano medio-superior) que fueron recolectados en la base de la secuencia estratigráfica de la sierra. En la unidad 2, en niveles de caliza con pedernal, se obtuvieron numerosas placas articulares del crinoideo de la especie L. erathense con una edad relativa correspondiente al Pensilvánico Temprano (Morrowano-Atokano). En la unidad 3, en niveles de caliza con textura mudstone y wackestone se identificaron fósiles de corales rugosos (Lophophyllidium sp., Fomichevella sp.), braquiópodos (Antiquatonia sp.) y placas articulares de crinoideos (Baryschyr anosus Moore & Jeffords, Cyclocaudex insaturatus Moore & Jeffords); esta asociación faunística presenta una edad del Pensilvánico Temprano (Morrowano-Atokano). En la unidad 4, conformada por caliza con textura mudstone y wackestone, se recolectaron esponjas coralinas del género Chaetetes y placas columnares de crinoideos de las morfoespecies Floricyclus angustimargo y P. rugosum (Fig. 4, Fig. 5).
Discusión
Consideraciones paleoecológicas: Durante el Misisípico Superior (Chesteriano) al Pensilvánico Medio (Morrowano-Desmoinesiano), la asociación biótica de la sierra Santa Teresa se desarrolló en una plataforma carbonatada de mares tropicales y someros ubicada en el margen suroeste del Cratón Norteamericano y el Océano Reico. La biota está principalmente representada por numerosas morfoespecies de crinoideos (Baryschyr anosus Moore & Jeffords, Cyclocaudex insaturatus Moore & Jeffords, Floricyclus angustimargo Moore & Jeffords, Cyclocion distictus Moore & Jeffords, L. erathense, P. rugosum) asociados a esponjas arrecifales (Chaetetes), corales solitarios (Lophophyllidium, Fomichevella), briozoos fenestélidos (A. stoyanowi) y braquiópodos prodúctidos (Antiquatonia).
Los quetétidos (Chaetetes) fueron los principales constructores de montículos arrecifales, localizados en facies de plataforma y áreas epicontinentales de mares someros con escasa sedimentación (Connolly et al., 1989; Gómez-Espinosa et al., 2006; Suchy & West, 2001; West & Kershaw, 1991). Los briozoos del género Archimedes suelen indicar que había movimientos leves en la columna de agua que permitieron su desarrollo, aunque existen formas asociadas a diferentes sustratos y niveles de energía (McGhee & McKinney, 2002). El género Archimedes ha sido considerado entre los briozoos más comunes de los mares someros del Misisípico Tardío (Chesteriano) en el Cratón Norteamericano (McKinney, 1993). McGhee y McKinney (2002) y McKinney y Raup (1989) consideran que la forma de crecimiento en briozoarios de láminas fenestradas poco abiertas se asocian a ambientes de menor profundidad, en comparación con los que tienen las láminas más abiertas están asociadas a zonas más profundas. Los corales solitarios y braquiópodos prodúctidos tuvieron una importante distribución mundial por la conexión marina a través del Océano Reico con los océanos de Panthalassa y Paleo-Tethys. Para el Pensilvánico se pierde la conexión marina generando regionalizaciones específicas o genéricas en estos grupos (Montañez & Poulsen 2013; Wang et al., 2022). Las condiciones climáticas y la libre circulación de corrientes marinas permitieron el incremento de crinoideos que son uno de los invertebrados más representativos de esta sierra. Los crinoideos fueron invertebrados marinos dominantes en los márgenes de plataformas del Carbonífero, estos organismos vivieron fijos al sustrato en asociación con otros organismos epibentónicos (corales, briozoos, braquiópodos, moluscos). Ausich (1978), Ausich (1980) y Kammer y Ausich (2006) consideraron que para comunidades con organismos suspensívoros, existen datos acerca de la relación que guardan con el sustrato (bentos) y se indica que las comunidades con organismos de tamaños menores o iguales a 15 cm habrían presentado mayor competencia por el alimento (Fig. 6).
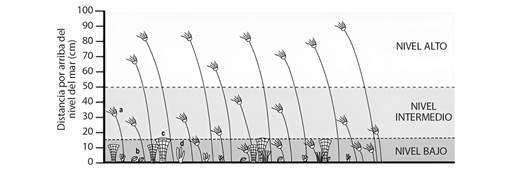
Fig. 6 Representación de comunidades de crinoideos y otros invertebrados bentónicos para el Misisípico temprano a) crinoideos, b) braquiópodos, c) briozoos, d) algas. Cuando la longitud del crinoideo era menor a 15 cm (Nivel Bajo) aumentaba la competencia por el alimento de otros organismos que también se alimentaban por filtración de partículas suspendidas; cuando la longitud era mayor a 15 cm (Niveles Intermedio y Alto), la competencia por el alimento era menor. Modificada de Ausich (1980). / Fig. 6. Representation of communities of crinoids and other benthic invertebrates for the Early Mississippian a) crinoids, b) brachiopods, c) bryozoans, d) algae. When the length of the crinoid was less than 15 cm (Lower Level), competition for food from other organisms that also fed by filtration of suspended particles increased; when the length was greater than 15 cm (Intermediate and Hight levels), competition for food was less. Modified from Ausich (1980).
Asociaciones bióticas como la anterior, son típicas del Carbonífero de plataformas marinas y montículos arrecifales (Newell, 1972; West, 1988). Los crinoideos de la Sierra Santa Teresa fueron los invertebrados más abundantes representados por placas articulares aisladas y fragmentos de las columnas. Las bajas tasas de sedimentación y lento enterramiento pudieron haber permitido que la biota, particularmente, los briozoos, braquiópodos y crinoideos, permanecieran por un tiempo más prolongado a la intemperie previo al enterramiento, favoreciendo que perdieran alguna de sus partes por desarticulación durante el proceso de fosilización (Gómez-Espinosa et al., 2006).
Consideraciones paleogeográficas: En el Misisípico, la mayoría de las masas continentales entre ellas América del Norte, Europa, América del sur, África, Arabia, India, Antártida, y Australia, se encontraban ubicadas en la parte paleoecuatorial y sur del globo terráqueo, en tanto que Mongolia y los bloques de China del norte y sur se localizaban en la parte norecuatorial y Siberia hacia el norte. La cercanía de los continentes y la reducción de barreras geográficas propiciaron el intercambio entre especies tropicales alrededor del Paleoecuador y a través del Océano Reico (Angiolini et al., 2022; Aretz et al., 2020; Lucas et al., 2022). Los briozoos fenestélidos de la especie A. stoyanowi del Chesteriano de la sierra Santa Teresa se han descrito anteriormente para la Formación Paradise en Arizona, Estados Unidos de Norteamérica (Condra & Elias, 1944). El braquiópodo Antiquatonia reportado para el Morrowano-Atokano de la Sierra Santa Teresa, se ha considerado como un género cosmopolita que se encuentra en distintas localidades de la región del Mid-Continent durante el Carbonífero (Torres-Martínez et al., 2018). Sobre los registros de crinoideos descritos para el Carbonífero de México es importante tomar en cuenta que presentan asociaciones con diversos organismos que vivieron en los mares tropicales y someros de distribución tethysiana entre estos, algas incrustantes y filoides (Eugonophyllum, Kamaena, Zidella), foraminíferos fusulínidos (Triticites, Fusulinella) típicos de la región del Mid-Continent que son similares a la biota de la Provincia Euroasiática-Ártica, también asociados a corales (Lophophyllidium, Neozaphrentis, Caninophyllum, Cyathaxinia, Lithostrotionella) citados para la provincia Cordillerana en América del Norte y para la provincia Euroasiática, en asociación con braquiópodos (Avonia, Wellerella, Composita, Hustedia), trilobites (Griffithides) y conodontos (Buitrón-Sánchez et al., 2006; Buitrón-Sánchez et al., 2008).
En el Misisípico y Pensilvánico de México, en varias localidades de Sonora, entre ellas, la Sierra Agua Verde, se han identificado crinoideos de las morfoespecies Cyclocaudex insaturatus, L. erathense y P. rugosum, coincidentes con registros de Sierra Santa Teresa; también se han descrito corales tabulados (Michelinea), corales solitarios (Lophophyllidium), braquiópodos espiriféridos (Spirifer, Anthracospirifer), prodúctidos (Antiquatonia), gasterópodos (Eumphalus) y conodontos (Buitrón-Sánchez, Almazán & Vachard, 2005; Buitrón-Sánchez, Gómez-Espinosa et al., 2007; Buitrón-Sánchez et al., 2015, Gómez-Espinosa et al., 2006; Gómez-Espinosa et al., 2008; Villanueva-Olea et al., 2019). Para Cerros El Tule se han registrado las morfoespecies Cyclocaudex insaturatus y P. rugosum (Buitrón-Sánchez et al., 2008; Buitrón-Sánchez et al., 2012). Para el Pensilvánico de la Sierra Las Mesteñas, Sonora, han sido identificados crinoideos de las morfoespecies Cyclocaudex insaturatus, Floricyclus angustimargo, L. erathense y P. rugosum (Buitrón-Sánchez, Almazán, Vachard, Gómez-Espinosa et al., 2005; Buitrón-Sánchez et al., 2008; Villanueva-Olea & Buitrón-Sánchez, 2013; Villanueva-Olea et al., 2016) las cuales guardan similitud con registros del Misisípico y Pensilvánico del estado de Durango (Formación Gran Tesoro) donde se han identificado Cyclocaudex insaturatus en asociación con P. rugosum, reconocidas del bloque metamórfico con protolito sedimentario del Pensilvánico Medio en asociación con briozoos fenestélidos (Eguiluz-de Antuñano et al., 2014). P. rugosum es una especie en común con la fauna del Wordiano (Pérmico medio) de la Formación Monos en Caborca, Sonora (Buitrón-Sánchez, Almazán et al., 2007). A su vez, Cyclocaudex insaturatus y P. rugosum tienen afinidad con registros del Wordiano-Capitaniano de la Formación Las Delicias en el estado de Coahuila (Villanueva-Olea et al., 2021).
En la región centro-oriente de México, los registros de crinoideos de Sierra Santa Teresa son similares con los del Pensilvánico de la Formación Del Monte en Calnali, Hidalgo, entre ellos Baryschyr anosus, Cylindrocauliscus fiski, Cyclocaudex sp. cf. C. costatus, C. insaturatus, C. jucundus, Mooreanteris waylandensis, Plummeranteris sansaba y Heterosteleschus keithi asociados a algas rodofíceas, briozoos fenestélidos y braquiópodos de las especies Neospirifer sp. cf. N. fasciger y Dictyoclostus sp. (Buitrón-Sánchez et al., 1987).
En la región de San Salvador Patlanoaya, Puebla, se han estudiado una de las más abundantes y diversas especies de crinoideos del Misisípico y Pensilvánico, de esta localidad se citan a Floricyclus angustimargo y P. rugosum, en asociación con Ampholenium apolegma, Cyclocaudex jucundus, Dierocalipter doter, Lomalegnum hormidium, Stiberostaurus aestimatus, Blothronagma cinctutum, Cyclocaudex typicus, Cyclocaudex sp., Cyclosteleschus turritus, Cyclocrista cheneyi, Floricyclus welleri, Mooreanteris perforatus, M. waylandensis, Pentagonomischus plebeius y Pentagonomischus sp. (Esquivel-Macías et al., 2000; Velasco-de León & Buitrón-Sánchez, 1992; Villaseñor-Martínez et al., 1987).
De la región del suroeste de México, se cita a P. rugosum de la Formación Olinalá del Pensilvánico Superior en el estado de Guerrero. En esta formación también se han estudiado las especies de crinoideos Cyclocaudex costatus, Heterostelechus jeffordsi, Pentaridica pentagonalis e Isocrinus sp. (Buitrón-Sánchez, Silva et al., 2005; Esquivel-Macías et al., 2000; Esquivel-Macías et al., 2004; Flores-de Dios & Buitrón-Sánchez, 1982; González-Arreola et al., 1994; Vachard et al., 1993). Para la formación Santiago del Pensilvánico Inferior-Medio del estado de Oaxaca, se describieron las morfoespecies Cyclocion distinctus Pentagonomischus sp. cf. P. plebeius, Lomalegnum sp. cf. L. hormidium, Axilinucrinus angustus?, Graphosterigma sp., Cylindrocauliscus fiski, Cyphostelechus sp. cf. C. claudus y Cyclocaudex typicus. Además, se ha mencionado la presencia de corales solitarios, briozoos, braquiópodos (Neochonetes granulifer, Neospirifer dunbari), pelecípodos, gasterópodos y trilobites (Villanueva-Olea et al., 2011), (Fig. 7).
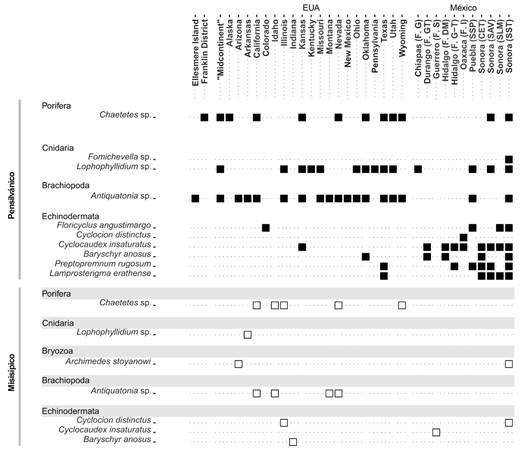
Fig. 7 Distribución de registros de la biota identificada en Sierra Santa Teresa y de otras localidades de México, Estados Unidos de Norteamérica y Canadá. Abreviaturas: F. G: Formación Grupera; F. GT: Formación Gran Tesoro; F. S: Formación Santiago; F. DM: Formación Del Monte; F. G-T: Formaciones Guacamaya-Tuzancoa; F. I: Formación Ixtaltepec; SSP: San Salvador Patlanoaya; CET: Cerros El Tule; SAV: Sierra Agua Verde; SLM: Sierra Las Mesteñas; SST: Sierra Santa Teresa. / Fig. 7. Distribution of records of the biota identified in Sierra Santa Teresa, Sonora, and other Mexico, USA, and Canada localities. Abbreviations: F. G: Grupera Formation; F. GT: Gran Tesoro Formation; F. S: Santiago Formation; F. DM: Del Monte Formation; F. G-T: Guacamaya-Tuzancoa Formations; F. I: Ixtaltepec Formation; SSP: San Salvador Patlanoaya; CET: Cerros El Tule; SAV: Sierra Agua Verde; SLM: Sierra Las Mesteñas; SST: Sierra Santa Teresa.
Conclusiones: La biota de la sierra Santa Teresa está representada por crinoideos de las morfoespecies Baryschyr anosus Moore & Jeffords, Cyclocaudex insaturatus Moore & Jeffords, Floricyclus angustimargo Moore & Jeffords, Cyclocion distictus Moore & Jeffords, L. erathense, P. rugosum en asociación con algas, foraminíferos fusulínidos, esponjas coralinas (Chaetetes sp.), corales solitarios (Lophophyllidium sp., Fomichevella sp.), briozoos fenestélidos (A. stoyanowi) y braquiópodos (Antiquatonia sp.). La distribución de la biota, y particularmente de las morfoespecies de crinoideos, permitió correlacionar los registros con otras localidades del Misisípico-Pensilvánico de México y de distintas regiones de los Estados Unidos de América, principalmente en Texas, Colorado, Illinois y Oklahoma. Se considera que el paleoambiente inferido con base en los registros paleontológicos de la Sierra Santa Teresa se trataba de mares someros que permitieron el desarrollo de comunidades de crinoideos, así como otros invertebrados como esponjas coralinas, corales solitarios, briozoos fenestélidos y braquiópodos, con un rango estratigráfico del Misisípico Medio-Alto (Chesteriano) al Pensilvánico Medio (Desmoinesiano).
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

