











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
Nearctic-Neotropical migratory birds are species that breed in North America and migrate to Southern locations to spend the winter in tropical latitudes in Central and South America. They depend on the wintering grounds and migration stopover sites in tropical regions to fuel, spend long periods in different habitats, and then return to their breeding grounds (Amaya-Espinel & Hostetler, 2019; Gómez et al., 2017). The quality of the wintering grounds has a significant influence on the body condition, survival, and reproductive success of migrants (Marra et al., 1998; Marra & Holmes, 2001; Ruiz-Sánchez et al., 2017). Currently, land use change and deforestation of these regions threaten or imperil the stability of some migratory bird populations through time (Billington et al., 1996; Rappole & McDonald, 1994; Taylor & Stutchbury, 2016).
Recent studies using large datasets have reported the decline of populations of many Neotropical migratory birds in North America with insights from across their distribution (Ballard et al., 2003; Robbins et al., 1989; Rosenberg et al., 2019). These data come mainly from bird counts on their breeding grounds. However, some population parameters such as occupancy and conditions in terms of habitat and resources available in the Neotropics (non-breeding sites) are not fully understood. Understanding the population status of these mobile taxa requires the study of the occupancy in their geographic range across the annual cycle. At specific times of the annual cycle, Central America concentrates high numbers of Nearctic-Neotropical migratory birds (La Sorte et al., 2017). Therefore, surveys of highly threaten migratory species in different forest types in tropical regions during the non-breeding season are key to preserve habitats and to implement conservation actions.
The Wood Thrush (Hylocichla mustelina) is one of the Neotropical migratory birds experiencing dramatic population declines in recent decades (Rosenberg et al., 2019; Sauer et al., 2013). In their wintering grounds in Central America, the main processes that contribute to its decline are habitat loss and deforestation (Rosenberg et al., 2019). Nonetheless, there are protected and private forests remnants, and some agricultural practices (e.g., shaded coffee plantations) that provide important role as habitat for Neotropical migratory birds (Boza, 1993; Powell et al., 1992), especially in areas with high concentrations of individuals of vulnerable species in Mexico, Honduras, Costa Rica (La Sorte et al., 2017), and South America (Bayly et al., 2016). Geolocator data for migrating Wood Thrushes indicate rapid long-distance movements and substantial time spent in certain areas in Central America (Stutchbury et al., 2009). A study in Belize found that the number of individuals was higher in less disturbed forests in comparison with more open areas, and the different habitats varied in precipitation through time resulting in changes in resource availability and Wood Thrushes body condition (McKinnon et al., 2015). More studies detailing the use of remnant forests (Roberts, 2011) through time are needed to elucidate the threats in the wintering grounds relative to those in the breeding grounds (Stanley et al., 2015; Stutchbury et al., 2009).
The main goal of this study was to estimate the occupancy of Wood Thrush in Northern Costa Rica considering forest types and environmental variables with point count data. We asked the following questions: (a) What is the variation in the occupancy and detection probability of Wood Thrush in different forest types? We recorded the detection history (presence/absence) of Wood Thrush in three different forest types: dry forest in the Pacific slope, wet forests in the Caribbean slope (two sites), and in the cloud forest located in the Guanacaste Mountain Range dividing the two slopes (Janzen et al., 2016). These forests are land protected in the Área de Conservación Guanacaste (ACG). At these locations, we explored how the forest types and the average precipitation influenced Wood Thrush occupancy. We incorporated precipitation as a predictor of occupancy, since rain patterns are known to influence food-resource availability for migratory birds (Santillan et al., 2018). (b) What is the variation in occupancy of Wood Thrush in different vegetation types of the Caribbean wet forest and the probability of persistence through the months in the migration season? Previous studies have reported that the Wood Thrush preferred habitat in the wintering grounds is the rain forest (Stanley et al., 2015; Winker et al., 1990); therefore, we decided to explore further its habitat preference in the Caribbean forest of ACG. We considered the influence of vegetation structure and precipitation in the wet forest as predictors of occupancy during a migration period (2018-2019). To answer this question, we used multi-season occupancy models. We predicted that the probability of a site being occupied by Wood Thrush in time t and time t+1 (survivorship of a site) would increase with precipitation. As stated above, the habitat in the wintering grounds may be limited for migratory birds, and the conditions can change from year to year; for Wood Thrush, not all the areas are suitable when on the wintering grounds. If true, we expected those good or acceptable habitats would be occupied by Wood Thrush early in the season and that individuals would remain in these areas throughout the over-wintering period. If birds are not detected as time passes in the over-wintering season, we assume that they have died or moved. Although, it is more likely that relative changes in occupancy over a small spatial scale may suggest that birds have moved from one area to another. Here, we contributed to the understanding of Wood Thrush occupancy in a key conservation region of Costa Rica.
Materials and methods
Study species: Wood Thrush (Hylocichla mustelina) over-winter in late-successional forests and forest fragments with intermediate levels of habitat modification in Central America; from late Fall (August) until early Spring (March). In Costa Rica, they defend feeding territories from conspecifics (Roberts, 2007). They present high fidelity to their territories, specifically in large fragments and continuous forests (Roberts, 2007). As part of this territorial behavior, they respond to recorded playbacks, using both calls and songs (pers. obs.; Yahner & Ross, 1995). Wood Thrush mostly forages on the ground looking for litter arthropods mixed with fruit from understory shrubs and small trees (Roberts, 2007).
Study site: The study was carried out in the Área de Conservación Guanacaste (ACG) with an extension of 160 000 + ha, located in Northern Costa Rica (Fig. 1A), during December 2016, and in the 2018-2019 migration season. This region has three major vegetation types: dry forest (Pacific slope Sector Santa Rosa), cloud forest (Volcán Cacao in Guanacaste Mountain Range; hereafter Cacao), and wet forest (Pitilla Biological Station in the Caribbean slope) (Janzen et al., 2016) (Fig. 1B). The variation in forest types across the region provides an opportunity to explore where these birds spend in Northern Costa Rica most of the time, which forest types have a higher occupancy, and whether the precipitation has an influence across the study areas.

Fig. 1 A . Study sites in the Área de Conservación Guanacaste (ACG): Santa Rosa, Volcán Cacao, Pitilla Biological Station, and Leiva Biological Station, located in Northern Costa Rica. B. Pictures of the four study locations showing the different forest types (dry forest, cloud forest, and wet forest). C. Pictures of three different vegetation structure sites in Pitilla Biological Station (wet forest). A picture of the study species, Wood Thrush (Hylocichla mustelina), is included in the figure (Photo: Adrian Delgado).
Precipitation patters across the study areas are variable. Sector Santa Rosa has two defined seasons: the dry season from December to April (high temperatures regularly exceeding 37 °C, low relative humidity and near absence of precipitation) and the rainy season from May to November; with a mean annual precipitation of 1 700 mm and a mean annual temperature of 25 °C (Kalacska et al., 2004; Woodworth et al., 2018). The mean annual precipitation in the wet forests in ACG varied from 1 060-4 782 mm (data from Pitilla Meteorological Station 2016-2019), usually receiving ~ 4 000 mm of rain falls between May and February. The cloud forest area in Volcan Cacao has a mean average temperature of 18.5-21.7 °C (10 °C in December and January) and 3 000 mm of rain falls between May and January (Lindblad, 2001).
Point counts: To record the presence and absence of Wood Thrush, we performed point counts in December 2016: nine point counts in Santa Rosa (10.8379° N, 85.7051° W), eight in Cacao (10.9337° N, 85.4482° W), ten in Pitilla (10.9892° N, 85.4258° W), and 13 in Leiva (10.943220° N, 85.317962° W); 40-point counts total. Each point count was separated by 200 m along trails at each site. We performed the point counts over three consecutive days at each site.
One observer performed active point counts for three consecutive days (NVS) from 0530-0930 h. The observer played the call of a Wood Thrush recorded in Costa Rica (by Peter Boesman 2011) for 12 s at the beginning of each point (Fig. 2) and waited for 10 min, recording whether the response was a response call or a visual detection of the bird, both auditory and visual detection, or no detection. Migratory birds such as Wood Thrushes and Golden-winged Warblers responded to playbacks (Kubel & Yahner, 2007; Yahner & Ross, 1995). The use of playbacks was expected to increase the probability of detection of Wood Thrushes and this technique was a consistent way to define a point counts protocol for the study species.
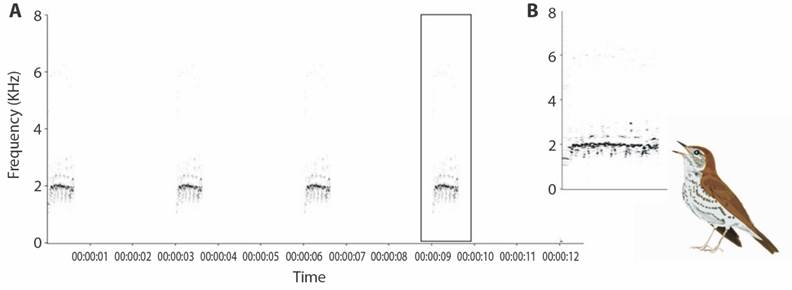
Fig. 2 A. Spectrographic view of the 12 s stimulus created to perform an active point count of a Wood Thrush call. B. Zoom in view of the Wood Thrush call.
To understand the variation of Wood Thrush occupancy during the non-breeding season, we performed active point counts in the Caribbean sector of ACG, at Pitilla Biological Station, in November 2018, December 2018, January 2019, and February 2019. The point counts were performed by three local parataxonomists that were familiar with the Wood Thrush calls and their responses to playbacks (calls or approaches to the sound source). Each person was assigned to an area with a different vegetation structure: open area (forest edge along a trail with dense understory at the edge), secondary forest, and old-growth forest (Fig. 1C). The parataxonomists recorded the presence and absence of Wood Thrush in nine points for the secondary and the old-growth forest, and in 12 points in the open area; following the same protocol described above.
Statistical analysis: Wood Thrush occupancy and detection probability for the four sectors of ACG were estimated using single-season occupancy models (MacKenzie et al., 2003; MacKenzie et al., 2017). Occupancy (psi) is defined as the probability that a randomly selected site or a sampling unit in an area of interest is occupied by a species; a site is occupied even with the detection of a single individual (MacKenzie et al., 2006). Julian Date and the site (ACG sector) were also included as covariates to account for the effect of sample day and different forest types, respectively. We used daily precipitation for the three consecutive days of counts and temperature as site covariates. To select the best model describing the Wood Thrush occupancy, we used Akaike’s Information Criterion adjusted for small sample size (AICc) (Burnham et al., 1995); we ranked the models by the difference (AICc) from the best-fitted model (AICmin).
We estimated Wood Thrush occupancy in different months in each vegetation type in the wet forest at Pitilla Biological Station using a multi-season occupancy model approach (MacKenzie et al., 2003; Webber et al., 2013). This type of model uses data from multiple sample periods, and the assumption of population closure is not expected between periods. We used daily precipitation collected a week prior to the survey days as a site covariate to explain Wood Thrush persistence. Site was used as a proxy for the movement of individuals in three vegetation types (open area, secondary forest, and old-growth forest). We defined movement when a site was occupied in time t and not occupied in time t+1. For the analysis, we divided the four months into three periods of time: period one (Nov-Dec 2018), period two (Dec 2018-Jan 2019), and period three (Jan-Feb 2019). The multi-season occupancy model calculated the parameter extinction (ε) used here as the probability of persistence (the probability of a site being occupied in a time t and the occupied in a time t+1) from one period to the next period. Multi-season models based on maximum likelihoods estimated occupancy (psi), colonization (γ), extinction (ε), and detection probability (p) for each site. We performed the occupancy models for single and multi-season data using PRESENCE software (Hines, 2006).
Results
Wood Thrush occupancy was best described by precipitation in the four sectors at the ACG, Costa Rica. As precipitation increased, the probability of Wood Thrush occupancy increased (Fig. 3, Table 1). The forest type (site) or Julian Date did not influence Wood Thrush occupancy or detection probability (Table 1). The wet forest had the highest Wood Thrush occupancy (Leiva and Pitilla sectors).
Table 1 Single season Wood Thrush occupancy (psi) models including detection probability (p). The covariates are precipitation (prep), temperature (temp), site (Área de Conservación Guanacaste sectors) and Julian Date (JDate). Models were ranked from the lowest to the highest AICc value.
| Models | nPars | AICc | delta AICc | AICwt |
| psi(prep)p(.) | 3 | 75.000 | 0.000 | 0.642 |
| psi(temp)p(.) | 3 | 77.910 | 2.820 | 0.157 |
| psi(site)p(.) | 5 | 78.620 | 3.530 | 0.110 |
| psi(.)p(.) Null | 2 | 80.230 | 5.140 | 1.049 |
| psi(.)p(JDate) | 3 | 80.780 | 5.690 | 0.037 |
| psi(.)p(sites) | 5 | 84.910 | 9.820 | 0.005 |
nPars = number of parameters.
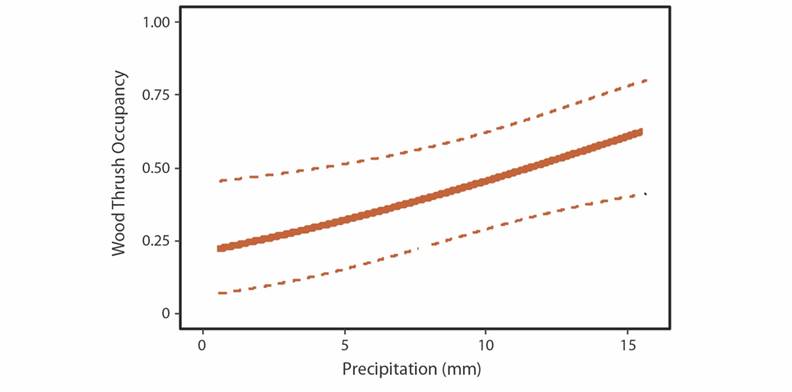
Fig. 3 Wood Thrush occupancy estimate explained by precipitation (mm) at the Área de Conservación Guanacaste (ACG), Northern Costa Rica. Point count data were collected in four ACG sectors (Santa Rosa, Cacao, Pitilla, and Leiva). Dashed lines indicate 95% CI intervals.
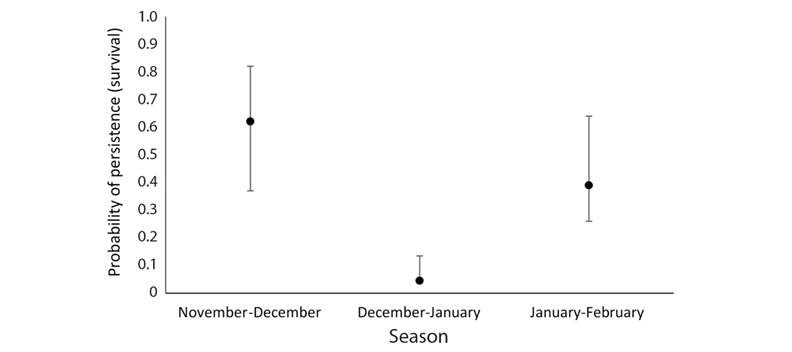
In the wet forest, we reported the occupancy at each vegetation type and how the probability of persistence varied according to changes in precipitation across the seasons. The occupancy of Wood Thrushes was higher in the open area (0.69 ± 0.09 SE), followed by the old-growth forest (0.61 ± 0.1 SE), and then the secondary forest (0.46 ± 0.1 SE). Wood Thrush probability of persistence in each site varied with precipitation, from November to December there was an increase in precipitation, then precipitation decreased from December to January (Fig. 4). In those sampling periods, the probability of persistence changed from high persistence as the rainfall increased (Nov-Dec) and changed to lower persistence (Dec-Jan) as the rainfall decreased (Fig. 5). From January to February the precipitation continued decreasing (Fig. 4); however, the probability of persistence of Wood Thrush in a site increased from January to February (Fig. 5).
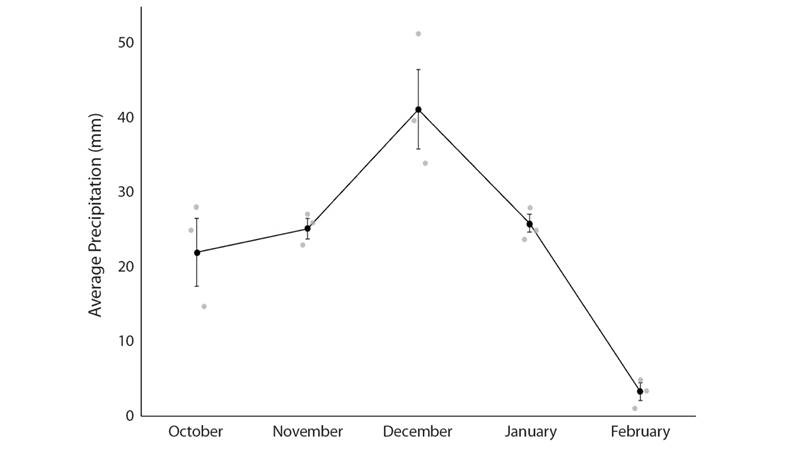
Fig. 4 Average precipitation (mm) collected for three days (gray dots) in a week previous to the point counts in Pitilla, Área de Conservación Guanacaste, Northern Costa Rica.
Discussion
Single-season occupancy results: These findings are congruent with studies for Wood Thrush (McKinnon et al., 2015) and other migrants elsewhere in the wintering grounds (Cespedes & Bayly, 2019; McKellar et al., 2013), showing a positive association between Wood Thrush occupancy and precipitation. Precipitation patterns are related with arthropod abundance in tropical forests, and consequently with habitat quality for migrant birds (Brown & Sherry, 2006; Sherry & Holmes, 1996). Therefore, higher occupancy is expected to change as the precipitation changes across the landscape. This region, the Western region of the ACG (wet forest), is particularly important since precipitation patterns change from the dry to the wet forest throughout the migration period.
Multi-season occupancy results: We observed 60 % of the point counts occupied during November-December by wood thrushes, and a decrease on the probability of persistence for the period December-January, in all the sampled sites. We predicted higher persistence of wood thrushes with higher levels of precipitation; therefore, it was expected to observe less persistence as the levels of precipitation decreased. However, for the last period (Jan-Feb) we expected to continue observing low Wood Thrush persistence, but we found an increase in persistence that was not related to precipitation. From January to February, as precipitation continued to decrease, we expected even more movements in the last season; however, it was the opposite according to the precipitation pattern.
The rationale of expecting more wood thrushes detections with increased precipitation (higher probability of persistence in a site from time t to time t+1) is associated with the relationship of precipitation with resource availability in terms of abundance of leaf-litter invertebrates in tropical wet forests (Lieberman & Dock, 1982). Therefore, Wood Thrush individuals would invest less time looking for food. As precipitation decreases because of the entrance of the dry season (Hilje et al., 2015) and therefore a decrease in food availability, it is expected that the birds invest more time foraging than in the previous period (Nov-Dec), decreasing the probability of persistence on the site. During the last season (Jan-Feb), Wood Thrushes decreased the movements, or in other words, the persistence probability was about 40 % on the sites, higher in comparison with the previous survey period (Fig. 5). A possible explanation for this pattern is the start of the breeding season of many resident birds (Stutchbury & Morton, 2001). In this scenario, Wood Thrushes might remain in the same areas, even though precipitation decreases, and therefore the abundance of leaf-litter arthropods decreases, to avoid conflict with territorial species such as wrens, ant-birds, and resident thrushes that share similar foraging substrates (Sánchez et al., 2014; Stratford & Stouffer, 2013). The increase of detections on the same sites during the survey period are also congruent with the establishment of Wood Thrush winter territories reported by Roberts (2007) and Roberts (2011). Therefore, they might be less mobile and avoiding conflicts with resident birds or saving energy before the spring migration.
In conclusion, we modeled Wood Thrush occupancy in Northern Costa Rica and the role of precipitation in four different sites within the Área de Conservación Guanacaste. Considering the three forest types (dry forest, cloud forest, and wet forest), we detected more birds in the wet forest of Northern Guanacaste, Costa Rica. In the wet forest, the probability of a site being occupied varied with precipitation which is correlated with food availability. Currently, the wet forests in Central America should be considered priority for Wood Thrush conservation. It is necessary; however, to evaluate the quality of protected and non-protected areas in terms of resources that these forests provide and evaluate the actual condition of the individual in the wintering grounds, as well as their interactions with resident species.
Ethical statement: the authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgments section. A signed document has been filed in the journal archives.

