











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
Environmental enrichment can be defined as any modification in the environment of captive animals which improves their biological functioning (Newberry, 1995). Enrichment provides the opportunity to display a wider range of adaptive behaviours and reduces the frequency of abnormal ones, enhancing animal welfare (Newberry, 1995; Young, 2013). There exist a diverse variety of enrichment types: nutritional, sensory, cognitive, social, physical features in their environment and so forth (Newberry, 1995; Young, 2013). Particularly, olfactory enrichment has been extensively used in mammals (Wells, 2009), especially for felids (Clark & King, 2008), but the importance of the olfactory dimension for other taxa such as reptiles and birds has been systematically overlooked (Nielsen et al., 2015). Birds kept in captivity often develop maladaptive conducts such as spot picking, route tracing and feather damaging behaviour (i.e. repetitive self-grooming including picking, plucking or chewing on feathers) (Brilot et al., 2009; Colton & Fraley, 2014; de Haas & van der Eijk, 2018; Jenkins, 2001; Mellor et al., 2018; Samson, 1996; van Hoek & Ten-Cate, 1998), this last one being particularly common in captive Psittaciformes (Jenkins, 2001; Mellor et al., 2021). Psittacine intellectual capacity appears to be similar to great apes and marine mammals (Kalmar et al., 2007; Krasheninnikova et al., 2019), hence, captive Psittaciformes require high levels of cognitive stimulation to prevent abnormal behaviours (Kalmar et al., 2010; Mellor et al., 2021; Rodriguez-Lopez, 2016). Although the benefits of implementing enrichment for parrots has been extensively described in the literature (Kalmar et al., 2007; Lumeij & Hommers, 2008; Rodriguez-Lopez, 2016; Young, 2013), it seems like routine enrichment for psittacines is constrained to only two types: foraging and physical modifications of their environment, whereas sensory-based enrichment has been neglected (Rodriguez-Lopez, 2016).
In the case of Ramphastids, there is a concerning lack of research regarding which types of enrichment they would benefit from, with only one study evaluating the effects of conspecific playbacks on a pair of Toco toucans Ramphastos toco (O’Brien, 2006). Psittacines and toucans are among the most common bird groups kept in rescue and rehabilitation centres across Central America (Hernández pers. obsv.). Thus, developing and implementing new enrichments to ensure high quality welfare standards must be a priority.
One challenge when it comes to designing new enrichment techniques is neophobia (Clark & King, 2008). Neophobia is described as the innate aversion to novel stimuli (Greenberg, 1990; Greenberg & Mettke-Hofmann, 2001). It is considered as an adaptive response because engaging an unknown object, food or location may entail being harmed or even killed by a predator, competitor, or poisonous food (Greenberg, 1990; Greenberg, 2003; Greenberg & Mettke-Hofmann, 2001). However, neophobia also carries disadvantages, as it prevents individuals from interacting with potentially new fitness-enhancing resources, obtaining less information, and losing the opportunity to exploit new food sources or microhabitats. Moreover, displaying a vigilant state can also divert energy and time from other vital activities such as foraging or mating (Crane & Ferrari, 2017). Hence, there exists a clear trade-off between the costs of displaying neophobia and its benefits (Greenberg, 2003; Greenberg & Mettke-Hofmann, 2001), and it is expected that this behavioural trait varies across species depending on their ecological particularities. Neophobia may be advantageous in species that inhabit high risk habitats or that exhibit a narrow or specialised trophic niche, while it could be detrimental for more generalist species which could benefit from exploring unpredictable and variable new resources (Brown et al., 2013; Greenberg, 2003; Greenberg & Mettke-Hofmann, 2001; Mettke-Hofmann et al., 2002). Furthermore, for species living in complex habitats, decreased neophobia would entail higher exploratory rates, providing a significant increase in acquired information which can impact the individual fitness (Biondi et al., 2010; Greenberg, 2003; Greenberg & Mettke-Hofmann, 2001; Mettke-Hofmann et al., 2002). However, it is also true that differences in neophobia can also be found within individuals of the same species, caused by previous experiences or personality types (Ensminger & Westneat, 2012; Fox & Millam, 2004; Herborn et al., 2010; Medina-García et al., 2017).
Therefore, the aim of this study was to evaluate differences in participation in an olfactory enrichment depending on the taxonomic group (Amazons Amazona / Macaws Ara / Toucans Ramphastos), as well as comparing neophobia response between them. Since neophobia seems to be particularly pronounced for at least some psittacine species (Mettke-Hofmann et al., 2002; Van Horik, 2014; Wilson & Luescher, 2006), we predicted that toucans would show a higher participation in the enrichment. In parallel, we also expected that the mean latency time to interact with the enrichment would be lower in toucans due to psittacine tendency towards neophobia.
Materials and methods
Study location and animals tested: Research was conducted in five Costa Rica wildlife rescue centers: The Toucan Rescue Ranch, Centro de Rescate Las Pumas, Parque Zoológico y Jardín Botánico Nacional Simón Bolívar, Rescue Center Costa Rica, and Rescate Wildlife Rescue Center. The study was performed with 73 red-lored amazons (Amazona autumnalis), 101 yellow-naped amazons (Amazona auropalliata), 52 scarlet macaws (Ara macao), 11 great green macaws (Ara ambiguus), 10 keel-billed toucans (Ramphastos sulfuratus) and 10 yellow-throated toucans (Ramphastos ambiguus) (Table 1). All individuals were adults, but sexes were unknown. Most of the tested birds were non-releasable animals because they were confiscated pets, lacking key behaviours to survive in the wild. Birds were held in communal aviaries, except for the toucans that were kept individually or in pairs. Enclosures were provided with natural vegetation and perches. Environmental enrichment was frequently provided by these centres, for example wooden toys, wrapped food, foraging devices with berries and so forth. The diet mainly consisted of fruits (banana, apple, papaya, pineapple, watermelon, cantaloupe), vegetables (carrot, broccoli, squash, celery, zucchini), beans, corn, and sunflower seeds. Also boiled eggs, pellets and worms were part of the diet for the toucans.
Table 1 Detailed description of the enclosures in each centre and the number of individuals of each species held in each facility.
| Center | Size of the enclosure (L × W × H, m) | Ara macao | Ara ambiguu | Amazona autumnalis | Amazona auropalliata | Ramphastos sulfuratus | Ramphastos ambiguus |
| Toucan Rescue Ranch | 10 × 3 × 4 | 6 | |||||
| Toucan Rescue Ranch | 10 × 3 × 4 | 6 | |||||
| Toucan Rescue Ranch | 10 × 4 × 4 | 10 | 13 | ||||
| Toucan Rescue Ranch | 3 × 2 × 3 | 2 | |||||
| Toucan Rescue Ranch | 5 × 3 × 3 | 2 | 3 | ||||
| Toucan Rescue Ranch | 2 × 2 × 2 | 2 | 2 | ||||
| Toucan Rescue Ranch | 2 × 2 × 2 | 4 | 2 | ||||
| Toucan Rescue Ranch | 10 × 4 × 6 | 2 | 2 | ||||
| Toucan Rescue Ranch | 10 × 4 × 6 | 2 | |||||
| Toucan Rescue Ranch | 4 × 2 × 4 | 1 | |||||
| Toucan Rescue Ranch | 5 × 3 × 4 | 1 | |||||
| Toucan Rescue Ranch | 4 × 2 × 4 | 1 | |||||
| Toucan Rescue Ranch | 4 × 2 × 4 | 1 | |||||
| Toucan Rescue Ranch | 4 × 2 × 4 | 1 | |||||
| Toucan Rescue Ranch | 4 × 2 × 4 | 1 | |||||
| Toucan Rescue Ranch | 4 × 2 × 4 | 1 | |||||
| Toucan Rescue Ranch | 4 × 2 × 4 | 1 | |||||
| Toucan Rescue Ranch | 4 × 2 × 4 | 1 | |||||
| Toucan Rescue Ranch | 4 × 2 × 4 | 1 | |||||
| Toucan Rescue Ranch | 4 × 2 × 4 | 1 | |||||
| Toucan Rescue Ranch | 4 × 2 × 4 | 1 | |||||
| Toucan Rescue Ranch Release Site | 10 × 4 × 4 | 11 | 4 | ||||
| Toucan Rescue Ranch Release Site | 10 × 4 × 4 | 10 | 4 | ||||
| Toucan Rescue Ranch Release Site | 4 × 2 × 4 | 2 | |||||
| Toucan Rescue Ranch Release Site | 10 × 4 × 8 | 6 | |||||
| Las pumas | 18 × 4 × 6 | 10 | 30 | ||||
| Las pumas | 10 × 8 × 4 | 3 | 5 | 10 | |||
| Las pumas | 8 × 6 × 4 | 5 | 15 | ||||
| Las pumas | 8 × 6 × 4 | 3 | |||||
| Simón Bolivar Zoo | 4.5 × 3.5 × 2.5 | 3 | |||||
| Simón Bolivar Zoo | 6 × 3 × 2.5 | 2 | |||||
| Simón Bolivar Zoo | 3 × 3 × 2.5 | 4 | |||||
| Simón Bolivar Zoo | 3 × 3 × 2.5 | 2 | |||||
| Simón Bolivar Zoo | 3 × 3 × 2.5 | 1 | |||||
| “Rescate” Wildlife Rescue Center | 8 × 6 × 6 | 10 | |||||
| “Rescate” Wildlife Rescue Center | 8 × 6 × 6 | 10 | |||||
| “Rescate” Wildlife Rescue Center | 6 × 3 × 4 | 1 | |||||
| “Rescate” Wildlife Rescue Center | 6 × 3 × 4 | 1 | |||||
| “Rescate” Wildlife Rescue Center | 8 × 6 × 6 | 7 | |||||
| Rescue Center Costa Rica | 10 × 3 × 3 | 4 | 10 | 16 |
All experiments were non-invasive and based exclusively on behavioural tests in which animals could decide whether to participate or not. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed. The experiments were performed in accordance with the ethical standards of each institution at which the studies were conducted, with Costa Rica animal welfare law (Ley No. 7451. Ley del bienestar de los animales, 1994) and with the Spanish Government animal research legislation (Real Decreto 53/2013, 2013).
Experimental design and behavioral data collection: Animals were exposed to an olfactory enrichment consisting of closed plastic containers (12 × 12 × 7 cm, all were opaque yellow) with small holes to allow the odours to come out following the Hernández et al., (2022) protocol which confirmed that at least toucans and macaws can discriminate between odours. Three different odour choices were simultaneously presented to the birds: a) water, b) apple vinegar, c) banana and papaya juice. Preliminary analyses showed that bird willingness to participate in the enrichment and latency times were not determined by the odour type, so we analyse them together. We kept the three-choice scent design because we demonstrated that it was interesting for the birds. Once they decide to engage with the enrichment, they alternate interest in the containers depending on what’s inside. Therefore, the neophobia/neophilia tendency analysed in this study was in response to the novel odour containers rather than to the odour itself, whereas previous studies revealed that the odour is the determinant of the total time they spend with each container (Hernández et al., 2022). Treatments consisted in eight cotton balls impregnated in 60 ml of one of each. The three plastic containers were placed in a random order inside the enclosures separated by 50 cm (Fig. 1). After installing the enrichment, the researcher left the enclosure and initiated the video recording of bird responses for 10 min. To establish comparable conditions, data collection was always performed before bird breakfast (between 6:00-8:00 h). Each enclosure was tested only once, to avoid pseudoreplication due to the inability to discriminate between individuals between videos (but we could discriminate between individuals within a video by following each bird movements throughout the entire video recording). Later, we analysed the video recordings to register how many individuals participated in the experiment and the latency. We considered participation in the study as the physical contact of the bird (touching any plastic container with the beak or foot to explore it) with the containers and we define latency as the delay between the exposure to the enrichment and bird physically interacting with it.

Fig. 1 Schematic illustration of the experimental design: Three scents (water, vinegar, papaya and banana juice) were simultaneously presented to the birds in separate plastic boxes. (Author: modified from Hernández et al., 2022).
Statistical analysis: To evaluate differences in bird participation between taxonomic groups (Amazona / Ara / Ramphastos), we performed a contingency table analysis using χ 2 and Cramer’s V tests. Bird latency to interact with the olfactory enrichment was analysed using a Generalized Linear Mixed effect model (GLMM), fitting normal distribution and identity link function. The response variable considered for the model was bird latency (s) (i.e. the delay between putting the olfactory enrichment in the enclosure and bird touching it) and we included the taxonomic group (Genus) as fixed factor (Amazona spp. / Ara spp. / Ramphastos spp.). We also set the individual as random factor to control for possible differences in animal personality traits. Preliminary analysis showed that there were no differences in the participation and latency of the birds between centers, so we did not include this variable in the models in order to have greater statistical power.
Results were considered significant at α < 0.05. Data are represented as mean (s) ± standard error (SE). The software used to perform the statistical analysis was SPSS 23.0 for Windows (IBM Corp, 2015).
Results
Bird participation: 257 birds were exposed to the olfactory enrichment: 175 amazons Amazona spp., 63 macaws Ara spp. and 19 toucans Ramphastos spp. Of them, only 28 (16.00 %) amazons Amazona spp., 12 macaws (19.04 %) Ara spp. and 10 toucans (52.63 %) Ramphastos spp. engaged in physical contact with the enrichment. Data analyses showed that there were differences in bird participation depending on the taxonomic group, with amazons and macaws being much less inclined to interact with the olfactory enrichment compared to toucans (χ 2 = 14.686, d.f. = 2, P = 0.001; Cramer’s V = 0.239, P = 0.001, Fig. 2).

Fig 2 Relationship between the number of individuals of each taxonomic group (Amazons Amazona spp. / Macaws Ara spp. / Toucans Ramphastos spp.) exposed to the olfactory enrichment and the number of individuals which participated in it: A. Absolute numbers and B. Percentages.
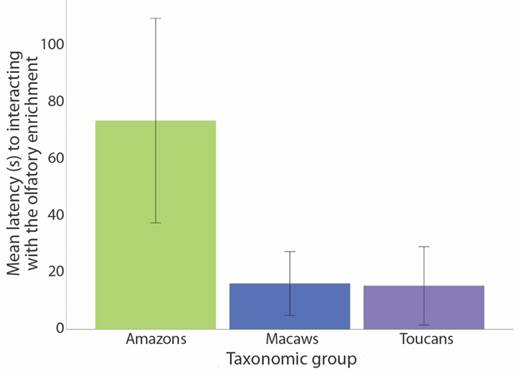
Bird latency time to interact with the enrichment: for birds participating in the enrichment, according to GLMM, the mean latency time was higher (73.11 ± 17.88 s) in amazons, Amazona spp., than for Ara spp. (16.08 ± 5.60 s) or Ramphastos spp. (15.30 ± 6.85 s) (Table 2, Fig. 3). Furthermore, the individual identity has influence as random factor over the variance of latency response of the birds (Table 2).
Table 2 Results of the GLMM analysing latency time (s) exhibited by the birds which interacted with the olfactory enrichment depending on the taxonomic group (Amazons Amazona spp. / Macaws Ara spp. / Toucans Ramphastos spp.) and the individual as random factor.
| Factors | F | df | P |
| Corrected model | 3.814 | 47 | 0.029 |
| Genus | 3.814 | 47 | 0.029* |
| Individuala |
aRandom factor. (Estimate = 2 657.30, SE = 1 097.14, Z-test = 2.424, P = 0.015). *Genus Amazona (SE= 26.86; t = 2.152; P = 0.037).
Discussion
The results of this study showed that bird participation in the olfactory enrichment was determined by the taxonomic group. As we expected, toucans engaged with the smell containers more frequently than amazons and macaws. This finding indicates that toucans are more prone towards neophilia compared to psittacines. Keel-billed and yellow-throated toucans have been traditionally considered generalist frugivores (Remsen et al., 1993), but they are also opportunistic foragers which consume invertebrate and vertebrate preys, eggs and even carrion (Cove et al., 2016; Davlantes & Howe, 2018; Romero et al., 2022; Zúñiga-Ortiz, 2014). Our results may suggest that keel-billed and yellow-throated toucans are neophilic species, since neophobia would be disadvantageous for opportunistic species, as they would lose chances to exploit unpredictable and unknown resources. On the contrary, red-lored amazons, yellow-naped amazons, scarlet macaws, and great green macaws, are not omnivorous species and only feed on seeds, fruits, flowers, leaves and bark (Benavidez et al., 2018; Berg et al., 2007; Vaughan et al., 2006; Volpe et al., 2022), which may explain their tendency towards neophobia. The main limitation of this study was the fact that birds were kept in different housing conditions and that we could not control the possible influence of social competition in the willingness of the birds and the latency times showed to approach the enrichment. Amazons and macaws are highly social species which benefit from being kept in communal aviaries (Seibert, 2006; van Zeeland et al., 2009) while toucans are usually held alone or in pairs because they are territorial, especially during breeding season (Bor-Mikich et al., 2008). It is probable that the hierarchy among parrots could delay the interaction of lower rank individuals with the scent containers. Consequently, further research is warranted to gain insight into how hierarchy influences participation in this type of enrichment. In large groups, we recommend including as many enrichment containers as possible, to guarantee that all individuals have access to it. In this regard, further research would be needed, testing animal responses individually to further verify our results.
Regarding the differences found in mean latency time, we found the delay to interacting with the olfactory enrichment was significantly higher in amazons than in macaws and toucans. These results suggest that amazons are particularly cautious when it comes to exploring new objects in comparison to macaws and toucans. This finding can be also linked to a neophobic tendency in red-lored amazons and yellow-naped amazons, explained by their life-history traits. As we mentioned above, amazons do not prey on other animals nor opportunistically consume eggs or carrion as toucans do. Moreover, amazons are much smaller psittacines than macaws so they may be easier to prey upon, whereas macaws’ predators mainly affect the nest (Fraixedas et al., 2014; Schruhl et al., 2012). This could entail that unknown settings are riskier for amazons than for macaws, so they would benefit from a neophobic approach.
Moreover, it is also important to highlight that there were individual differences in the latency time. As previously described in the literature, personality and previous experiences can affect neophobia responses (Ensminger & Westneat, 2012; Fox & Millam, 2004; Herborn et al., 2010; Medina-García et al., 2017). Therefore, each bird response to the enrichment was not only determined by the taxonomic group, but probably also by personality types and the reason why they came to the rescue centres. Many birds were confiscated because they were pets while others were born and raised in the wild but suffered from injuries or other health problems. Future studies could focus on evaluating differences in personality traits and neophobic behaviour depending on the animal background history.
We found that participation success was higher in toucans than in amazons and macaws possibly because of a lesser neophobic reaction to the containers. Red-lored amazons and yellow-naped amazons seemed to be particularly neophobic species, showing higher latency times to interact with the enrichment, probably due to their life-history attributes. Our results indicate that the container determined the neophobic/neophilic response, whereas the odour inside would determine the time spend which each container (Hernández et al., 2022). We also highlight that keel-billed and yellow-throated toucans are highly participative in environmental enrichments, so it would be of relevance to routinely include this kind of enrichment in captivity welfare plans. In the case of amazons and macaws, they seem to be more cautious so it may be more successful to present odours in natural containers or to apply them on features in their enclosure. Finally, the next step would be to evaluate if this kind of enrichment produces differences in bird conduct (more desirable behaviours and less stereotypies) and in the physiological stress response, to properly assess its impact on animal welfare.
Ethical statement: the authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgments section. A signed document has been filed in the journal archives.

