











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
The eastern tropical Pacific has suboptimal oceanographic conditions for coral community development (Glynn & Ault, 2000). The Mexican Pacific coast is a region with high primary productivity enhanced by the nutrient enrichment coming from riverine discharge, upwelling currents, and a high seasonal temperature variability jointed with high turbidity of the water column that difficult coral survival (Reyes-Bonilla, 2003). The development of human settlements near coral communities also has exerted negative impact through the discharge of sewage, solid waste, and land transformation, causing coastal erosion and enhancing the sedimentation rate at some sites harboring corals (Nava & Ramírez-Herrera, 2012). Added to these anthropized conditions is the impact of the El Niño phenomenon, which in the last decades has decimated entire populations of reef corals at some localities in this region (Nava et al., 2019). Some of these impacts, chronic and silent, exceptional or devastating, have occurred without being recorded in detail. This is not the case with the coral communities from Zihuatanejo, which remain among the most conserved on the Mexican central Pacific coast. These communities vary in their extension and structural characteristics, but all have provided relevant life support systems to the inhabitants of the municipality of Zihuatanejo de Azueta for 400 years (Sánchez-Briones, 2010). Since that time, the activity of traditional fishing has been part of the identity of the inhabitants of this municipality and the region (Reyes-García, 2012). Since the decade of 1980, Playa las Gatas has been an important tourist destination in Zihuatanejo, allowing the establishment of several service providers offering lodging, food, and recreational services. The first studies of the conservation state of five locations with coral communities in the area began in 2009, which are Islote Zacatoso, Playa las Gatas, Caleta de Chon, Playa Manzanillo, and Playa Riscalillo (Fig. 1). Such studies provided relevant information about the environmental conditions of each site, suggesting that these communities are under pressure of anthropogenic origin (Nava & Ramírez-Herrera, 2012). Data on the coverage of living corals and their ratio concerning dead coral substrata have evidenced the presence of locations with reef patches with an acceptable conservation status (Nava & Ramírez-Herrera, 2012; Nava et al., 2019; Nava et al., 2021). These data suggest such communities present a high potential to develop restoration initiatives. Assuming the process of sponge bioerosion as an indicator of the conservation state of the reef ecosystem, the survey of boring sponge abundance in reefs from Zihuatanejo revealed that bioerosion may weaken the reef framework in locations with high cover of dead corals (Nava et al., 2019). Previous tests of a reliable and low-cost strategy for coral transplantation on rocky substrata showed to be effective and that it can be used by local inhabitants at sites with reduced coral coverage. Nonetheless, the impact of the event of the El Niño of 2015-16 showed that it is necessary to implement a multidisciplinary strategy to restore the coverage with living corals resistant to thermal stress (Cárdenas-Alvarado et al., 2021). The algal symbionts of corals (Symbiodiniaceae) are relevant since they provide corals with nutrients and oxygen and allow them to increase their calcification rate (Muller-Parker et al., 2015). Since Cladocopium and Durusdinium are the most tolerant genera to high seawater temperatures (Baker et al., 2004; LaJeunesse et al., 2018), we aimed to test if the identity of algal symbionts may influence the response of coral bleaching in Zihuatanejo and now are attempting restoration emerging from a citizen science approach. Recent research has demonstrated that local inhabitants are owners of their elaborated ecological knowledge that is used to manage natural resources from coral communities (Drouet-Cruz, 2020). Current efforts aim at integrating all available information to sustain strategic alliances among academic, citizen, and government agencies to develop restoration initiatives based on the participation of local inhabitants that promote coral communities restoration with thermally resistant reef corals. This contribution provides an analysis of the outcome of this work in process and proposes an approach to establish a long-term initiative for the conservation of coral communities from Zihuatanejo, Guerrero.
Materials and methods
Study area: The municipality of Zihuatanejo de Azueta is located at 17° 37'- 17° 41'N and 101° 31'- 101° 39'W. Throughout the coast, there are abrupt rocky formations alternated with sandy beaches with variable slopes. The most evident landforms are headlands, cliffs, coves, islets, estuaries, and a coastal lagoon. The bottom substrate type varies according to the geographic orientation and it is divided into two main areas: the upper northwest, where rocky outcrops dominate with a cover approximately 70 %, and the lower southeast, covered mainly by sandy bottoms, reaching an 80 %. Along this area there are distinct locations with relevant coral communities whose distribution varies from very shallow areas to more than 20 m in depth. Some of these locations are Islote Zacatoso, Playa las Gatas, Caleta de Chon, Playa Manzanillo, and Playa Riscalillo (Fig. 1).

Fig. 1 Detail of the study area and coral communities in Zihuatanejo, Guerrero. A. location of the sites in the study area, B. coral reef in Islote Zacatoso, C. coral community of Playa las Gatas and D. coral bleaching of Playa Manzanillo during the El Niño 2015-16. Images: Héctor Nava.
Islote Zacatoso (17° 39'13.2” N and 101°37'13.3” W) is an islet located 1 km southwestward to the Ixtapa Zihuatanejo village. There is a monospecific fringing reef, located at the islet area protected from the surge, that spreads out from 1 m to near 6 m in depth. It is mainly composed by Pocillopora damicornis, and P. verrucosa, although other species, like P. capitata, Pavona gigantea and Porites lobata are also present. This reef seems to be well conserved. The access to this site is only with boats and the use of the reef includes low-impact artisanal fishing and non-intensive recreational scuba diving (only few scuba agencies bring a reduced amount of scuba divers to visit the reef each week).
Playa las Gatas (17° 37'16.8” N and 101° 33'8.55” W) is inside the Zihuatanejo Bay. The coral community in this area is mainly composed by colonies of the genera Pocillopora, Pavona, and Porites, growing over a substratum composed by boulders that can reach 2 m in diameter and distributed between 3 and 6 m in depth. Since more than three decades ago, these corals have been exposed to strong anthropogenic pressure by locals and tourists (mainly boat anchoring, touching corals and domestic sewage discharge). Nowadays, this coral community presents the lowest cover of living corals.
Caleta de Chon (17° 36'52.8” N and 101° 33'16.3” W) harbors a fringing reef that extends from 1 m to near 7 m in depth and is composed of reef corals of the genera Pocillopora and Pavona that build a reef framework of more than 1 m in height. To date, this reef presents one of the highest covers of living corals in Zihuatanejo and spreads out parallel to the shoreline at the head of the cove. This impact comes from the construction of roads and buildings near the cove that have caused land erosion at least during the last 10 years. The main access to this site is by boat, but the opening of roads allows the access of visitors by land.
Playa Manzanillo and Playa Riscalillo (17° 37'11.4” N, 101° 31'27.6” W and 17° 37'0.7” N, 101° 31'3.5” W) are areas close to each other, with monospecific fringing reefs that spread out from the surface level to 6 m in depth. Until the event of the El Niño of 2015-16, both reefs were the best preserved in the area. To date, the reef of Playa Riscalillo is highly damaged, although both reefs are far from inhabited sites and access is mainly by boat; the only human activities at the sites are snorkeling, scuba diving, and artisanal fishing.
Baseline study of the conservation status of coral reefs: The coverage of the major components of the substrata was recorded by the Marine Biodiversity Laboratory of the Universidad Michoacana de San Nicolás de Hidalgo. Coverage of live corals (LC), dead coral framework (DC), algae, sand, and rocks was quantified using 20m-long photo-transects (five transects in 2010, three transects in 2015 and 2018) recording the proportion (%) covered by each category in ten photo-quadrants of 1m2 per transect. Each image was analyzed with CPCe (Coral Point Count with Excel extensions) software developed by the National Coral Reef Institute (FL, USA), using 100 random dots. To assess the conservation status of each coral reef, the coral mortality index (CMI) (Gómez et al., 1994) was used. This index is based on the ratio between the coverage of living and dead corals, and values near zero indicate a dominance of living corals (well conserved coral reef) and values near 1 indicate a dominance of dead corals (deteriorated coral reef):
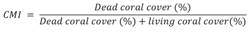
The environmental conditions were characterized in each site recording the sedimentation rate using sediment collecting bottles (n = 6) and the transparency of the water column using the Secchi disk method (n = 3). The concentration of total suspended solids (mg·l-1, n = 3) and chlorophyll in the water column were measured in samples of 3l of seawater (mg·m3, n = 3). The records of 𝛿15N and 𝛿13C in samples of dried Amphiroa sp. (n = 5) because were taken due both parameters are indicators of terrestrial sources of nitrogen and carbon (see Nava et al., 2014 for more details).
Assessment of the sponge bioerosion in the reef framework: The species richness and abundance of boring sponges were recorded at each coral reef collecting 9 cm3 coral fragments of three types of substrata: living corals (n = 25), dead corals (n = 25), and coral rubble (n = 25) developed by Pocillopora sp. along a 50 m long transect and replicating this sampling in three transects. Each coral fragment was broken into small pieces and revised with a stereoscope to locate tissue of boring sponges inside the fragment. Small samples of sponge tissue were digested with sodium hypochlorite and observed in an optic microscope to confirm the presence of boring species. The total abundance was recorded as the proportion (%) of all 75 fragments collected per transect and containing boring sponges, averaged from the tree replicates (mean ± standard deviation). The abundance of boring sponge per substrate type was recorded as the proportion (%) of the 25 fragments collected per transect and containing boring sponges, averaged from the three replicates (mean ± standard deviation) (Nava et al., 2014).
Experimental test of the transplantation technique of living coral fragments: The experimental technique for transplanting living corals was implemented in Playa las Gatas, a site with low coverage of living corals (~ 4 %) and abundant rocky boulders (57 %), as well as high content of loose fragments growing on the sand, considering these as an opportunity to be used as transplants (Table 1, Fig. 1C). This study assessed two treatments of transplantation of living corals (Fig. 2): 1) using freshly dislodged coral fragments 10 cm long, without signs of healing in their fractured section (named coral fragments) and 2) using loose coral fragments of the same size that already healed the damage suffered during the dislodgement (named loose coral colonies). Such discrimination was taken into account assuming that coral fragments will show a faster fixation to the rocky substrata due to their activated process of regeneration of the fractured section. Likewise, we also evaluated the possible effect of environmental seasonality, starting the first experiment at the beginning of the rainy season and a second experiment at the beginning of the dry season. All corals from both treatments were transplanted onto the surface of rocky boulders using steel grids of 60 cm per side, with square division of 15 cm, which were tied with 12-gauge wire to five boulders for each treatment. A total of 25 coral transplants were established on the surface of each boulder, immobilizing them below the grid intersections and tying them with plastic zip ties (Fig. 2). The mean proportion (%) of these 25 transplants fixed on the rock was recorded quarterly for 12 months. The growth of transplants was also measured with a 30 cm metal ruler as the increase of the maximum diameter in the vertical (mean vertical growth) and horizontal (mean horizontal growth) planes considering their original size as 100 %. The mean proportion of survival of transplants (%) was recorded as the proportion of the 25 transplants that still remained alive during each sampling.
Table 1 Record of the coverage of major substrata components and coverage of living corals in the coral communities of Zihuatanejo, Guerrero.
| LC | BC | PC | DC | CR | FA | RO | SN | CMI | ||
| 2010 (n = 5) | Islote Zacatoso | 65.85 | 0.00 | 0.00 | 12.04 | 6.92 | 5.10 | 13.76 | 4.86 | 0.16 |
| 16.86 | 0.00 | 0.00 | 5.33 | 3.28 | 3.46 | 7.30 | 5.04 | 0.09 | ||
| Playa las Gatas | 8.00 | 0.00 | 0.00 | 15.13 | 9.30 | 6.00 | 61.71 | 9.85 | 0.68 | |
| 6.10 | 0.00 | 0.00 | 1.94 | 5.07 | 4.34 | 5.97 | 2.87 | 0.15 | ||
| Caleta de Chon | 61.11 | 0.00 | 0.00 | 13.87 | 8.95 | 6.78 | 16.64 | 8.23 | 0.19 | |
| 12.74 | 0.00 | 0.00 | 8.58 | 1.54 | 5.30 | 6.76 | 9.88 | 0.12 | ||
| Playa Manzanillo | 54.95 | 0.00 | 0.00 | 29.49 | 11.36 | 19.06 | 7.56 | 6.03 | 0.35 | |
| 14.34 | 0.00 | 0.00 | 11.62 | 4.83 | 12.90 | 5.33 | 3.06 | 0.15 | ||
| Playa Riscalillo | 77.23 | 1.01 | 0.00 | 8.27 | 2.60 | 5.64 | 11.48 | 1.79 | 0.10 | |
| 15.68 | 2.26 | 0.00 | 6.41 | 1.10 | 4.26 | 9.94 | 1.13 | 0.09 | ||
| 2015 (n = 3) | Islote Zacatoso | 52.24 | 0.00 | 27.44 | 2.71 | 5.09 | 5.61 | 6.90 | 0.00 | 0.03 |
| 35.74 | 0.00 | 35.23 | 1.07 | 3.98 | 9.72 | 5.25 | 0.00 | 0.01 | ||
| Playa las Gatas | 0.37 | 31.89 | 1.97 | 0.74 | 5.91 | 0.08 | 49.64 | 9.40 | 0.02 | |
| 0.65 | 20.98 | 2.93 | 0.58 | 2.15 | 0.14 | 15.13 | 7.20 | 0.01 | ||
| Caleta de Chon | 58.13 | 0.27 | 0.00 | 5.51 | 7.20 | 2.99 | 25.73 | 0.18 | 0.09 | |
| 5.09 | 0.46 | 0.00 | 4.81 | 4.57 | 5.17 | 9.32 | 0.31 | 0.07 | ||
| Playa Manzanillo | 0.49 | 9.63 | 40.09 | 30.26 | 13.11 | 0.00 | 5.94 | 0.48 | 0.43 | |
| 0.84 | 15.44 | 27.09 | 14.36 | 11.99 | 0.00 | 7.64 | 0.49 | 0.32 | ||
| Playa Riscalillo | 0.00 | 28.76 | 42.91 | 17.13 | 0.33 | 0.00 | 10.87 | 0.00 | 0.20 | |
| 0.00 | 13.21 | 26.85 | 11.82 | 0.57 | 0.00 | 4.12 | 0.00 | 0.15 | ||
| 2018 (n = 3) | Islote Zacatoso | 79.93 | 0.00 | 0.00 | 3.82 | 3.33 | 1.42 | 12.92 | 0.00 | 0.05 |
| 12.46 | 0.00 | 0.00 | 2.48 | 3.25 | 2.45 | 7.42 | 0.00 | 0.03 | ||
| Playa las Gatas | 0.50 | 0.00 | 0.00 | 15.00 | 20.73 | 5.00 | 56.08 | 7.02 | 0.94 | |
| 0.50 | 0.00 | 0.00 | 8.66 | 12.26 | 3.61 | 24.61 | 4.39 | 0.09 | ||
| Caleta de Chon | 82.33 | 0.00 | 0.00 | 0.73 | 2.00 | 0.00 | 13.18 | 1.75 | 0.01 | |
| 8.19 | 0.00 | 0.00 | 1.27 | 2.00 | 0.00 | 8.09 | 2.22 | 0.01 | ||
| Playa Manzanillo | 62.65 | 0.00 | 0.00 | 22.75 | 2.42 | 10.25 | 11.43 | 1.00 | 0.33 | |
| 10.03 | 0.00 | 0.00 | 12.76 | 1.46 | 9.50 | 7.81 | 1.00 | 0.02 | ||
| Playa Riscalillo | 24.42 | 0.00 | 0.00 | 59.17 | 0.50 | 25.17 | 16.08 | 0.33 | 0.67 | |
| 23.07 | 0.00 | 0.00 | 36.11 | 0.87 | 18.11 | 13.13 | 0.58 | 0.35 |
LC = living corals, BC = bleached corals, PC = pale corals, DC = dead corals, CR = coral rubble, FA = filamentous algae, RO = rocks, SN = sand, and CMI = coral mortality index. In bold: the mean proportion (%). In cursives: standard deviation.

Fig. 2 Detail of coral transplant experiments in Playa las Gatas. A. loose coral colonies at the beginning of the experiment, B. after 13 months, C. coral fragments at the beginning of the experiment and D. after 13 months.
Assessment of the genetic diversity of algal symbionts (Symbiodiniaceae) in coral populations of Zihuatanejo, Guerrero: The Laboratory of Genetics for Conservation of the Institute of Research in Ecosystems and Sustainability of the National Autonomous University of Mexico coordinates this line of research. We evaluated the composition and the genetic diversity of the populations of algal symbionts in four coral communities in the area (Islote Zacatoso, Playa Las Gatas, Caleta de Chon, and Playa Riscalillo) after the event of El Niño of 2015-2016. We obtained DNA of 102 coral fragments of Pocillopora verrucosa, which were randomly sampled between 4 and 12 m in depth in each site during January and June 2018. Total DNA was extracted using the Invitrogen PureLink™ Genomic DNA Mini Kit following the manufacturer's instructions with slight modifications (Cárdenas-Alvarado et al., 2021). Afterwards, a fragment of the 28S ribosomal RNA gene and a fragment of the Internal Transcribed Spacer 2 (ITS2) of the ribosomal RNA genes were amplified with the primer pairs designed by Tong et al. (2018) and by Appril and Gates (2007), respectively. To determine the identity of zooxanthellae, we constructed two phylogenetic trees with the sequences of the 28S and ITS2 regions from this study and the different genera of Symbiodiniaceae from GenBank. The genetic diversity of zooxanthellae for each reef was calculated with haplotype diversity (h) and nucleotide diversity (π).
Record of the local ecological knowledge: Understanding local knowledge and perception is especially relevant when promoting citizen participation in conservation efforts as local ideas of ecosystem functioning are valuable sources of useful long-duration data points that serve as the blueprints for participatory conservation plans. For this reason, this project stands on methodologies recording the local knowledge rigorously and systematically and the CoLaboratories of Social Oceanography of the El Colegio de Michoacán is working towards this goal. Qualitative social data used to record and analyze local ecological knowledge of corals and coral reefs were obtained through a mixed methods approach. Firstly, to assess the collective perception about the composition and diversity of the coral community we used free listing. This technique is a simple and easy way to obtain and systematize empirical data (Quinlan, 2005) and is generally used to identify elements of a cultural domain (e.g., fish species) (Smith et al., 1995). A second component of our mixed methods was an ethnographic approach based on colloquial conversations and unstructured interviews. This approach allowed interaction with more than eighteen individuals representing the fishing, tourism, and governmental sectors in Zihuatanejo. The interactions were recorded as notes and these, in turn, were transformed into a field log (Bernard, 1995). The starting point, Drouet-Cruz's work (2020), is now moving forward by fostering characterization of the sociopolitical context. By accounting the connections among different actors and powerful groups that have access to the coral reef, we look to foster new collaborative strategies to protect coral communities and manage their diversity. Understanding the social, political, and economic context will allow for designing strategies of citizen participation that can be strengthened and appropriated by the people of the Azueta municipality. The strategies that this project considers 1) to include a diversity of local actors and deep knowledge about the biology and ecology of corals, 2) to incentivize dialogue and exchange among local and scientific knowledge, to generate a common language that facilitates collaboration, 3) to make available to local stakeholders the technical information, that allows them to participate in tasks destined to restore and monitor the coral communities, 4) to encourage the creation of collaborative networks among all actors, to implement actions that preserve these coral communities, and 5) to incentivize the proposal of new local initiatives for this purpose. In this way, we expect that this initiative can endure.
Data analysis: Both environmental and biological data were tested for normality and homogeneity of variances (Kolmogorov Smirnov test). In the case of data not normally distributed, they were square-root transformed. A series of separate one-way analyses of variance (ANOVA) were conducted on the environmental and biological parameters to determine significant differences in the coverage of substrata and abundance of boring sponges. The effectiveness between transplantation techniques were determined with two-way analysis of variance (ANOVA) with repeated measures, in which the replication factor was the sampling periods and the independent factor was each parameter measured (proportion of transplant fixation, growth, and survival among either seasons, locations and treatments).
Results
The conservation state of the coral reefs from Zihuatanejo: The data provided from this study in 2010 confirmed the existence of locations with a relevant coverage of living corals, being high in almost all locations (between 54.95 ± 14.34 % and 77.23 ± 15.68 %) with the exception of Playa las Gatas, where the cover of living corals was the lowest (8.00 ± 6.10 %) (F = 15.619, P < 0.001). The cover of dead corals was low at almost all locations (between 8.27 ± 6.41 % and 15.13 ± 1.94 %) with exception of Playa Manzanillo (29.49 ± 11.62 %) (F = 5.817, P < 0.005). The lowest record of CMI (0.10 ± 0.09) was made at the most preserved sites and the highest record of CMI was found in Playa las Gatas, the closest site to the human settlements (0.68 ± 0.15) (F = 17.862, P < 0.001, Table 1). Some of the symptoms of environmental degradation during 2010 were high rates of sedimentation in Playa Riscalillo (1.57 ± 0.27 kg·m-2 d-1) and high levels of d15N and low water transparency in Islote Zacatoso (9.49 ± 0.43 and 7.46 ± 1.99 m, respectively) as well as high levels of d13C and a high chlorophyll concentration for Playa las Gatas (up to -13.25 ± 0.67 and 1.82 ± 0.86 mg m-2 d-1) (Table 2).
Table 2 Averaged record (mean ± standard deviation) of sedimentation rate (Sed), water transparency (Trans), total suspended solids (TSS), chlorophyll concentration (Cla), levels of d15N, and d13C recorded at Islote Zacatoso (IZ), Playa las Gatas (PG), Caleta de Chon (CC), Playa Manzanillo (PM), and Playa Riscalillo in 2010*
| Sed (kg m-2 d-1) n = 6 | Trans (m) n = 3 | Cla (mg · m-3) n = 3 | TSS (mg · l-1) n = 3 | d15N n = 5 | d13C n = 5 | |
| IZ | 1.03 ± 0.49 | 7.46 ± 1.99 | 0.79 ± 0.11 | 23.69 ± 9.85 | 9.49 ± 0.43 | -14.55 ± 0.63 |
| PG | 0.48 ± 0.19 | 3.71 ± 0.70 | 1.82 ± 0.86 | 18.74 ± 11.57 | 9.23 ± 0.50 | -13.25 ± 0.67 |
| CC | 1.23 ± 0.27 | 5.45 ± 0.95 | 0.71 ± 0.43 | 34.75 ± 6.60 | 9.35 ± 0.47 | -14.44 ± 0.70 |
| PM | 0.71 ± 0.14 | 5.97 ± 1.90 | 0.27 ± 0.11 | 32.35 ± 9.53 | 8.38 ± 0.53 | -15.67 ± 0.97 |
| PR | 1.57 ± 0.27 | 6.13 ± 1.70 | 0.58 ± 0.07 | 27.43 ± 11.97 | 8.84 ± 0.34 | -14.07 ± 1.06 |
* Modified from Nava et al., 2014.
Boring sponge abundance: A total of 12 species of boring sponges belonging to the genera Cliona, Pione, Siphonodictyon, and Thoosa were recorded at the study area. Among all sites, Caleta de Chon (59.9 ± 11.2 %) was the most affected site by sponge bioerosion and Playa Manzanillo (23.2 ± 4.9 %) showed the lowest abundance of boring sponges (F = 12.170, P < 0.001). The dead corals were the most invaded substrata by boring sponges (63.72 ± 25.165) and living corals were the less affected (25.12 ± 12.69 %) (F = 34.982, P < 0.001), with the highest records in Playa Riscalillo and Caleta de Chon (up to 86 ± 19.8 %) (F = 4.567, P < 0.01). The substrata composed by living corals showed the highest invasion by boring sponges in Caleta de Chon (up to 32.8 ± 7.2 %) in comparison to Playa las Gatas and Playa Manzanillo (up to 12.8 ± 7.6 %) (F = 4.359, P < 0.01, Fig. 3).
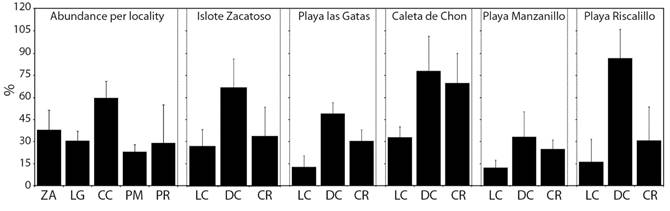
Fig. 3 Average abundance of boring sponges per locality and by each type of coraline substrata. ZA = Islote Zacatoso, LG = Playa las Gatas, CC = Caleta de Chon, PM = Playa Manzanillo, PR = Playa Riscalillo, LC = living corals, DC = dead corals, and CR = coral rubble. Bars above the columns indicate the standard deviation.
Experiences in coral transplantation: During the dry season, the fixation of coral transplants was faster in coral fragments (98.00 ± 4.0 % in 9 months) in comparison to the loose coral colonies (86.43 ± 18.86 % in 12 months) (F = 6.258 P < 0.05), putatively caused by the environmental stability. The vertical and horizontal growth of coral fragments were almost twice as large (161.18 ± 68.87 % and 106.80 ± 24.45 %, respectively) as in the loose coral colonies (88.29 ± 32.12 % and 72.94 ± 7.58 %, respectively) (F = 8.035, P < 0.05 and F = 15.274, P < 0.05, Fig. 4). Survival at the end of the experiment made during the dry season was higher in coral fragments than in loose coral colonies (90.98 ± 6.48 % vs. 62.80 ± 43.42 %) (F = 3.061, P < 0.05). During the rainy season, the maximum fixation occurred after 8 months in coral fragments (89.27 ± 7.18 %), contrasting the record made in loose coral colonies, whose maximum record was made only after 11 months (84.46 ± 13.09 %) (F = 20.489, P < 0.05, Fig. 4). The vertical and horizontal growth were higher in coral fragments (209.63 ± 43.65 % and 107.26 ± 25.75 %) than in loose coral colonies (124.33 ± 15.83 % and 99.98 ± 19.74 %) (F = 25.532, P < 0.05 and F = 32.135, P < 0.05, respectively). At the end of the experiment of the rainy season, survival diminished in both treatments, but it was higher in coral fragments in comparison with loose coral colonies (62.95 ± 42.78 % vs. 45.65 ± 53.19 %) (F = 5.272, P < 0.05, respectively, Fig. 4).
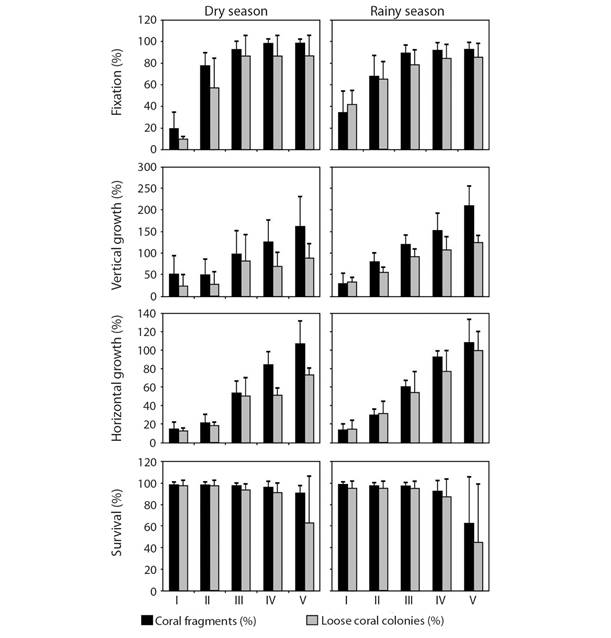
Fig. 4 Temporal variation of the fixation success, the vertical and horizontal growth, and survival of coral transplants recorded in the treatment of coral fragments (black columns) and loose coral colonies (gray columns) throughout 13 months and started during the rainy and dry seasons. Vertical lines represent the standard deviation. Taken from Nava & Figueroa-Camacho (2017).
Impact of the El Niño 2015-16 event: During the conclusion of the transplantation experiments, we witnessed the development of the El Niño event of 2015-16. This phenomenon reached a level of 2.9 on the Oceanic El Niño Index (ONI) and a lethal level of 8.5 of stress by warming in corals throughout 12 weeks (DHW), causing one of the strongest impacts of this phenomenon in the last decades (Fig. 5). As a result, corals in Zihuatanejo, Guerrero were exposed to thermal stress for almost 13 months. In June 2015, we recorded major changes in coral reefs as the presence of pale corals in almost all sites (up to 27.44 ± 35.23 %) with exception of Caleta de Chon. The presence of bleached corals was moderate at Playa Mazanillo, Playa Riscalillo and Playa las Gatas (up to 31.89 ± 20.89 %) but almost or totally absent at Islote Zacatoso and Caleta de Chon (up to 0.27 ± 0.46 %), although their coverage seemed to be similar among locations (F = 3.246, P = 0.08 and F =1.547, P = 0.276, respectively). In 2018, the cover of dead corals increased almost sixfold in Playa Riscalillo (59.17 ± 36.11 %) in comparison to its previous records (F = 6.666, P < 0.05, Table 2). The coverage of living corals was also reduced significatively through de time, especially in comparison to 2010 (24.42 ± 23.07 % vs. 77.23 ± 15.68 %) (F = 6.520, P < 0.05) and the significant change in the CMI confirmed that the conservation state at this locality in 2018 was the most reduced since 2010 (0.67 ± 0.35 vs. 0.10 ± 0.09) (F = 7.762, P < 0.05).
Fig. 5 Monthly variation of the Degree Heating Weeks (DHW) and the Oceanic El Niño Index (ONI) since January 2005 to May 2021 recorded at the sublittoral area of Zihuatanejo, Guerrero (modified from Nava et al. 2021).
Genetic diversity of zooxanthellae in coral populations of Zihuatanejo Guerrero: The results of the phylogenetic trees obtained from the two DNA regions (28S and ITS2) showed that 100 % of the samples corresponded to the genus Durusdinium (Fig. 6) and were closer to the subclades D1.1 and D1.2 (Fig. 7). The 28S molecular marker showed three haplotypes in each reef and the haplotype diversity values were h = 0.11 for Playa Riscalillo and Islote Zacatoso, h = 0.12 for Playa Las Gatas, and h = 0.14 for Caleta de Chon. The nucleotide diversity (π) varied between 0.00034 and 0.00044. The genetic diversity for the ITS2 molecular marker was higher than for the 28S molecular marker. Playa Las Gatas recorded three haplotypes and h = 0.16, Playa Riscalillo five haplotypes and h = 0.34, Caleta de Chon seven haplotypes and h = 0.29, and Islote Zacatoso 17 haplotypes and h = 0.61. Nucleotide diversity varied between 0.00107 and 0.02784.
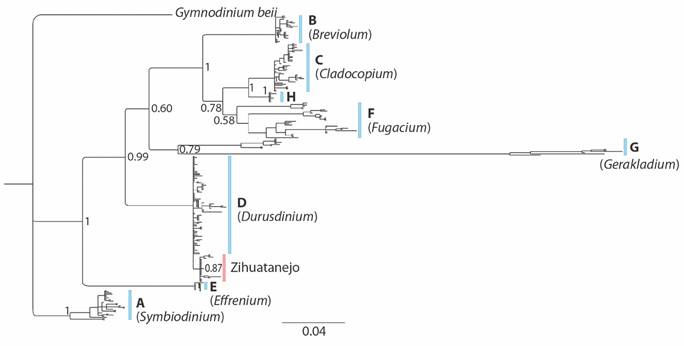
Fig. 6 Phylogenetic tree for the genus Durusdinium built with sequences of the molecular marker 28s obtained from GenBank and of zooxanthellae of the coral communities of Zihuatanejo, Guerrero (Islote Zacatoso, Playa las Gatas, Caleta de Chon, and Playa Riscalillo) during 2018 and 2019 (modified from Cárdenas-Alvarado et al. 2021).
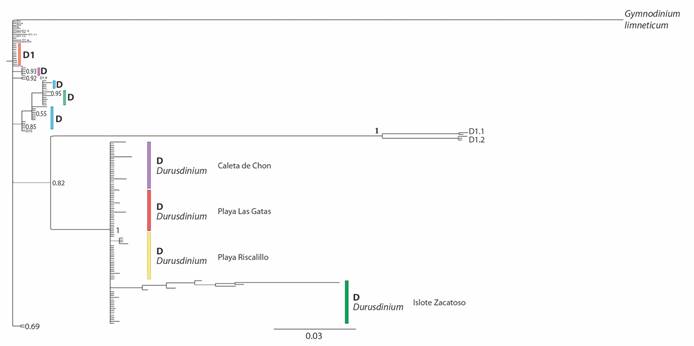
Fig. 7 Phylogenetic tree for the genus Durusdinium built with sequences of the molecular marker ITS2 obtained from GenBank and of zooxanthellae of the coral communities of Zihuatanejo, Guerrero (Islote Zacatoso, Playa las Gatas, Caleta de Chon, and Playa Riscalillo) during 2018 and 2019 (modified from Cárdenas-Alvarado et al. 2021).
Using local ecological knowledge as the starting point for the inclusion of citizen involvement: Local ecological knowledge is not limited to the recognition and identification of species. Like the case of the fish 'cocinero'(Caranx caballus), which, even though it is not a highly-priced species compared to other fishes in the area, it is well known and valued by locals inhabitants in Zihuatanejo for its frequent use as homemade food (Table 3). Drouet-Cruz (2020) explained that the knowledge around local biological resources is as rich as to determine and describe a) the species abundance, b) life cycles, including reproductive seasonality, c) behavior or species etiology, d) seasonal distribution, and finally, e) the way in that species in Zihuatanejo interact. Along with biological knowledge, local collaborators possess complementary observations linking coral ecology with physical and climatological phenomena. They use available local knowledge about ocean currents, the morphology of sites, and weather patterns to plan their daily activities and future scenarios, as well as to prevent environmental and productive contingencies. Specifically of knowledge about reefs, the inhabitants of Zihuatanejo sort their species into two main categories: a) corals and b) 'ripio'(gravel). For these people, corals are defined exclusively as those plants that 'seem like a black tree, with phosphorescent branches (Drouet-Cruz, 2020: 66, our emphasis). In other words, in Zihuatanejo, only arborescent corals, such as those in the genus Antipathes, are considered corals. Ripios or scleractinian corals, are more abundant in shallow areas. Also, they are extensive, with 20 or 30 m in length and are not conceptualized as corals in the minds of local fishers. In all cases, corals and ripios harbor numerous and diverse gilds of fishes; in the case of ripios, they are of great importance for local fishermen communities because many fishes of commercial importance spawn and refuge among them, including 'huachinango'and 'pargo'(belonging to the Lutjanidae and Sparidae families, respectively, Drouet-Cruz, 2020).
Table 3 Frequency (F), average rank (AR), and salience (S) of most salient commercially important species in Zihuatanejo according to a free listing exercise among fishers.
| Phylum | Family | Scientific name | Common local name | F | AR | S |
| Chordata | Lutjanidae | Lutjanus campechanus - Poey, 1860 | Huachinango | 84.4 | 3.56 | 0.684 |
| Carangidae | Carax caballus - Günther, 1896 | Cocinero | 72.9 | 5.63 | 0.482 | |
| Haemulidae | Hameulon spp. - Gill, 1862 | Ronco | 64.6 | 4.95 | 0.452 | |
| Lutjanidae | Lutjanus aratus - Günter, 1864 | Pargo | 68.8 | 5.92 | 0.411 | |
| Scombridae | Thunnus obesus - Lowe 1839 | Atún | 47.9 | 8.57 | 0.275 | |
| Carangidae | Selar crumenophtalmus - Bloch y Schneider, 1801 | Ojotón | 49 | 7.72 | 0.254 | |
| Coryphaenidae | Coryphaena hippurus - Linnaeus, 1758 | Dorado | 43.8 | 7.93 | 0.253 | |
| Istiophoridae | Istiompax indica - Cuvier, 1832 | Marlin | 42.7 | 9.39 | 0.204 | |
| Mollusca | Ostreidae | Crassostrea spp. | Ostión | 16.7 | 5.69 | 0.132 |
| Octopodidae | Octopus spp. | Pulpo | 16.7 | 7.13 | 0.123 | |
| Haliotidae | Haliotis spp. | Abulón | 5.2 | 17.6 | 0.03 | |
| Arthropoda | Palinuridae | Panulirus spp. | Langosta | 15.6 | 4.13 | 0.128 |
Discussion
The baseline study confirmed the relevance of several sites such as Playa Manzanillo, Playa Riscalillo and Islote Zacatoso because they harbored coral communities with a moderate-high conservation status, confirming their potential for conservation. The relative position to the coast of these sites also favors the dilution of the sewage contamination from Ixtapa and Zihuatanejo Bay. Moreover, the lack of access to Islote Zacatoso by land excludes some users, which means a less intensive activity impacting the site. In contrast, results evidenced that locations with coral communities inside the bay of Zihuatanejo (Playa las Gatas) or nearby to places subjected to land transformation (Caleta de Chon), were exposed to relevant anthropogenic pressure, visible as high concentration of chlorophyll or high sedimentation rates, attributed to the land transformation (Nava & Ramírez-Herrera, 2012). Nonetheless, all reefs support a high diversity of actors that have access to the resources. Playa Manzanillo and Playa las Gatas, more accessible by land and closer to the bay, are places with a diversity of types of fishing (e.g. recreational on boat, recreational on-shore, commercial, and commercial diving) and touristic activities coexisting.
The boring sponge assemblages present in the coral reefs were comparable to other assemblages distributed throughout the Mexican Pacific in terms of abundance and diversity (Carballo et al., 2019). The concentration of chlorophyll and stable isotopes of nitrogen and carbon (𝛿15N and 𝛿13C) coincided with the development of boring sponge assemblages in the reefs near the bay. Likewise, the results showed that the skeleton of living coral colonies was the least affected and confirmed that the calcareous substratum formed by dead corals was more vulnerable to bioerosion by boring sponges. Although the weakening and detaching of the reef framework free new spaces for colonization, it can cause a progressive degradation of the coral reef if bioerosion surpasses the reef's capability of recovery (Marlow et al., 2019; Nava et al., 2019). Because boring sponges are filter-feeding organisms, it has been suggested that the water enrichment with dissolved and particulate organic matter and nutrients can enhance sponge abundance and its bioerosion activity (Alvarado et al., 2017).
The increase of dead coral coverage attributed to environmental degradation and climate change have been considered as a cause of the rise in boring sponge abundance and bioerosion rate during the last decades (Schönberg et al., 2017), highlighting the importance of promoting a high cover of living corals in the reef to prevent the reef deterioration. For this purpose, the implementation of coral reef restoration strategies are useful (Nava et al., 2014) and we found that coral fragments were an excellent choice as coral transplants because they reach the fixation more quickly, and reach the highest survival and growth over the loose coral colonies. This could be explained by the initial stimulation of healing after breakage, which may favors the calcium carbonate deposition (Rempel et al., 2020).
Seasonality was a relevant factor during the implementation of our restoration plan, because the strike of storms and hurricanes could be prevented during the dry season, reducing transplant detaching and mortality. It is also important to highlight the relevance of the immobilization of coral transplants and the kind of substrate where they were transplanted (Casey et al., 2015; Oviedo, 2011). We learned that transplants should be attached to the natural substrate from the beginning, where they contribute permanently to the coverage of living corals in the reef framework. Several restoration initiatives that have reported low survival of coral transplants used artificial substrata that never will be part of the reef framework and natural materials that disintegrate or suffer deterioration very quickly (Cabaitan et al., 2015; Suzuki et al., 2011; Toh et al., 2017). The use of pita fiber ropes as a holding material and wood stakes as substrata (García et al., 1995) for example, resulted in a final survival of 0 %. On the other hand, when the same treatment is carried out with plastic ties, the percentage of survival was close to 67 %, increasing up to 80-95 % when adherent solutions are used (García et al., 1995; Ruiz et al., 2013; Tortolero-Langarica et al., 2014). Procedures that provide stability long enough to promote the integration of transplants in the reef framework, seem to be a key factor to the success of the restoration (Edwards & Gomez, 2007).
The approach tested in Playa las Gatas showed a high potential due to its low cost and the possibility to be implemented by non-specialized people. The steel grids were a reliable material to transplant corals in a damaged coral community because they endured the impact of the waves and currents until the fixation of coral transplants occurred. After several months, this material disintegrated, leaving the coral transplants fixed to the substratum. Nonetheless, as many experimental approaches, the experiment presented here requires the participation of the local communities to reach a significant outcome of coral reef restoration. As we observed during the end of the experiment, it is also important to prevent the future impacts of natural phenomena, such as the El Niño, that has the potential to revert the natural recovery of coral reefs or even the positive outcome of restoration initiatives (Nava et al., 2021). During the El Niño event of 2015, Islote Zacatoso and Caleta de Chon endured the thermal stress that caused bleaching and coral mortality at the other localities.
After the examination of the symbiotic relationship of corals with their algae symbionts, which can confer them thermal resistance (LaJeunesse et al., 2010; Wang et al., 2021), we found that the current presence of only one genus of Symbiodiniaceae in Zihuatanejo may be the resulting selection made by previous El Niño events in the region, that could promote the acquisition of resistant symbiotic algae to endure thermal stress (Baker et al., 2004; Stat et al., 2009; Van Oppen et al., 2005; Wang et al., 2021). The genetic diversity also could be related to the resistance to thermal stress, because sites such as Playa las Gatas and Playa Riscalillo, with the lowest genetic diversity, showed the highest records of coral bleaching and mortality during the El Niño event of 2015-16, being the opposite with Islote Zacatoso and Caleta de Chon (Cárdenas-Alvarado et al., 2021). In addition, the haplotypes with more mutational steps from the common haplotype corresponded to symbiotic algae from Islote Zacatoso which showed the highest genetic differentiation. This suggests that different haplotypes of Islote Zacatoso could be conserved and their genetic diversity contributed to the resistance to bleaching by thermal stress. It is important to assess the thermal resistance of corals under more controlled conditions focused on the entire coral holobiont (zooxanthellae and other associated microorganisms, such as bacteria, fungi, viruses, etc.) (Brener-Raffalli et al., 2022; Li et al., 2021; Van de Water et al., 2022).
To date, specialized experiments are being made in the laboratory under different levels of seawater temperature testing thermal resistance of corals from Islote Zacatoso, Caleta de Chon, and Playa Riscalillo. The identification of coral populations resistant to thermal stress is a key goal in this conservation effort in Zihuatanejo, Guerrero, 1) to detect coral reefs with high potential as source of transplants for coral communities impacted by El Niño events, and 2) to increase the possibilities of success of future initiatives of coral reef conservation in front of climate change. An additional lesson learned is the importance of the evolving of local inhabitants in coral reef restoration. Marine ethnobiological knowledge is the empirical understanding that coastal populations have acquired on the ecological properties (structure and function) of coastal and marine ecosystems that provide them with resources and aesthetic, cultural, and spiritual assets (Narchi et al., 2014). In Zihuatanejo, only a few efforts have accounted for this knowledge. One such effort conducted by Drouet-Cruz (2020), registered and analyzed the local ecological knowledge held by different groups of actors regarding their economic activity and closeness with the coral communities and associated species. A clear example is the capability that local inhabitants have for recognition and naming marine species. Through free-listing Drouet-Cruz (2020, and references therein) identifies 23 species belonging to five phyla (Table 3); four of them were animals a) Chordata, b) Mollusca, c) Arthropoda, d) Echinodermata and e) Chlorophyta (macroalgae).
As reported in other realms of ethnobiology (Berlin, 2014), as the management of species increases, different groups of actors tend to correctly identify these species with greater ease as the salience in Table 3 suggests. Conversely, as the management decreases, fewer people will be able to identify and relate to specific species. The easiness of species recognition is closely related to their overall importance, and not only to the commercial relevance of a species. At the same, time the complex and diverse constellation of actors convening in Zihuatanejo has allowed us to realize how different conceptualizations of resources and the expectations about the rights of use can hinder conservation efforts. For example, people making a living from recreational tourism, in collaboration with municipal government agencies intended to define conservation polygons. Various groups of local fishers have not taken those initiatives very well, because they felt their historical rights of use hadn't been included in these initiatives, and they also felt excluded from the circles where important decision-making is being taken. Therefore, these previous initiatives hadn't been successfully implemented since they have not had the citizen support that is required in projects of this nature. Zihuatanejo is an area with several groups of users with different motivations, and after a year of working with various stakeholder groups, we have realized that the citizen participation strategy requires deep grassroots work. It requires a great effort in environmental education. While some users have deep ecological knowledge about species diversity and abundances (Douet-Cruz, 2020), this does not exactly translate into direct empowerment while promoting conservation.
Coral reefs of Zihuatanejo are relevant ecosystems with a high potential for coral reef restoration, although the anthropogenic and natural sources of impact must be attended to in order to meet a significant level of restoration. The transplant of coral fragments is a reliable approach to recover coverage of living corals. To confront the increase in the use of coral reefs and the impact of climate change, we recommended the use of thermal resistant corals and to scale the restoration efforts, extending the projects to local stakeholders, joining the formal and local ecological knowledge.
Ethical statement: the authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgments section. A signed document has been filed in the journal archives.

