











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
Inselbergs are defined as isolated rocky outcrops forming small and discrete ecosystems scattered worldwide. Besides its cultural and economic importance, this kind of habitat is characterized by their singular biotas (Porembski & Barthloltt, 2012; Fitzsimons & Michael, 2017). Overall, the importance of these ecosystems for the conservation of biodiversity has been attributed to their unique microclimatic conditions regarding to their surrounding areas, higher habitat heterogeneity, and relative isolation, which locally increase the diversity of unaltered land cover (Porembski & Barthlott, 2012; Burke, 2003; Speziale & Ezcurra, 2015; Fitzsimons & Michael, 2017). In the case of bats, inselbergs increase availability and predictability of food (especially insects) and provide variety of roosts, being important sites for their conservation (Fredericksen, Fredericksen, Flores, Mcdonald, & Rumiz, 2003).
Bat diversity in Argentina comprises 67 species (Barquez, Díaz, Montani, & Pérez, 2020), with most species (~49) being distributed in the Northern portions of the country (Barquez, Díaz, & Ojeda, 2006). Accordingly, most of the available knowledge about these mammals is restricted to the forested areas of Northern Argentina, including the Yungas and the Paranense forests (Sánchez, 2016). However, more in general, our regional knowledge about bats is mostly historical and based on punctual records, with few studies focusing on entire assemblages (Sandoval, Sánchez, & Barquez, 2010; Barquez, Sánchez, & Sandoval, 2011; Sánchez, Carrizo, Giannini, & Barquez, 2012a; Sánchez, Giannini, & Barquez, 2012b; Idoeta, 2018).
The Paraje Tres Cerros (PTC), Corrientes Province, Argentina, is characterized by the presence of isolated rocky outcrops surrounded by a plain matrix of grasslands and wetlands. Several studies have pointed out their importance for the biodiversity conservation, highlighting the presence of species with different biogeographic affinities and some endemisms (Meregalli, 1998; Ravenna, 2003; Ravenna, 2009; Cajade et al., 2013a; Cajade, Etchepare, Falcione, Barrasso, & Álvarez, 2013b; Gervazoni, 2017; Fandiño, Fernandez, Thoman, Cajade, & Hernando, 2017; Ojanguren-Afilastro et al., 2017; Nadal, Achitte-Schmutzler, Zanone, Gonzalez, & Avalos, 2018). Previous contributions to the knowledge of the bat fauna of PTC include the first record of Eumops bonariensis for Corrientes Province and the mention of an unidentified species of Myotis that roosts under rocks (Idoeta, Cajade, Piñeiro, Acosta, & Pautasso, 2015; Cajade et al., 2013a). Based on these and other unpublished studies (Argoitia, 2016), this area was recognized as an AICOM (Spanish acronym for “Important area for the conservation of bats”) in March of 2016 (RELCOM, 2020).
In this study, we report and summarize new natural history data of the bat fauna of PTC. We discuss the importance of inselbergs and the surrounding matrix for conservation of tropical bat diversity at a regional level. In this way, we studied the richness of bats associated to different habitats (rocky outcrops vs. pediments) in relation to their trophic guilds and roosts. Finally, we highlight the need to conduct additional ecological studies on these poorly known habitats, with special focus on their conservation.
Materials and methods
Study area: The studied inselbergs are in PTC, San Martín Department, Corrientes Province, Central-Eastern Argentinian Mesopotamia. Mean annual temperatures are between 21.5 and 19.5 °C, varying from 16-13 °C in the coldest month (July) and 27-26 °C during the warmest (January). Accumulated annual precipitations varies between 1 500 and 1 000 mm (Carnevali, 1994). The relief in this area consists of three small hills of quartz sandstone of dune geo-morphology and trunked summits, oriented SE-NW along 10 km of extension (Herbst & Santa Cruz, 1999). Individual hills are locally known as “Cerro Nazareno” (179 m.a.s.l., 83 ha, 29º06’78” S & 56º55’19” W), “Cerro Chico” (148 m.a.s.l., 34 ha, 29º06’89” S & 56º55’87” W) and “Cerro Capará” (158 m.a.s.l., 79 ha, 29º09’55” S & 56º51’43” W) (Fig. 1). A geological interface of pediments of low slope is placed between the rocky outcrops and the flat matrix with a typical vegetation of the “Campos y Malezales” ecoregion (Burkart, Bárbaro, Sánchez, & Gómez, 1999). The rocky outcrops are characterized by hydrophilic forests at the Southern flanks and grasslands on the summits and the Northern sides, and they lack waterbodies. The pediments contain some small artificial ponds, and are principally covered by grasslands, with some small patches of hydrophilic forest (Parodi, 1943; Cajade et al., 2013a).

Fig. 1 A. Location of the isolated rocky outcrops of Paraje Tres Cerros (black star), Corrientes Province, Argentina. B and C: general satelital imagens of the hilly system, depicting the Cerros Nazareno (Nz) and Chico (Ch) in B and Capará (Cp) in C. References for symbols: ■ Rocky grasslands, ● Rocky forest patches, ▲ Pediment forest patches and ♦ Cutwaters.
Bat sampling: We did field trips in 2012 (one time in August and September and two times in November), 2013 (two times in February and one time in April), 2014 (two times in February, and one in March and July), 2015 (February to May, September and October), 2016 (February to May, July and September to December) at 13 sites distributed in two types of habitats: (i) the rocky outcrops (including three sites in grasslands and three in forest patches), and (ii) the pediment (including five forest patches and two cutwaters) (Fig. 1). Each site was explored twice (26 in total), during two consecutive nights, using five mist nets (four of 6 x 3 m and one of 12 x 3 m, with a size mesh of 12 x 12 mm), placed 40-50 m apart from each other. The total sampling effort was of 52 nights with 130 nets for the entire area and 104 hours of roosts search. Only days with occurrence of bats (= 30) were considered for the analyzes. Nets were activated in time bands between 18:00 and 01:00 h depending on seasons and revised for ~5 hours at intervals of 20 minutes. The placement of nets was carried out during the daylight hours, considering potential flight paths (e.g., edges and clearings of forests, rocky cliffs) or food resources (e.g., near to plants known to be consumed by bats; Kunz & Parsons, 2009). Surveys were conducted in each site during the night, except when there was a full moon (due to the lunar phobia of bats), rains or strong winds, to avoid biases in the capture of bats (Morrison, 1978). Captured bats were photographed and taxonomically identified in situ using specific keys (Barquez, Mares, & Braun 1999; Díaz, Solari, Aguirre, Aguiar, & Barquez 2016). Each captured specimen was placed in a cloth bag, then we took the following data: sex, relative age, reproductive condition, total length, tail length, hind foot length, ear length and forearm length (Díaz, Flores, & Barquez, 1998), and then they were released. All measurements were taken with a digital caliper to the nearest 0.5 mm. Animals were trapped, manipulated, and a sample was euthanized to confirm species identification, following the guidelines of the American Society of Mammalogists (Sikes, Gannon, & The Animal Care and Use Committee of the American Society of Mammalogists, 2011). Voucher specimens were deposited at the Museo Argentino de Ciencias Naturales “Bernardino Rivadavia” (MACN, Buenos Aires, Argentina), and the Colección de Mastozoología de la Facultad de Ciencias Exactas, Naturales y Agrimensura de la Universidad Nacional del Nordeste (CM-FaCENA, Corrientes, Argentina) (Digital Appendix 1).
Species richness: We inventoried and described the bat species richness (S) of PTC, including both the rocky outcrops and their pediments. A species accumulation curve was made to evaluate the representativeness of the inventory. The sampling effort was considered as the total number of days with occurrence data for pediments and rocky environments. We built a matrix of presence/absence (Digital Appendix 2), calculating the completeness of the inventory through a non-parametric estimator (Jacknife 1; first-order Jackknife richness) for incidence data. This estimator uses the number of species occurring in once sample, and is efficient with low sampling efforts (Moreno, 2001). Samples were randomized 1 000 times to eliminate the influence of the order in which the data was taken (Colwell & Coddington, 1994). The degree of completeness of the inventory was obtained as the percentage of species observed in relation to the number of species predicted by the richness estimator. Bat assemblages were characterized using the relative frequency (F), as the occurrence of each species in the area regarding to the total number of captures expressed as percentage. The similarity in the composition of species between two types of habitats (rocky outcrops vs. pediments) was compared by a Non-Metric Multidimensional Scaling (NMDS), using Jaccard index as a measure of similarity, based on incidence data matrix. The distortion of the resolution of the two-dimensional arrangement is represented by a stress value. The difference between the richness values for the studied habitats was tested with an ANOSIM test. In all analyses, the significance level was of α = 0.05. All analyses were made using the R Version 3.6.2, packages: BiodivesityR, ggplot2, vegan (R Development Core Team, 2019).
Bat ecology: Trophic guilds were assigned following the criterion proposed by Sánchez et al., (2012a) and Sánchez (2016), based on the species diet, foraging habitat and type of flight of each taxon. The conservation status of each species was taken from the assessment of extinction risk of Mammals from Argentina (SAyDS-SAREM, 2019). Roost searches were carried out randomly at each site during the day, lasting two hours before to the placement of nets. We reviewed all those possible places in which bats usually rest, such as trees, crevices between or under rocks, and human constructions. Roosts in trees were identified and classified according to the keys proposed by Andrews (2013). Individuals in each roost were taxonomically identified and counted whenever possible. In the case of Myotis sp., given the large number of found specimens, we also recorded the seasonal patterns of the use of the refuges between or under rocks through walks in each rocky outcrops of two hours on each field trip (N = 8), from August 2013 to August 2014. We marked each checked rock with paper tape to avoid recounting.
Results
Species richness: Thirteen species were recorded, including three families (Molossidae, Phyllostomidae, and Vespertilionidae) and seven genera (Eptesicus, Eumops, Lasiurus, Molossops, Molossus, Myotis, and Sturnira). The relative frequency (F) for most species in the area varied between 2.3 to 14.8 % (N = 10); the highest F corresponded to Eptesicus furinalis, while Lasiurus blossevillii and Molossus rufus were recorded only once (Table 1). All specimens were determined to species level, except for a sample of individuals of a distinctive morphotype of Myotis that were identified to the genus level, which is currently under study. According to the estimator Jacknife 1, inventory completeness reached high levels of 80 % (Obs = 13/Exp =17, with a 1.4 of standard deviation). An accumulation curve of softened species for our entire dataset (i.e., clustering all sites) seemed to asymptote, indicating that a large percentage of the regional bat richness was sampled (Fig. 2).
Table 1 Species and trophic guilds of the bats of the Paraje Tres Cerros, Corrientes province, Argentina
| Code | Taxa | Nazareno | Chico | Capará | Pediment | F | Trophic guild | ||||
| RG | RFP | RG | RFP | RG | RFP | FP | C | ||||
| Phyllostomidae | |||||||||||
| Sli | Sturnira lilium | + | + | + | + | 14.8 | FHCGU | ||||
| Vespertilionidae | |||||||||||
| Mal | Myotis albescens | + | + | + | + | 5.7 | IBCT | ||||
| Msp | Myotis sp. | + | + | + | + | + | + | 12.5 | IBCA | ||
| Mri | Myotis riparius | + | + | + | + | 13.6 | IHCA | ||||
| Mrb | Myotis ruber | + | + | 4.5 | IHCA | ||||||
| Leg | Lasiurus ega | + | 2.3 | IBUnA | |||||||
| Lbl | Lasiurus blossevillii | + | 1.1 | IBUnA | |||||||
| Efu | Eptesicus furinalis | + | + | + | + | + | 17 | IHCA | |||
| Molossidae | |||||||||||
| Ebo | Eumops bonariensis | + | + | 4.5 | IBUnA | ||||||
| Epa | Eumops patagonicus | + | + | + | 5.7 | IBUnA | |||||
| Mte | Molossops temminckii | + | + | 12.5 | IBCA | ||||||
| Mmo | Molossus molossus | + | + | 4.5 | IBUnA | ||||||
| Mru | Molossus rufus | + | 1.1 | IBUnA | |||||||
Reference: RG, rocky grassland; RFP, rocky forest patches; FP, forest patches; C, cutwaters; FHCGU, highly cluttered space/gleaning/understory frugivore; IBCT, background-cluttered space/trawling insectivore; IBCA, background-cluttered space/aerial insectivore; IBUnA, uncluttered space/aerial insectivore; IHCA, highly cluttered space/aerial insectivore and F, relative frequency. + = presence.

Fig. 2 Bat species accumulation curve for Paraje Tres Cerros, Corrientes Province, Argentina. The gray area corresponds to the standard deviation.
Regarding the composition of bat assemblages based in two NMDS axes, the arrangement of species in the bi-dimensional space shows a difference in relation to compared habitats (pediments vs. rocky outcrops). Low values of stress (0.5) and R2 (0.9) show a high adjustment to the explained proportion of the variation (Fig. 3). The ANOSIM analysis did not express significant differences between the composition of both habitats (R = 0.2; P = 0.06).

Fig. 3 Composition of the Non-Metric Dimensional Scaling (NMDS) species from the bat assemblage of the Paraje Tres Cerros, Corrientes Province, Argentina. The plot represents two habitats, rocky outcrops, and pediment, in eight sites of the study area. References are as follow: (1) gray light labels are the sites names; NzRFP, Nazareno rocky forest patches; ChRFP, Chico rocky forest patches; CpRFP, Capará rocky forest patches; NzRG, Nazareno rocky grasslands; ChRG, Chico rocky grasslands; CpRG, Capará rocky grasslands; PPF, Pediment forest patches (5); C, Cutwaters; (2) black labels correspond to the species (see code on Table 1). Ellipses represent 70% confidence intervals of centroids for rocky outcrops and pediments.
Bat ecology: We registered five trophic guilds represented differently in each environment. Insectivorous bats were represented by one background-cluttered space/trawling species (IBCT), two background-cluttered space/aerial species (IBCA), three highly cluttered space/aerial species (IHCA) and six uncluttered space/aerial species (IBUnA), while the only frugivore was represented by a highly cluttered space/gleaning/understory (FHCGU) species (Fig. 4).

Fig. 4 Guild structure assemblage for the bats from Paraje Tres Cerros, Corrientes Province, Argentina. Relative frequency of each guild per site are depicted. Reference: IBCA, background-cluttered space/aerial insectivore; IBCT, background-cluttered space/trawling insectivore; IBUnA, uncluttered space/aerial insectivore; IHCA, highly cluttered space/aerial insectivore; FHCGU, highly cluttered space/gleaning/understory frugivore.
We registered nine types of refuges, five of them in trees trunks (3 in pediment and 2 in rocky outcrops), one on palm leaf (in pediment), two on rocks (in rocky outcrops) and one in a human construction (rocky forest patches) for eight species (Table 2).
Table 2 Identified roosts for bats of Paraje Tres Cerros, Corrientes Province, Argentina
| Roosts | Taxa | N | Sites | Observations |
| Tree trunks | Sturnira lilium | 8 | CpRFP | Adults in knot-hole and tear-out of Ocotea puberula |
| 5 | Adults in trunks of unidentified Myrtaceae | |||
| Tree trunks | Myotis riparius | 6 | PFP | Juveniles and adults between aerial roots of Ficus luschnathiana |
| Tree trunks | Molossops temminckii | UN | PFP | In desiccation-fissures unidentified dead tree |
| Tree trunks | Molossus molossus | UN | PFP | Tear-out unidentified tree |
| Palm leaf | Lasiurus ega | 4 | PFP | Adults in Washingtonia sp. |
| Rock crevices | Myotis sp. | 4 | NzRG | Northern slope vertical and horizontal ~150 m of height |
| Molossops temminckii | 1 | ChRG | Northern slope vertical ~130 m of height | |
| Lasiurus blossevillii | 1 | |||
| Under rock | Myotis sp. | * | NzRG, ChRG and CpRG | Adults solitary and in group |
| Human construction | Eumops patagonicus Myotis sp. | 3 ~20 | ChRFP | Adults in a mix colony |
References: NzRFP, Nazareno rocky forest patches; ChRFP, Chico rocky forest patches; CpRFP, Capará rocky forest patches; NzRG, Nazareno rocky grasslands; ChRG, Chico rocky grasslands; CpRG, Capará rocky grasslands; PPF, Pediment forest patches; C, Cutwaters. *See figure 5 for specific information.
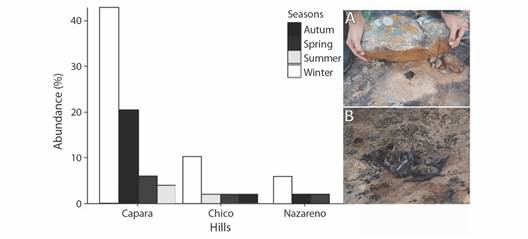
Two types of roosts were alternatively used by Myotis sp. in rocky microhabitats: (i) within horizontal and vertical crevices between larger rocks, and (ii) under rocks. In this case, each rock provides refuge by presence of small spaces formed between them and large rock blocks (rock terraces) near to cliffs or towards flanks of the rocky outcrops. In some cases (not counted), Myotis sp. shared these roosts with arachnids, blatids and lizards. We recorded 17 occurrences for 168 revised rocks and a total of 49 individuals (6 in Cerro Nazareno, 24 in Cerro Capará, and 16 in Cerro Chico). During the cold seasons, the occurrence of individuals between or under rocks was higher, with groups varying between 3 to 18 specimens. Contrarily, only solitary individuals were recorded during the warm seasons (Fig. 5).
Discussion
The Northern portion of Argentina constitutes the Southernmost limit for several species of tropical to subtropical distribution (Gardner, 2007). At least 37 species were documented for the ecoregion of “Campos y Malezales,” belonging to four families (Noctilionidae, Phyllostomidae, Vespertilionidae and Molossidae) (Idoeta, 2018). In this study, 13 species were recorded, representing 35 % of regional chiropterofauna. Other localities sampled in this ecoregion have less than 7-6 species in bat assemblages (59; Idoeta, 2018); only three localities placed in an ecotonal area with the Interior Atlantic Forests, Misiones Province, reach a richness of 15 species. Overall, the “Campos y Malezales” ecoregion is dominated by phyllostomids bats -with Artibeus lituratus and Sturnira lilium as most abundant species-, followed by molossids and vespertilionids (Idoeta, 2018), while in PTC molossids and vespertilionids are better represented than phyllostomids, being Eptesicus furinalis the most frequent species. Further studies, including the use of acoustic monitoring could increase the number of recorded species, especially for those taxa that avoid traditional samplings (i.e., mist nets).
The PTC bat fauna includes species from different trophic guilds, but with a dominance of insectivorous species (i.e., Vespertilionidae and Molossidae). This could be explained by the microclimatic conditions of the rocky outcrop ecosystems that favors the availability of arthropods (Fredericksen et al., 2003). According to Fredericksen et al. (2003), rocks capture heat during day, releasing it gradually during the night, and attracting numerous flying insects.
Therefore, both molossids and vespertilionids bats roost preferably near or in rock crevices (Kunz, 1982). Background-cluttered species (i.e., Myotis riparius, Myotis ruber, Eptesicus furinalis) were frequent in edge forest patches, while highly cluttered species (i.e., Myotis sp., Molossops temminckii) were most commonly caught in closed vegetation. Finally, uncluttered species (i.e., Lasiurus ega, Lasiurus blossevillii, Eumops bonariensis, Eumops patagonicus, Molossus molossus, Molossus rufus) were mostly found in open rocky grasslands. Myotis albescens, a background-cluttered space/trawling insectivorous, was mostly captured in cutwater, but also in forest patches. The only frugivore understory bat species, Sturnira lilium, was caught in a wide variety of sites within PTC.
The rocky outcrops of PTC provide a wide array of refuges, both in rocky and pediment environments for at least eight species (i.e., Eumops patagonicus, Lasiurus blossevillii, Lasiurus ega, Molossops temminckii, Molossus molossus, Myotis riparius, Myotis sp., Sturnira lilium). Overall, the recording roost sites are in accordance with the available data in the literature (Barquez, et al., 2020). The use of rock roosts (mostly by Myotis sp.), specially at Cerro Capará, is perhaps related to the availability of more suitable rocks at this place (i.e., quadrangular in outline and with rounded edges, with a mean surface of 107.5 cm2 and the entrances orientated to sunset direction). However, we do not rule out the possibility that other factors may influence the use of rocks at Cerro Capará, such as the absence of predators, or the availability of food resources. Although no specific studies of the microclimatic conditions of these refuges have been carried out so far, we think that the conglomeration of individuals of Myotis sp. in the colder seasons would be a behavior to deal with low temperatures, as was documented for other bat species that inhabits in temperate to cold areas (Ransome, 1968; Raesly & Gates, 1987; Herreid, 1967; Lewis, 1996). This and other aspects of the social behavior of this species should be studied in depth to explain the observed patterns in the use of roosts.
Both Lasiurus blossevilli and Lasiurus ega were documented only by individuals found in their roosts. This fact shows the importance of adding this kind of sampling to produce complete bat inventories.
The isolated condition of most inselbergs has a fundamental role as an evolutionary force over their biota, reflected in the presence of endemisms, microhabitat specialized species, or the evolution of singular social behaviors (Mares, 1997; Speziale & Ezcurra, 2012; Cajade et al., 2013b; Fitzsimons & Michael, 2017). For example, Myotis sp. developed the ability to take flight from the ground rocks instead from a free fall, using sometimes a quadruped locomotion to reach their shelters (Cajade et al., 2013a). Similar behaviors were developed by other rocky dwelling bats, such as Molossops mattogrossensis, Myotis vivesi and Platymops setiger (Blood & Clark, 1998; Happold, 2013).
At a local and regional level, rocky outcrops lead to an increase of mammal diversity compared to the surrounding non-rocky habitat (Mares, 1997). The richness of the assemblage and their different ecological aspects observed, seen among the bats of PTC is an indicator of the importance of inselbergs for the regional conservation of these mammals. Although the NMDS shows differences in the composition of the bat sets in both environments, the ANOSIM test indicates that these differences are not significant; this highlights the importance of both rocky outcrops and surrounding landscape on the pediments for the bat assemblage in the area. This situation is due that some species are exclusively found in one of the environments, Lasiurus ega and Molossus rufus in pediments and Lasiurus blossevillii in rocky outcrops. Also, the importance of the forest patches is that they offer shelter to the only species considered as threatened within the area (i.e., Myotis ruber, a little-known bat in this region, categorized as Near Threatened (NT) in Argentina) (Lutz, Díaz, Giménez, Sánchez, & Sandoval, 2019).
Overall, our findings highlight the importance and need to continue protecting inselbergs ecosystems, avoiding those anthropic activities of great impact (e.g., large-scale agricultural activities, intensive tourism) (Kunz, 1982; Turner, 1996). The conservation of isolated rocky outcrops will not only maintain a high regional richness of bat species, but also their functional importance for the maintenance of ecological processes (Kunz, de Torrez, Bauer, Lobova, & Fleming, 2011). We observe that both the rocky outcrops and the surrounding matrix are important for the local chiropterofauna, as both provide foraging areas and roost sites.
Ethical statement: authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.

