











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
The taphonomic attributes of a faunal assemblage provide information about which agents affect both the distribution and preservation of ancient or newly formed biogenic materials during their depositional and post-depositional processes. These processes are regulated by the surrounding environmental conditions and the life habits of the organisms that constitute such assemblages (Callender et al., 2002; El-Sorogy, 2015; Gómez-Espinosa, Gío-Argáez, Farinati, Aliotta, & Salgado-Souto, 2018). Thus, actuotaphonomy is a valuable tool for reconstructing fossil communities because it establishes analogies between observable processes in the present and those that happened in the past. Actuotaphonomy helps elucidate the events that lead to preservation of organic remains, which in turn assists in interpretation of sedimentological and ecological conditions that gave rise to the formation of a fossil deposit (Gómez-Espinosa & Gío-Argáez, 2009).
Kowalewski, Flessa, and Hallman (1995) proposed a triple classification graphical approach to taphonomic interpretations, using a diagram called “ternary taphograms,” in which a taphonomic signature is assigned to a specific degree of alteration. With this classification assigned according to the percentage of shell coverage, it can be “good” (extensive or abundant coverage), “medium” (moderate coverage), or “poor” (little or no coverage).
Mollusks are major components of many current and fossilized marine ecosystems, and thus have been frequently incorporated into taphonomic analyses, with a special focus on gastropods and bivalves (Gómez-Espinosa et al., 2018). The in situ study of preserved mollusk assemblages is essential for understanding the taphonomic processes involved in the preservation or destruction of shell remains before their burial (Erthal, Korsian, & Simoes, 2013). For this reason, actuotaphonomic studies are particularly useful for predicting and estimating the physiochemical and/or biological processes that prevailed in a given environment, as well as speculating about the capability preserving mollusk assemblages in each environment (Kowalewski & Labarbera, 2004).
Different characteristic conditions may lead to taphonomic losses that affect the preservation of remains in a taphonomically active zone (TAZ), where the greatest dissolution, destruction, or taphonomic alteration of shells occurs. Among these conditions are the permanence of the organisms on the surface, the energy of the system, currents, salinity, and anthropogenic activity (Parsons-Hubbard et al., 1999; Olszewski, 2004; Gordillo, Bayer, Boretto, & Charó, 2014).
The objective of the present study was to analyze the different degrees of taphonomic alteration that might enhance either the preservation or destruction of a gastropod and bivalve assemblage in the present marine marginal depositional environment of the Laguna de Mandinga, Veracruz, Mexico. By characterizing the fragmentation, bioerosion, and encrustation on both gastropod and bivalve shells, along with the subsequent construction of ternary taphograms for each of these taphonomic attributes, we generated an actuotaphonomic model that may be applicable in analogous ecosystems.
Materials and methods
Study site: Laguna de Mandinga is located in the state of Veracruz, Mexico (19º00’00”-19º06’00” N & 96º02’00”- 96º06’00” W), 18 km South of Veracruz Port (Fig. 1). One of the main coastal lagoons along the Gulf of Mexico (Krutak, 1971; Reguero & García-Cubas, 1994), Laguna de Mandinga is a sedimentary environment of the Quaternary made of a fluvial accumulation of fine to very fine particle size, consisting of sand, silt, and clay overlying sandy and alluvial sediments. In some areas, it emerges with the gravel derived from the accumulation of mainly molluscan shells (Contreras, 1985). The fact that Laguna de Mandinga generates high concentrations of organic remains makes it a suitable location for taphonomic studies. The samples for this study were collected in a shell-remain accumulation called “El Conchal” located in “Laguna Redonda”.
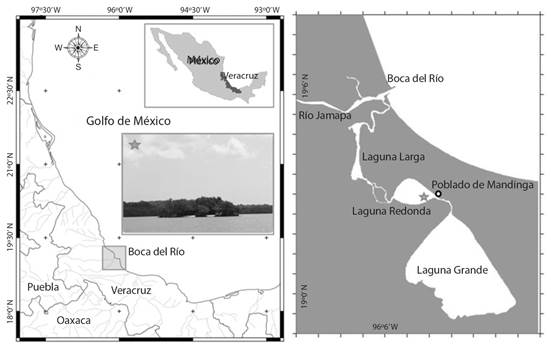
Fig. 1 Location of the study area “Laguna de Mandinga” Veracruz, close up mark with a star “El Conchal” shell remain accumulation.
At present, Mandinga is recognized as a tourist location with the development of commercial fishing. This status has meant that the lagoon and the channels that connect it to the Jamapa River have been regularly dredged since 1980 (Heimo, Siemens, & Hebda, 2004).
Sediment sampling and processing: A total of 1 697.9 g of sediment was collected from the ends and core from a shell mound known locally as “El Conchal”, following the sieving method described by Anderson, McBride, Taylor, and Byrnes (1998). The sample was processed in the Laboratory of Environmental Micropaleontology, ICMyL, UNAM. Only complete specimens were included in the present taphonomic analysis, defined as retention of more than 70 % of the original shell or valve surface. Among those with a smaller preservation percentage, we retained gastropod specimens that still had a last whorl and/or columella and bivalves that still had a hinge. A total of 1 165 specimens were evaluated, of which 173 were gastropods and 922 were bivalves.
Taxonomic identification, taphonomic attributes, and ternary taphograms: All bivalves and gastropods sampled, along with bioeroding and encrusting organisms associated with them, were taxonomically identified according to the illustrated catalogs of García-Cubas and Reguero (2004, 2007) for bivalve mollusks and gastropods from the Gulf of Mexico and the Caribbean Sea. The identification of ichnogenus and encrusting was carried out following the ethological classification of Seilacher (1964) and the World Register of Marine Species platform (http://www.marinespecies.org/). The compared taphonomic attributes considered were fragmentation, bioerosion, and encrustation, assessed using ternary diagrams based on Kowalewski, Flessa and Hallman (1995).
Results
Taxonomic identification: From the analyzed sample (N = 1 165), we identified five genera of bivalves (N = 922): Anadara Gray, 1847; Crassostrea Sacco, 1897; Ischadium Jukes-Browne, 1905; Mulinia Gray, 1837, and Rangia Desmoulins, 1832. We also identified four gastropod genera (N = 173): Bailya Smith, 1944; Cerithideopsis Thiele, 1929; Nassarius Duméril, 1805; and Neritina Lamarck, 1816 (Fig. 2). Among the bioeroding organisms found in the sample, three types of perforations were identified related respectively to ichnogenus Entobia Bronn, 1937; Caulostrepsis Clarke, 1908; and Oichnus Bromley, 1981. The specimens presented either one or more types of perforations per individual.
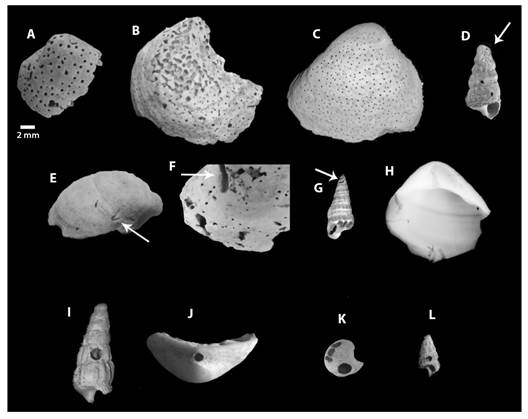
Fig. 2 Identification of bioerosive traces. A-D. Entobia perforations. E-F. Caulostrepsis perforations, I-L. Oichnus perforations.
Ethological classification:
DomichniaSeilacher, 1964
Ichnogenus Entobia Bronn, 1837
Type ichnospecies Entobia cretacea
Portlock, 1843
Description: Most of the perforations were small, rounded, and widely distributed (Fig. 2A, Fig. 2B, Fig. 2C, Fig. 2D). Some perforations were so strongly marked that they caused shells to lose their shape because of fragmentation. Perforation was more commonly found in bivalves than in gastropods, possibly because of the greater surface area of bivalves.
Ichnogenus Caulostrepsis Clarke, 1908
Type ichnospecies Caulostrepsis taeniola Clarke, 1908
Description: This ichnogenus was the least common in the sample. It was observed only in the outer layers of bivalves, exhibiting a wider diameter at the beginning of the perforation than at the end. C. taeniola was found in the gastropod shell towards the apex, manifesting as shallow elongated perforations (Fig. 2E, Fig. 2F, Fig. 2G, Fig. 2H).
Ethological classification:
PraedichniaSeilacher, 1964
Ichnogenus Oichnus Bromley, 1981
Type ichnospecies Oichnus simplex
Bromley, 1981
Description: This perforation was the second most abundant type in the sample, appearing in both bivalves and gastropods, although more commonly in bivalves. Perforations were frequently located at the umbo of bivalves and at the last whorl of gastropods but were seen towards the apical region of some specimens (Fig. 2I, Fig. 2J, Fig. 2K, Fig. 2L).
Regarding the encrusting specimens detected, we identified calcareous serpulid polychaete tubes and calcareous bryozoan and barnacle skeletons. The polychaete tubes were the most common of the three. Serpulid polychaetes were found in both the dorsal and ventral surfaces of the bivalve shells analyzed (Fig. 3A, Fig. 3B, Fig. 3C, Fig. 3D), as well as the external surface of gastropods (Fig. 3C).
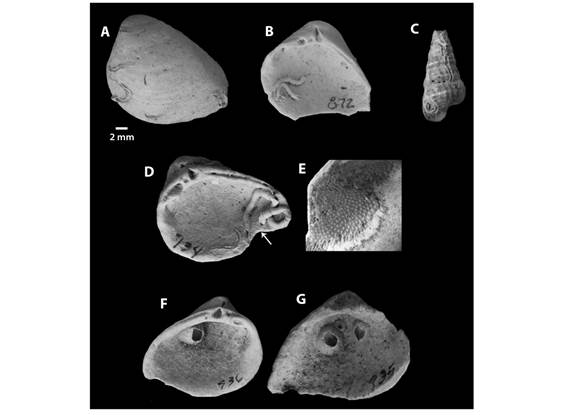
Fig. 3 Presence of encrusting organisms on mollusk shells. A-D. Serpulid polychaetes. D-E, G. Bryozoan colony. F-G. Barnacles.
Bryozoans from the Order Cheilostomatida Nusk, 1852 were identified in four bivalves in the sample, either on the ventral surface of the valves or on top of other encrusting serpulids and barnacles (Fig. 3D, Fig. 3E, Fig. 3G). Barnacles of the genus Amphibalanus Pitombo, 2004 were recognized on the ventral surface of two bivalve specimens, at the umbo region of this location, and it is inferred that encrustation happened postmortem (Fig. 3F, Fig. 3G).
Taphonomic attributes and ternary taphograms: Table 1 represents the taphonomic attribute percentage according to the coverage degree of the shells, based on the number of specimens observed in each sample.
Table 1 Total number of specimens and taphonomic attributes analyzed (fragmentation, bioerosion, and encrustation)
| Bivalves (N = 992) | Gastropods (N = 173) | |||||
| Taphonomic attribute | Poor (Grade 2) | Regular (Grade 1) | Good (Grade 0) | Poor (Grade 2) | Regular (Grade 1) | Good (Grade 0) |
| Fragmentation | 416 (41.9 %) | 489 (49.3 %) | 87 (8.8 %) | 100 (57.8 %) | 57 (32.9 %) | 16 (9.3 %) |
| Bioerosion | 526 (53 %) | 440 (44.3 %) | 26 (2.7 %) | 90 (52 %) | 65 (37.6 %) | 18 (10.4 %) |
| Encrustation | 942 (9 %) | 48 (4.8 %) | 2 (0.20 %) | 172 (99.4 %) | 1 (0.06 %) | - |
Fragmentation was observed in 58.1 % of the total sample; bivalves were the most affected group. While bioerosion appears in a great number (47 %) of mollusk specimens analyzed and encrustation is quite scarce (5 %) in the sample.
Analysis of the results of ternary taphograms (Fig. 4) for bivalves showed medium-grade (< 50 % shell coverage) fragmentation and bioerosion (Fig. 4A, Fig. 4B), according to Kowalewski, Flessa and Hallman (1995). The gastropod fragmentation also was medium grade (Fig. 4A), but the bioerosion was poor (Fig. 4B). For both groups, encrustation (Fig. 4C) was poor (absent attribute).
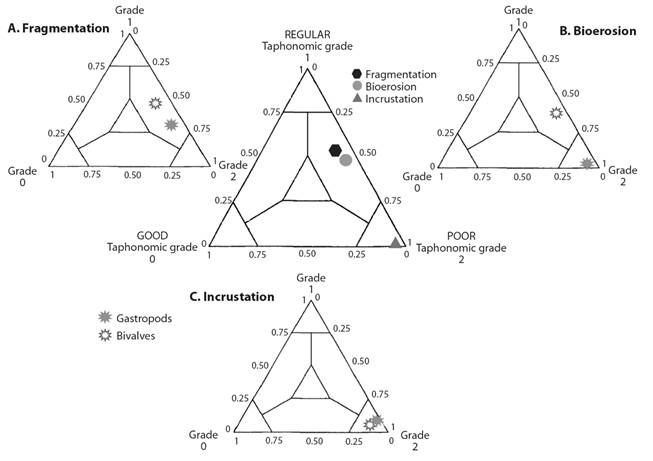
Fig. 4 Ternary taphograms. Global representation of the taphonomic attributes analyzed. Fragmentation and bioerosion exhibit a regular degree, while encrusting presents a poor degree. A. Bivalve and gastropod fragmentation, regular. B. Bivalve bioerosion, regular; gastropod bioerosion, poor. C. Bivalve and gastropod encrustation, poor.
Among bivalves, fragmentation was present in 58.1 %, bioerosion in 47 %, and encrustation in 5 %. Within the gastropods, fragmentation was present in 42.2 %, bioerosion in 48 %, and encrustation in 0.6 %. Of the total, fragmentation was present in 55.7 %, bioerosion in 47.1 %, and encrustation in 4.4 %.
Based on the taphonomic analysis, bivalves were the most affected group (Table 2).
Table 2 Organisms exhibiting bioerosion and encrustation according to their taphonomic degree and taxonomic group
| Bivalves | Gastropods | |||||
| Poor | Regular | Good | Poor | Regular | Good | |
| Bioeroders | ||||||
| Entobia | 181 | 17 | 28 | 12 | ||
| Caulostrepsis | 83 | 7 | ||||
| Oichnus | 109 | 1 | 18 | |||
| Entobia/Caulostrepsis | 29 | 5 | 2 | |||
| Entobia/Oichnus | 35 | 3 | 1 | |||
| Caulostrepsis/Oichnus | 2 | 11 | 4 | |||
| Entobia/Caulostrepsis/Oichnus | 1 | |||||
| Encrusters | ||||||
| Barnacles | 1 | |||||
| Bryozoans | 3 | |||||
| Serpulids | 43 | 1 | 1 | |||
| Barnacles/Bryozoans | 1 | |||||
| Serpulids/Bryozoans | 1 | |||||
Poor (Grade 2) = absent attribute; regular (Grade 1) = < 50 % coverage of the shell; good (Grade 0) = > 50 % coverage of the shell.
Discussion
Modern marine environments offer information about past geological processes and conditions that affected organisms and their eventual burial. However, the fossil record in brackish water bodies, such as coastal lagoons, has a lower diversity of representation compared to the marine record because of how organisms are preserved and the ephemeral nature of the habitats (Adomat, Gishler & Oschmann, 2016). For this reason, actuotaphonomic studies in brackish water bodies are relevant to understanding the past extrinsic and intrinsic conditions of these ecosystems.
Different factors regulate the abundance and composition of fossil deposits, such as susceptibility to modification and destruction by organisms, taphonomic conditions exposed in the environment, and the time scale accumulation (Behrensmeyer & Kidwell, 1985). These factors are clearly observable in current marine-marginal environments, where ecological interpretations of the species can be made based on the factors that influence the transport of the organic remains.
In order to study a recent mollusk assemblage in a marine marginal environment, just as the ones that are developed in coastal lagoons, requires consideration of the energy rates, environmental interactions, and trophic relationships of the system under study. These factors regulate molluscan communities and are crucial tools in interpreting their taphonomic attributes.
Taphonomic interpretation: The sample position in each ternary taphogram expresses the variation of taphonomic characteristics, which may result from the different conservation environments (hydrodynamic regime, biological activity, degree of subaerial exposure), variation in size, and architecture of the shells (Kowalewski, Flessa, & Hallman 1995). The use of taphograms saves space and allows a quick comparison of the taphonomic characteristics among samples.
Fragmentation was observed in 58.1 % of the total sample, and bivalves were the most affected group. This characteristic is associated not only with high environmental energy but also with biological interactions. The origin of the fragments could depend on different stages of taphonomic processes (depredation, bioturbation, and biostratinomic processes), system energy, and anthropogenic action (opening of canals) that provokes constant cycles of rework and exhumation of organic remains (Kidwell, Fürsich, & Aigner, 1986; Zuschin, Stachowitsch, & Stanton, 2003).
Because of the nature of the encrusting organisms and bioeroders, it is believed that they developed in underwater conditions, so that the shells were deposited and accumulated in islets called “conchales”. Bioerosion was present in a great number (47 %) of mollusk specimens collected from Laguna de Mandinga, but encrustation (5 %) was quite scarce. Such scarcity might arise from the considerable sediment reworking and exhumation that characterize the study site. Both the burial and the frequent subaerial exposure may have protected the shells from bioeroders and encrusting organisms (Lescinsky, Edinger, & Risk, 2002).
Most of the organisms responsible for the perforations identified in our sample were clionid sponges (Porifera), which produce the ichnogenus Entobia. To a lower degree, there were carnivorous gastropods that produce the ichnotrace Oichnus and the serpulid polychaetes that produce the ichnogenus Caulostrepsis. All three ichnogenera were present in both bivalves and gastropods, but their incidence was higher in bivalves. The same ichnofossils and their higher prevalence on bivalves were registered on recent shell assemblages from a siliciclastic beach on the Gulf of Mexico (Gómez-Espinosa et al., 2018).
Given the relatively high percentage of bioerosion detected in the Laguna de Mandinga sample (47 %), it can be inferred that a great number of the analyzed organisms spent considerable time in the TAZ, where bioeroding organisms could settle on the exposed surface of the mollusk shells (Davies, Powell, & Stanton, 1989; Walker, Parson-Hubard, Powell, & Brett, 1998).
Encrusting organisms were found in only 5 % of the specimens, and all but one individual were bivalves. Most encrustations were located on the ventral surface of the valves, suggesting a postmortem occupation, most likely to gain protection (McKinney, 1996). A large number of encrusting bryozoans were observed colonizing the surface of other encrusting organisms (i.e., serpulid polychaetes and barnacles), so that the first arrivals could be considered as secondary colonizers capable of inhabiting surfaces with varying degrees of roughness and/or curvature (Gherardi & Bosence, 1999; Gibson, 1992; Taylor & Wilson, 2003). The encrusting bryozoans did not have a preference regarding substrate specificity, and differences were related to the variable physical stability of the shells exhibiting different frequencies of circumrotatory growth (Jackson, 1984; McKinney, 1996).
The high rate of fragmentation and low degree of biogenic processes calculated for Laguna de Mandinga imply that the taphonomic processes occurring in mollusk assemblages are primarily driven by a high sedimentation rate and a constant reworking of the skeletal concentrations in the area. These conclusions agree with inferences from the models of fossil concentrations by Kidwell (1986) and Kidwell, Fürsich, and Aigner (1986).
In some cases, the bioerosion and encrustation contributed to the deterioration of the remains, producing the subsequent fragmentation of the shells. These effects would be expected to inhibit the conservation of the shells and could be the cause of their loss from the fossil record.
Several factors may be responsible for the low fouling density of mollusks. Among these are a relatively short exposure period before the burial (Brett et al., 2011); low nutrient levels (Lescinsky et al., 2002), constant movement of the rests or rapid wear of scales, and decreases in biological activity because of high turbidity (Parsons-Hubbard, 2005).
Formation of skeletal concentrations: In the fossil record, evidence of disturbance prior to burial can be used to infer peculiarities of the burial history or the life habits of the organisms (Brett & Baird, 1986). This association can be inferred in the present work thanks to the taphonomic analysis, in which cause-effect relationships of processes observed in Mandinga Lagoon could be established and contrasted with observations in the obtained sample.
Kidwell et al. (1986) recognized three types of processes responsible for skeletal concentration formations: sedimentological, biogenic and diagenetic. In Laguna de Mandinga, the taphonomic processes in mollusks are determined by a high rate of sedimentation and a constant reworking of skeletal concentrations. These conditions produce a high rate of fragmentation and a low degree of biogenic processes.
The formation of different biotopes (islets) in the lagoon traces to the constant dragging of sediments and dead organisms, which after a certain time are associated with a death assemblage (thanatocoenosis) with high sedimentation rates. Thus, as a result of constant reworking and high sedimentation rates, the encrusting organisms would rarely fossilize because of the long periods of time they require to become established on the remains (Asch & Collie, 2008).
Death assemblage represents the first step in the formation of a fossil assemblage, so our results represent a useful tool for biological and paleontological studies aimed at understanding the ecological, geological, and taphonomic processes that occur in marine-marginal environments with similar characteristics.
Ethical statement: authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.

