











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Los ecosistemas lóticos de montaña son sistemas complejos en su estructura y funcionamiento; en estos ocurren diferentes procesos que involucran variables físicas, químicas y biológicas, que interactúan de forma sinérgica y hacen que estos sistemas tengan alta heterogeneidad espacio-temporal (Allan & Castillo, 2007; Elosegi & Sabater, 2009). Así, esta variación espacio-temporal contribuye a la dinámica y distribución de las comunidades biológicas (Brooks, Haeusler, Reinfelds, & Williams, 2005; Vimos-Lojano, Martínez-Capel, Hampel, & Vázquez, 2019). Particularmente, los arroyos y ríos de los Andes albergan una alta diversidad biológica que se integra en los procesos ecosistémicos y determina el funcionamiento de éstos. Sin embargo, la más básica información sobre esta diversidad, historias de vida y roles ecológicos, es escasa para varios ecosistemas andinos de Colombia, Ecuador, Bolivia y Perú (Ramírez & Gutiérrez-Fonseca, 2014; Encalada, Flecker, LeRoy, Suárez, Herrera, Ríos-Touma, Jumani, Larson & Anderson, 2019).
El ensamblaje de macroinvertebrados acuáticos, es uno de los más representativos en los ríos de montaña tropical, se caracteriza por tener una fuerte relación con el ambiente circundante, por tanto, sus poblaciones son sensibles a las variaciones de las condiciones ambientales locales y regionales (Merritt, Cummins & Berg, 2008). Además, estos organismos contribuyen a la estructuración y funcionamiento del ecosistema, por ejemplo, mediante el procesamiento de materia orgánica y los flujos de nutrientes dentro de la red trófica (Bonada, Prat, Resh, & Statzner, 2006; Schmera, Heino, Podani, Erős, &Dolédec, 2017).
Son múltiples los factores que inciden sobre los macroinvertebrados en un río o arroyo de montaña. El ambiente físico es un factor fundamental, donde el sustrato y la dinámica hidráulica modifican la disponibilidad de recursos y la estructura de las poblaciones (Death & Winterbourn, 1995; Rempel, Richardson & Healey, 2000; Brooks et al., 2005; Mesa, 2010; Vimos-Lojano et al., 2019). La materia orgánica según la fuente, su forma y calidad son el otro factor de gran relevancia en la estructura de estos ensamblajes, especialmente en sistemas lóticos andinos (Lujan, Roach, Jacobsen, Winemiller, Vargas, Ching & Maestre, 2013).
Los macroinvertebrados poseen mecanismos de respuesta, conocidos como rasgos funcionales. Estos se definen como las características morfológicas, fisiológicas o ecológicas de los organismos que repercuten en su crecimiento, reproducción y supervivencia (Violle, Navas, Vile, Kazakou, Fortune, Hummel, & Garnier, 2007; Reynaga & Dos Santos, 2012). El estudio de los rasgos representa una gran aplicabilidad en biomonitoreo, conservación y restauración de ecosistemas lóticos a nivel global, siendo un enfoque complementario que permite aproximarse objetivamente a la comprensión de los roles ecológicos y por ende al funcionamiento y estado del ecosistema (Bonada et al., 2006; Schmera et al., 2017). El estudio de los rasgos biológicos y su relación con la expresión y variabilidad del entorno es la base de la diversidad funcional, entendida como el valor y el rango de aquellos componentes de la biodiversidad que determinan como opera o funciona un ecosistema (Tilman, 2001). Si bien su medición es compleja y se conocen múltiples formas de aproximarse a ella (Petchey, Hector & Gaston 2004; Ricotta, 2005), es importante considerar el efecto diferencial que puede tener cada población según su abundancia (Villéger, Mason & Mouillot, 2008).
Los estudios de macroinvertebrados en Colombia han adicionado en los últimos años un enfoque funcional, sin embargo, estos son escasos en comparación con lo reportado en Norteamérica y Europa (Richards, Haro, Johnson, & Host, 1997; Lamouroux, Dolédec, & Gayraud, 2004; Heino, 2005; Bonada, Doledec, & Statzner, 2007; Schmeraet al., 2017). La mayoría de estos estudios se enfocan en las estrategias alimentarias de la comunidad y se concentran en la región Pacífica, Caribe y en menor proporción en la región Andina (e.g., Chará-Serna, Chará, Zúñiga, & Pearson, 2012; Guzmán-Soto & Tamarís-Turizo, 2014; Longo & Blanco, 2014; Motta, Longo, & Aranguren-Riaño, 2017; Villada, Triana, & Dias, 2017). El objetivo de este estudio fue determinar la estructura, diversidad taxonómica y diversidad funcional del ensamblaje de macroinvertebrados bajo diferentes expresiones físicas en el río Hato Laguna, tributario del Lago de Tota. Se planteó como hipótesis de investigación: el ensamblaje presentará una mayor diversidad taxonómica en los sitios donde existan mayor heterogeneidad espacial; por otra parte, se espera que la diversidad funcional esté relacionada con la diversidad taxonómica, sin embargo, se harán evidentes los momentos y condiciones que promueven la diversificación de rasgos morfo-fisiológicos como respuesta a las diferentes expresiones del hábitat. Nuestra predicción es que, en el tramo donde la velocidad, profundidad y rugosidad sean altas, predominarían taxones de tamaño pequeño, formas de cuerpo flexible, adaptaciones de sujeción al sustrato, grupo funcional alimenticio filtrador. Por otro lado, el tamaño máximo, cuerpo cilíndrico, tolerancia moderada al déficit de oxígeno, endobentónicos y grupo funcional alimenticio colectores-recolectores se expresarían más en el tramo con bajos valores de velocidad y rugosidad.
Materiales y métodos
Área de Estudio: El estudio fue realizado en el río Hato Laguna (longitud 13 Km y área de drenaje de 34 Km2), tributario de la cuenca del Lago de Tota, en la Cordillera Oriental de los andes colombianos (CONPES 3801, 2014; Plazas, 2016). El patrón de lluvias es de tipo bimodal con una precipitación media anual entre 625 y 825 mm; con periodos máximos en los meses abril-mayo y octubre-noviembre, humedad relativa anual del 83% y temperatura media multianual de 11 °C (González & Cañón, 2016; Wanumen, 2019). La vegetación está constituida por relictos de bosque altoandino, y plantaciones de especies de aliso (Alnus sp.), pino (Pinus patula), eucalipto (Eucaliptus globulus) entre otras especies exóticas debido a los diferentes procesos de transformación antrópica (Moreno, 2015; Plazas, 2016).
Diseño de muestreo:Se realizaron cuatro muestreos en época de bajas precipitaciones entre noviembre del 2017 y febrero del 2018 (Fig. 1), en dos tramos: tramo alto (TA) ( 5° 37´2”N & 72° 52´35.2”) (Altitud: 3 332 msnm), con presencia de vegetación de ribera tipo arbórea con extensión de 30 m de ancho y tramo bajo (TB) ( 5° 35´ 18.6” N & 72° 53´ 34.5” W) (Altitud: 3 042 msnm) con presencia de vegetación de ribera tipo herbácea, arbustiva y cultivos aledaños de cebolla larga (Allium fistulosum), papa (Solanum tuberosum), arveja (Pisum sativum).
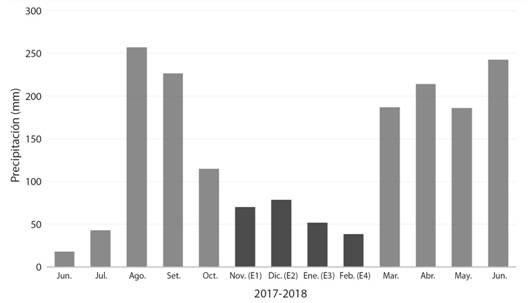
Fig. 1 Fig. 1 Valores totales de precipitación mensuales (mm). En oscuro se muestran los meses de muestreo: noviembre (E1)- diciembre (E2) 2017 y enero (E3) -febrero (E4) 2018. Tomado de: Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM). Total monthly precipitation (mm). In darker color, sampling months are shown: November (E1) -December (E2) 2017 and January (E3) -February (E4) 2018. Taken from: Institute of Hydrology, Meteorology and Environmental Studies (IDEAM).
En cada tramo (300 metros) se seleccionaron 15 puntos de muestreo de forma aleatoria estratificada: se estimó visualmente la proporción de cada tipo de mesohábitats presente para el cálculo del número de muestras biológicas a recolectarse de forma aleatoria en los mismos. Se realizó este tipo de muestreo ya que según Roleček, Chytrý, Hájek, Lvončík & Tichý, (2007), en medios ambientes heterogéneos se obtiene una alta representación de la variabilidad espacial.
Variables físicas: En cada tramo se determinó la proporción de los siguientes mesohábitats: rápido (alta-velocidad y flujo-turbulento), corriente (alta-velocidad y flujo laminar), poza (nula o baja velocidad de flujo) (Parasiewicz, 2007). Se midió el ancho húmedo del canal en tres puntos dentro del tramo, y por cada muestra biológica se estimaron: el tipo de sustrato como bloque (diámetro >1 m), canto (diámetro 1-0.5 m), guijarro (diámetro 0.5-0.2 m) y grava (diámetro 0.2-0.05 m) arenas (diámetro 0.005-0.002 m) y limo (diámetro < 0.002 m) (Apéndice 1) (Wadeson, 1994). La velocidad media de flujo (m/s) y la profundidad (m) se midieron con un Sensor Hidrometric OTT. La rugosidad se calculó mediante la proporción entre la longitud lineal/ longitud contorno del sustrato. A partir de estos valores se estableció el índice de rugosidad, en el que un mayor valor indica una menor rugosidad y un menor valor una mayor rugosidad (Motta, Obregón & Vimos, 2018).
Variables biológicas: Se recolectaron 15 muestras por tramo (n=2) y por muestreo (n=4) con un N total de 120, utilizando una red Surber (área de 0.09 m2, abertura de malla de 300 µm), por un tiempo de esfuerzo de 1 minuto cada muestra. El material recolectado fue preservado con etanol al 70 % y fue transportado al laboratorio donde los macroinvertebrados fueron separados del resto del material, realizando la determinación al mayor nivel de resolución taxonómica posible, mediante claves especializadas como las de: Pennak (1989), Merrit et al. (2008), Domínguez & Fernández (2009). Los especímenes fueron depositados en la colección limnológica de la Universidad Pedagógica y Tecnológica de Colombia (UPTC) con permiso de colecta dado en la resolución No. 01744 Autoridad Nacional De Licencias Ambientales (ANLA).
Rasgos funcionales: Los rasgos definidos abarcan seis categorías con sus respectivas modalidades; incluyen características morfológicas, fisiológicas y tróficas (Tabla 1). Se seleccionaron estas categorías de acuerdo con la posible relación con las variables físicas (Bonada, Rieradevall & Prat, 2007; Tomanova & Usseglio-Polatera, 2007). La asignación de los rasgos de: gremio trófico y largo del cuerpo se realizaron mediante mediciones directas de los taxones más abundantes (> 50 individuos en total), el resto de taxones, al igual que los rasgos forma del cuerpo, tipo de respiración, movilidad y grupos funcionales alimenticios se obtuvieron de información disponible de Tachet, Richoux, Bournaud, & Usseglio-Polatera, (2002); Tomanova & Usseglio-Polatera (2007); Chará-Serna, Chará & Zuñiga, (2012); Longo & Blanco, (2014).
Tabla 1 Table 1 Rasgos funcionales aplicados en el estudio con sus categorías y códigos Functional features applied in the study with its category and codes
| Rasgo | Categoría | Código |
| Largo del cuerpo(mm) | <2.5 | LA |
| 2.5-5 | LB | |
| 5-10 | LC | |
| 10-20 | LD | |
| 20-40 | LE | |
| 40-80 | LF | |
| >80 | LG | |
| Forma del cuerpo | Cilíndrico | Cil |
| Esférico | Esf | |
| Aplanado | Apla | |
| Optimizado | Opt | |
| Respiración | Branquias | Bran |
| Plastrón | Plas | |
| Tegumento | Teg | |
| Estigma | Est | |
| Movilidad | Volador | Vol |
| Nadador Superficial | N Sup | |
| Nadador Profundo | N Pro | |
| Rastrero | Rast | |
| Epibentónico | EpB | |
| Endobentónico | EnB | |
| Unido al Sustrato | U Sust | |
| Gremio Trófico | Material Inorgánico | M Ing |
| Materia orgánica particulada fina | MOPF | |
| Materia orgánica particulada gruesa | MOPG | |
| Micrófitas | Micro | |
| Macrófitas | Macro | |
| Animales | Ani | |
| Grupo Funcional Alimenticio | Colector | CL |
| Triturador | TR | |
| Raspador | RS | |
| Filtrador | FL | |
| Depredador | DP |
Basado en Tachet et al., 2002, Bonada et al., 2007 y Tomanova & Usseglio-Polatera, 2007.
Para las categorías de gremio trófico, se examinó el tracto digestivo de 1 038 organismos según lo propuesto por Muñoz, Romaní, Rodrigues, González, & García, (2009). Así, por cada taxón se realizó la disección de mínimo diez individuos y se procedió bajo un estereoscopio a extraer el tubo digestivo; el contenido se colocó en una placa de forma homogénea para su posterior observación en el microscopio. Por último, se seleccionaron 15 campos al azar, teniendo como área definida el cubreobjetos y se identificó de forma visual por medio de una reglilla ocular micrométrica las siguientes categorías: material inorgánico, materia orgánica particulada fina (MOPF) (tamaño < 50 μm), materia orgánica particulada gruesa (MOPG) (50 μm - 1 mm), micrófitas, macrófitas y restos de animales. Finalmente, se establecióla proporciónde cadacategoría en los campos observados (Apéndice 3). Dado que las asignaciones de estas categorías en su mayoría están adaptadas a los ecosistemas norteamericanos y europeos; se evaluó el contenido estomacal para complementar o confirmar la información de la literatura de referencia.
Análisis estadístico: Las variables físicas se analizaron realizando una ANOVA no paramétrica de Kruskal-Wallis (Guisande, Barreiro, Maneiro, Riveiro, & Vergara, 2006), para identificar posibles diferencias entre los tramos utilizando el programa PAST v.3.13; las variables fueron estandarizadas según lo propuesto por Guisande et al. (2006).
Se evaluó la variación en la composición y estructura de la comunidad de macroinvertebrados entre los tramos y muestreos mediante el análisis de similitud (ANOSIM) basado en Bray-Curtis, con 9 999 permutaciones en el programa PAST versión 3.13 (Clarke, 1993). Para aquellos casos en que se encontraron diferencias significativas mediante el análisis de similitud, se realizó un análisis de porcentaje de similitud (SIMPER) con el propósito de identificar las especies responsables de la diferenciación.
La representatividad de las muestras para hacer estimaciones de diversidad se evaluó mediante análisis de cobertura (Apéndice 4) (Chao & Jost, 2012):
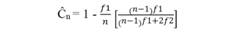
donde:
n= Abundancia Total
f1= Número de singletons (especies que tienen solo un individuo en la muestra de referencia)
Para estimar la diversidad taxonómica de los macroinvertebrados por tramos y muestreos, se calcularon los índices de diversidad de Simpson (1-D), índice de Shannon Wiener (H´) y el índice de equidad de Pielou (J´) (Moreno, 2001).
En cuanto a los rasgos funcionales, se construyó una matriz de los taxones con cada una de las categorías de los rasgos, estableciendo una codificación difusa: 0 (no tiene ningún vínculo), 1 (vínculo débil), 2 (vínculo moderado), 3 (vínculo fuerte); este enfoque integra la plasticidad de cada taxón teniendo en cuenta que este puede vincularse a más de una categoría por rasgo (Chevenet, Doléadec, & Chessel, 1994). Luego, esta matriz fue multiplicada por la matriz de las abundancias de los taxones con el fin de obtener una matriz final de los rasgos por tramo y muestreo. Con esta información se realizó un Análisis de Correspondencia Difusa (ACD) y un Análisis de Componentes Principales Difuso (ACPD) en el programa Rstudio versión 1.2.1335, con los cuales se determinó la relación de dependencia entre los rasgos funcionales y la variación temporal. Posteriormente, se identificaron los rasgos funcionales que presentaron diferencias significativas en el espacio y el tiempo, por medio de la prueba de Fisher.
La diversidad funcional se estimó como medidas del espacio funcional ocupado por las especies según la expresión de los rasgos, se consideró la riqueza funcional (RF), la divergencia funcional (DF) y la equidad funcional (EF) como descriptores (Villéger, et al.,2008). Los rasgos seleccionados según las categorías y codificación presentados en la tabla 1 y las abundancias relativas obtenidas a partir de los valores absolutos presentados en la tabla 2. Los descriptores de diversidad funcional fueron estimados mediante el paquete FD (Laliberté, Legendre, Shipley, Laliberté, 2014) en el software Rstudio versión 1.2.1335 (R Development Core Team, 2019).
TABLA 2 TABLE 2 Variables físicas registradas en el río Hato Laguna, tributario del Lago de Tota. Donde X, α y CV representan el promedio, desviación estándar y el coeficiente de variación, respectivamente. Las siglas en la columna izquierda indican la combinación del tramo (TA= tramo alto; TB = tramo bajo) y el momento de muestreo (E1= noviembre; E2= diciembre; E3= enero; E4= febrero) Physical variables registered in the Hato Laguna river, tributary of Lake Tota. Where X, α and CV represent the average, standard deviation and the coefficient of variation, respectively. The acronyms in the left column indicate the combination of the section (TA = high section; TB = low section) and the sampling events (E1 = November; E2 = December; E3 = January; E4 = February)
| Velocidad (m/s) | Profundidad (m) | Índice de rugosidad | Ancho (m) | |||||||||
| X | α | CV | X | α | CV | X | α | CV | X | α | CV | |
| TAE1 | 0.532 | 0.377 | 70.917 | 0.214 | 0.192 | 89.512 | 0.781 | 0.105 | 13.471 | 5.076 | 0.461 | 9.089 |
| TBE1 | 0.382 | 0.225 | 58.866 | 0.183 | 0.106 | 57.833 | 0.793 | 0.080 | 10.134 | 4.490 | 0.442 | 9.852 |
| TAE2 | 0.372 | 0.357 | 95.942 | 0.167 | 0.129 | 77.101 | 0.844 | 0.111 | 13.179 | 3.387 | 0.534 | 15.778 |
| TBE2 | 0.264 | 0.213 | 80.670 | 0.214 | 0.075 | 35.166 | 0.855 | 0.098 | 11.445 | 3.245 | 0.667 | 20.549 |
| TAE3 | 0.304 | 0.307 | 101.121 | 0.096 | 0.087 | 91.444 | 0.904 | 0.098 | 10.796 | 3.307 | 0.363 | 10.991 |
| TBE3 | 0.188 | 0.275 | 145.932 | 0.149 | 0.172 | 115.44 | 0.853 | 0.114 | 13.320 | 3.768 | 0.398 | 10.569 |
| TAE4 | 0.544 | 0.421 | 77.332 | 0.072 | 0.045 | 62.234 | 0.809 | 0.136 | 16.747 | 2.490 | 0.208 | 8.335 |
| TBE4 | 0.287 | 0.295 | 103.067 | 0.082 | 0.071 | 86.391 | 0.832 | 0.140 | 16.862 | 3.590 | 0.562 | 15.650 |
| Kruskal-Wallis | H | gl | p | H | gl | p | H | Gl | p | H | gl | p |
| Tramos | 0.162 | 7 | <0.05 | 0.232 | 7 | 0.450 | 0.137 | 7 | 0.210 | 0.001 | 7 | <0.05 |
| Muestreo | 3.560 | 3 | 0.312 | 24.530 | 3 | <0.001 | 1.520 | 3 | 0.676 | 17.700 | 3 | <0.001 |
Promedio (X), Desviación Estándar (α), Coeficiente de Variación (CV).
Para establecer las relaciones entre las variables físicas y los rasgos funcionales se realizó un Análisis de Correspondencia Canónica (ACC) empleando la matriz de rasgo por tramo y muestras y los datos físicos estandarizados. Previo al uso del análisis, se exploraron los datos mediante DCA (Análisis de correspondencias sin tendencias) para estimar la longitud de la gradiente, la cual fue de un gradiente 1.9 unidades de desviación estándar, correspondiente a un modelo unimodal seleccionado para el análisis (Lepŝ & Ŝmilauer, 2003). Por otro lado, las abundancias de individuos por cada taxón fueron transformados con log10(x + 1) y se eliminaron los taxones raros (abundancias <10 individuos del total) para el análisis. Para la validación del mismo se utilizó la prueba de permutación de Monte Carlo con 499 permutaciones. Además, se verificó que las variables físicas no estuvieran correlacionadas (Spearman: ρ> |0.8|). Por último, se aplicó la corrección de Bonferroni con el fin de evitar el error de tipo I en los datos. Este análisis se realizó con el programa CANOCO 5.
Resultados
Las variables físicas mostraron una variación entre tramos y muestreos. La velocidad del flujo fue mayor en el tramo alto (Kruskal Wallis test: H= 0.162, gl1= 7, p < 0.05), mientras que el ancho del canal fue mayor en el tramo bajo (Kruskal Wallis test: H= 0.001, gl1= 7, p < 0.05). Por otro lado, el ancho del canal (Kruskal Wallis test: H= 17.7, gl1= 3, p < 0.001) y la profundidad (Kruskal Wallis test: H= 24.530, gl1= 3, p < 0.001) tuvieron valores mayores en el muestro E1. Además, se observó que los valores de velocidad y la profundidad tuvieron un alto coeficiente de variación dentro de los tramos (>50%), respecto a los valores del índice de rugosidad y el ancho del canal (<20%) (Tabla 2).
Respecto de los macroinvertebrados, se recolectaron 4 531 individuos distribuidos en 38 taxones. El orden más diverso fue Diptera, distribuido en nueve familias, donde Chironomidae fue lamás diversa, con el 86 % de representación dentro del orden. Asimismo, la subfamilia de quironómidos Orthocladiinae presentóla mayor abundancia. Seguidamente, el orden Coleoptera estuvo representado por cuatro familias de lascuales la más diversa fue Elmidae con cuatro géneros. Por otro lado, el orden Trichoptera se registró en todos los tramos y muestreos con dos familias y dos géneros (Tabla 3). La composición y abundancia no mostraron diferencias significativas entre los tramos (ANOSIM, R= 0.037, p= 0.002) y muestreos (R= 0.167, p= 0.0001). Por otra parte, se resaltó la variación en la abundancia entre tramos de algunos taxones como Orthocladiinae sp7 (TA = 731, TB = 54 Individuos/m2), Nectopsyche sp.(TA = 48, TB = 232 Individuos/m2), Andesiops sp. (TA = 87, TB = 22 Individuos/m2) y Simulium sp. (TA = 167, TB = 76 Individuos/m2) entre muestreos los taxones Orthocladiinae sp2 (E1= 418, E2 = 95, E3= 70 y E4= 322 Individuos/m2), Orthocladiinae sp4 (E1= 301, E2 = 305, E3= 153 y E4= 419 Individuos/m2) (Tabla 3).
Tabla 3 Table 3 Lista de taxones y abundancia absoluta (Individuos/m2) de los macroinvertebrados en el río Hato Laguna en el Tramo Alto (TA) y Tramo Bajo (TB) y momentos de muestreo (E1, E2, E3 y E4) List of taxa and absolute abundance (Individuals / m2) of the macroinvertebrates in the Hato Laguna river in the High Section (TA) and Low Section (TB) and sampling events (E1, E2, E3 and E4)
| Orden | Familia | Taxón | TAE1 | TBE1 | TAE2 | TBE2 | TAE3 | TBE3 | TAE4 | TBE4 |
| Haplotaxida | Lumbriculidae | Lumbriculus sp. | 1 | 16 | 0 | 0 | 0 | 0 | 0 | 3 |
| Tubificidae | Tubificidae | 0 | 0 | 1 | 3 | 0 | 7 | 5 | 16 | |
| Naididae | Sthephensoniana sp. | 14 | 9 | 0 | 1 | 0 | 0 | 0 | 19 | |
| Ephemeroptera | Baetidae | Andesiops sp. | 75 | 12 | 10 | 8 | 2 | 1 | 0 | 1 |
| Baetodes sp. | 3 | 36 | 34 | 9 | 0 | 1 | 5 | 4 | ||
| Odonata | Aeshnidae | Aeshnidae | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trichoptera | Hydrobiosidae | Atopsyche sp. | 6 | 17 | 4 | 2 | 5 | 0 | 2 | 2 |
| Leptoceridae | Nectopsyche sp. | 8 | 39 | 9 | 87 | 27 | 12 | 4 | 94 | |
| Diptera | Blephariceridae | Blepharicera sp. | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| Ceratopogonidae | Ceratopogonidae | 0 | 0 | 1 | 1 | 0 | 17 | 11 | 91 | |
| Chironomidae | Chironominae | 0 | 3 | 2 | 5 | 0 | 15 | 0 | 21 | |
| Orthocladiinae sp1 | 19 | 77 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| Orthocladiinae sp2 | 120 | 298 | 27 | 68 | 27 | 43 | 202 | 120 | ||
| Orthocladiinae sp3 | 1 | 103 | 3 | 56 | 6 | 5 | 4 | 12 | ||
| Orthocladiinae sp4 | 87 | 214 | 206 | 99 | 54 | 57 | 149 | 270 | ||
| Orthocladiinae sp6 | 2 | 37 | 0 | 3 | 0 | 0 | 0 | 15 | ||
| Orthocladiinae sp7 | 95 | 3 | 336 | 7 | 13 | 0 | 287 | 44 | ||
| Tanypodinae | Tanypodinae | 2 | 7 | 7 | 1 | 9 | 3 | 30 | 3 | |
| Empididae | Empididae | 4 | 0 | 0 | 0 | 0 | 0 | 7 | 2 | |
| Muscidae | Muscidae | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | |
| Sciomyzidae | Sciomyzidae | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | |
| Simuliidae | Simulium sp. | 61 | 23 | 68 | 30 | 5 | 3 | 36 | 10 | |
| Tipulidae | Hexatoma sp. | 2 | 1 | 4 | 7 | 0 | 1 | 3 | 16 | |
| Limonia sp. | 0 | 1 | 3 | 0 | 0 | 2 | 0 | 1 | ||
| Tipula sp. | 3 | 7 | 1 | 0 | 2 | 0 | 3 | 0 | ||
| Coleoptera | Elmidae | Heterelmis sp. (Adt) | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Neoelmis sp.(Adt) | 9 | 4 | 6 | 4 | 26 | 0 | 4 | 3 | ||
| Neoelmis sp.(larv) | 10 | 1 | 1 | 1 | 3 | 1 | 2 | 20 | ||
| Hexacylloepus sp. (Adt) | 0 | 0 | 2 | 0 | 2 | 0 | 0 | 0 | ||
| Hexacylloepus sp.(Larv) | 8 | 0 | 4 | 14 | 7 | 4 | 5 | 8 | ||
| Ptilodactylidae | Ptilodactylidae | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Scirtidae | Scirtes sp. | 0 | 1 | 2 | 0 | 3 | 0 | 2 | 0 | |
| Staphylinidae | Staphylinidae | 0 | 0 | 0 | 8 | 1 | 0 | 5 | 0 | |
| Neoophora | Dugesiidae | Dugesia sp. | 1 | 2 | 0 | 0 | 0 | 0 | 4 | 5 |
| Amphipoda | Hyalellidae | Hyallela sp. | 0 | 12 | 0 | 1 | 1 | 2 | 0 | 2 |
| Isopoda | Sphaeromatidae | Sphaeroma sp. | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Abundancia Total | 534 | 924 | 733 | 418 | 193 | 175 | 770 | 784 | ||
| Riqueza Total | 24 | 24 | 23 | 24 | 17 | 17 | 20 | 26 | ||
Respecto a la diversidad, la cobertura de la muestra estimada fue superior al 97% en todos los casos, lo que permite mayor confiabilidad en los estimadores de diversidad (Apéndice 4). Así, los resultados de los índices de Simpson, Shannon y Pielou, muestran variación en la estructura taxonómica de los macroinvertebrados entre tramos y entre muestreos, sin que se logre identificar un patrón o tendencia en su expresión (Tabla 4).
TABLA 4 TABLE 4 Índices de diversidad taxonómica aplicados al ensamblaje de macroinvertebrados en el río Hato Laguna. Índice de Simpson (1-D), Índice de Diversidad de Shannon-Wiener (H´) eÍndice deEquidad de Pielou (J´). Las siglas en la columna izquierda indican la combinación del tramo (TA= tramo alto; TB = tramo bajo) y el momento de muestreo (E1= noviembre; E2= diciembre; E3= enero; E4= febrero) Taxonomic diversity indices applied to the macroinvertebrate assemblages in the Hato Laguna river. Simpson Index (1-D), Shannon-Wiener Diversity Index (H´) and Pielou Equity Index (J´). The acronyms in the left column indicate the combination of the section (TA = high section; TB = low section) and the sampling events (E1 = November; E2 = December; E3 = January; E4 = February)
| Tramo Alto | Simpson (1-D) | Shannon (H´) | Pielou (J´) | Tramo Bajo | Simpson (1-D) | Shannon (H´) | Pielou (J´) |
| TAE1 | 0.856 | 2.231 | 0.702 | TBE1 | 0.816 | 2.118 | 0.667 |
| TAE2 | 0.696 | 1.615 | 0.515 | TBE2 | 0.847 | 2.206 | 0.694 |
| TAE3 | 0.853 | 2.243 | 0.792 | TBE3 | 0.808 | 2.04 | 0.72 |
| TAE4 | 0.752 | 1.748 | 0.584 | TBE4 | 0.823 | 2.203 | 0.676 |
En cuanto a los rasgos funcionales, en general se resalta la dominancia de ciertas categorías tales como: longitud del cuerpo entre 20-40 mm, forma del cuerpo cilíndrico, respiración por branquias, la categoría de movilidad rastrero y epibentónico, tipo de alimentación de MOPF y MOPG, y el grupo funcional alimenticio colector y filtrador.
Según la prueba de Fisher las categorías que presentaron diferencias significativas (p< 0.05) entre tramos y muestreos fueron; largo del cuerpo de 20 a 40 mm (tramos), forma del cuerpo cilíndrico (muestreos), respiración por branquias (tramos), movilidad de tipo rastrero, gremio trófico MOPF y grupo funcional alimenticio colector
En el ACPD el porcentaje de inercia proyectado en el eje uno fue de 60.64% y en el eje dos de 18.24 %. Según este análisis, el tramo alto en los muestreos (E2 y E4) se asoció con los rasgos; largo del cuerpo 5 a 10 mm, respiración por tegumento, gremio trófico MOPF y grupo funcional alimenticio colector (Fig. 2, Lado derecho). El tramo bajo en el muestreo (E2) y tramo alto (E3) se asociaron con tamaño del cuerpo de 10-20 mm, gremio trófico MOPG, grupo funcional alimenticio triturador (Fig. 2, Lado izquierdo). Por otra parte, los tramos bajos en los muestreos (E3 y E4) presentaron mayor proporción de los rasgos: forma del cuerpo aplanado, respiración por branquias, gremio trófico micrófitas y grupo funcional alimenticio raspador (Fig. 2, Parte inferior central).
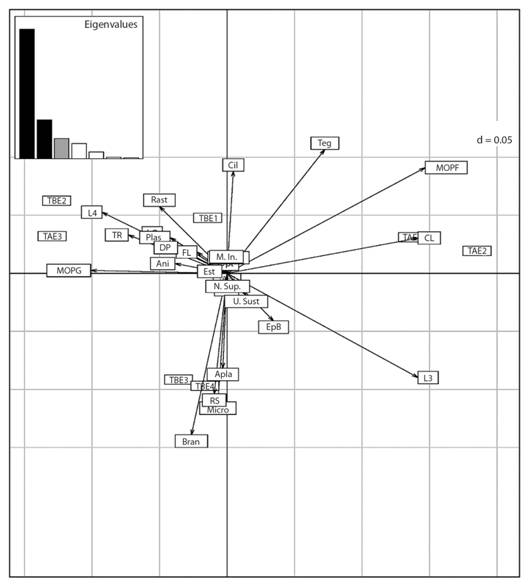
Fig. 2 Fig. 2 Análisis de componentes principales difuso (ACPD) de las categorías de los rasgos en relación con la variación temporal. E1 (noviembre), E2 (diciembre), E3 (enero), E4 (febrero). Tramo Alto (TA), Tramo Bajo (TB). Ver códigos de los rasgos funcionales en la Tabla 1. Fuzzy Principal Component Analysis (ACPD) of the trait categories in relation to temporal variation. E1 (November), E2 (December), E3 (January), E4 (February). High Section (TA), Low Section (TB). See codes of functional trait in Table 1.
Los descriptores de diversidad funcional muestran diferencias entre tramos y muestreos (Fig. 3). La riqueza funcional es mayor en el tramo alto, aunque en E4, se expresan magnitudes similares entre los tramos, se destaca una reducción de la RF en el E3 para ambos tramos. La equidad funcional muestra una tendencia similar entre tramos, sin embargo, en el E4 este descriptor se incrementa en el tramo alto. Respecto a la divergencia funcional, el tramo alto presenta mayor expresión de DF consistente durante los cuatro muestreos.
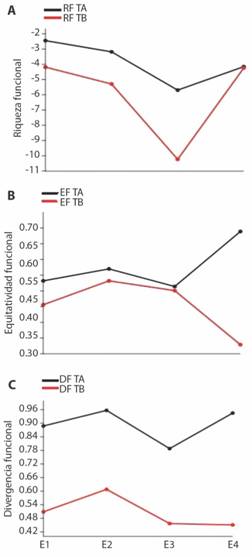
Fig. 3 Fig. 3 Índices de diversidad funcional aplicados al ensamblaje de macroinvertebrados por tramos y muestreos. (A). Riqueza Funcional (RF), (B) Equidad Funcional (EF), (C) Divergencia Funcional (DF). Functional diversity indices applied to the macroinvertebrate assemblages by sections and samples. (A). Functional Richness (FR), (B) Functional Evenness (FE), (C) Functional Divergence (FD).
Respecto al ACC de los rasgos funcionales y las variables físicas, se presentó una inercia total de 81%, con un valor de significancia de (p <0.05). El primer eje explicó el 46% de la varianza y en el segundo el 35% (Fig. 3). El primer eje se relacionó con valores altos de ancho del canal (p< 0.01, 52.4% contribución) y velocidad (p< 0.01, 33.4% contribución), que a su vez se relacionaron con las categorías unido al sustrato, endobentónico y volador. Por otro lado, el segundo eje se relacionó con valores altos velocidad (p< 0.01, 47.2% contribución), valores bajos de ancho del canal (p< 0.01, 30.1% contribución), rugosidad (p< 0.05, 22.54% contribución) y con las categorías forma del cuerpo aplanado, tipo de alimentación micrófitas y tamaño del cuerpo mayor a 80 mm (Fig. 4).
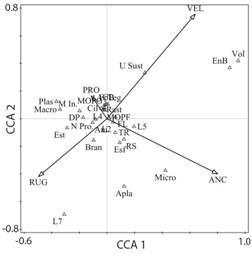
Fig. 4 Fig. 4 Análisis de Correspondencia Canónica (ACC) entre los rasgos funcionales y las variables físicas. Velocidad (VEL), Rugosidad (RUG), Profundidad (PRO), Ancho (ANC). Varianza explicada, Componente 1 (horizontal): 46%, Componente 2 (vertical): 35%. Test de permutación Monte Carlo (p= 0.034). Ver códigos de los rasgos funcionales en la Tabla 1. Canonical Correspondence Analysis (CCA) between functional traits and physical variables. Speed (VEL), Roughness (RUG), Depth (PRO), Width (ANC). Variance explained, Component 1: 46%, Component 2: 35%. Monte Carlo permutation test (p = 0.034). See codes of functional trait in Table 1.
Discusión
Este estudio demostró que las variables físicas mostraron fluctuaciones entre tramos y muestreos. De esta forma, la variación en la velocidad y ancho del canal entre los tramos se puede explicar por los cambios a lo largo del río en la pendiente, geomorfología del canal y cantidad de flujo transportado (Vannote, Minshall, Cummins, Sedell, & Cushing, 1980; Montgomery & Buffington, 1998). Estas variaciones longitudinales generan, en los tramos altos de los ríos, condiciones hidráulicas heterogéneas en la profundidad y altas velocidades del flujo, mientras que en los tramos bajos disminuye la velocidad debido a una menor pendiente y aumento del ancho del canal por el incremento en la cantidad de flujo transportado (Allan & Castillo, 2007; Elosegi & Sabater, 2009). Por otro lado, se observó que a pesar de que todos los muestreos corresponden a épocas de bajas precipitaciones, algunas variables como ancho y profundidad cambiaron entre muestreos. Esto se puede explicar por el tamaño pequeño de la cuenca, que genera una respuesta rápida de aumento de flujo incluso con precipitaciones esporádicas, que se ve reflejado en el aumento del ancho y la profundidad en el muestreo E1 (noviembre) (IDEAM, 2018; Lewis, 2008).
En general, los órdenes Diptera, Coleoptera y Trichoptera mostraron la mayor riqueza taxonómica. Estos grupos se consideran habituales en los sistemas de montaña andinos (Domínguez & Fernández, 2009; Longo, Zamora, Guisande, & Ramírez, 2010; Vásquez-Ramos & Reinoso, 2012; Roldán, Zúñiga, Zamora, Reinoso & Longo, 2014). En cuanto a la estructura del ensamblaje, los taxones dominantes encontrados en el estudio pertenecen a la subfamilia Orthocladiinae (Chironomidae), reportados para ríos de alta montaña de Colombia, Ecuador y Perú (Prat, Ribera, Rieradevall, Villamarin, & Acosta, 2013). Estos taxones son considerados abundantes en estos sistemas acuáticos y pioneros en la recolonización (Acuña, Muñoz, Giorgi, Omella, Sabater, & Sabater, 2005), debido a su capacidad de dispersión, a sus hábitos tróficos generalistas y a sus adaptaciones para evitar las condiciones hidráulicas extremas relacionadas con su ciclo de vida o el tamaño del cuerpo pequeño que los favorece en la búsqueda de refugios para evitar la fuerza de arrastre de la corriente en velocidades altas (Matthaei, Arbuckle, & Townsend, 2000; Henriques-Oliveira, Nessimian, & Dorvillé, 2003; Mesa, 2010; Vimos-Lojano, 2017). Lo anterior pudo explicar su dominancia en los tramos altos donde la velocidad fue mayor pero también en menor proporción en los tramos bajos donde la velocidad disminuyó y aumentó el ancho de canal y la profundidad.
Por otra parte, la composición y la abundancia de los organismos no presentaron diferencias contrastantes entre tramos y muestreos según los análisis realizados. Sin embargo, se resalta la variación en la abundancia de taxones como los dípteros Orthocladiinae y Simulium sp., el efemeróptero Andesiops sp. y el tricóptero Nectopsyche sp. Este último, tuvo mayor abundancia en los tramos bajos en donde las velocidades de flujo fueron menores. Este hallazgo concuerda con lo reportado por Ríos-Touma, Encalada & Prat. (2011) y Vimos-Lojano, Martínez-Capel & Hampel (2017) para sistemas de montaña andinos de Ecuador, en donde se demuestra que Nectopsyche sp. tiene una preferencia marcada por hábitats con bajas velocidades donde se acumula material de piedras y hojas en el lecho del río que es utilizado para la construcción de estuches característicos de este género y esta materia orgánica gruesa también puede ser aprovechada como recurso alimenticio por estos organismos trituradores (Holzenthal & Ríos-Touma, 2018).
Por otro lado, Orthocladiinae, Simulium sp. y Andesiops sp. fueron más abundantes en los tramos altos. Este último, presenta un tamaño pequeño y uñas tarsales con dentículos que le confieren adherencia al sustrato y le permiten contrarrestar la fuerza de arrastre en condiciones de altas velocidades. (Domínguez & Fernández, 2009) Asimismo, Simulium sp. se caracteriza por ser una especie reofílica, destacándose por su capacidad de adherencia al sustrato por medio de un anillo de ganchos anales y la utilización de un hilo de seda, siendo considerada una especie con movilidad epibentónica. (Pavitra, Low, Tan, Lim & Ya´cob, 2019)
La diversidad taxonómica no expresó tendencias marcadas por tramos o muestreos, según Poff (1997) y Jacobsen (2004) esto puede explicarse porque estos estimadores están relacionados con patrones que se presentan a escalas espaciales mayores (segmento, río) como variaciones geomorfológicas, cambios en el tipo de vegetación y usos del suelo; así mismo, el estudio abarco cuatro meses, lo que limitaría ver cambios estacionales. (Villamarín, 2008). Sin embargo, Carrera & Gunkel (2003); Rodríguez-Barrios, Ospina-Torres & Turizo-Correa (2011), han encontrado que variaciones ambientales a escalas pequeñas en las condiciones físicas de velocidad, profundidad y ancho de canal como las halladas en el sistema de estudio, pueden incidir en la diversidad taxonómica. Por ejemplo, se ha reportado que un aumento en la velocidad y profundidad del flujo están relacionados con una menor diversidad de macroinvertebrados acuáticos en ríos de Chile y Argentina (Oyanedel, Valdovinos, Azócar, Moya, Mancilla, Pedreros & Figueroa, 2008; Moya, Valdovinos, Moraga, Romero, Debels & Oyanedel, 2009). Para el caso de estudio, en el tramo bajo con menor velocidad y mayor ancho del cauce, la tendencia a una mayor diversidad en los estimadores H´ y 1-D, está relacionada con mayor grado de equidad.
En cuanto a los rasgos funcionales, se destaca la dominancia generalizada de organismos colectores que se alimentan de MOPF. Estos hallazgos coinciden con lo reportado por Moretti, (2005), Tomanova, Goitia, & Helešic, (2006) y Rivera, Zapata, Pérez, Morales, Ovalle, & Alvarez (2010). Estos autores sugieren que la dominancia de este grupo en los ríos Neotropicales, se debe al aporte de MOPF derivada de la cuenca y/o de la descomposición por microorganismos y/o la abrasión física de la MOPG que posteriormente es acumulada por procesos de la hidráulica del río (Príncipe, Gualdoni, Oberto, Raffaini, & Corigliano, 2010; Meza-S, Rubio-M, Dias, & Walteros, 2012).
Las evidencias de variación en algunos rasgos funcionales, se observaron en el tramo alto en los muestreos de diciembre (E2) y febrero (E4) con aumento de la dominancia de los organismos que se alimentan de MOPF (colectores) como los Orthocladiinae, lo que se puede explicar por la alta velocidad del flujo en este tramo, dado por la pendiente que genera mayor transporte y posterior acumulación de sedimentos y materia orgánica particulada fina, fuente de alimento para los organismos colectores (Montgomery & Buffington, 1998; Chará-Serna, Chará, Zuñiga, Pedraza & Giraldo, 2010; Rolls, Leigh, & Sheldon, 2012). Estos resultados sugieren que procesos descritos a gran escala como los propuesto para regiones templadas por Vannote et al. (1980), en los que los colectores dominan en la parte baja de la cuenca debido a la relevancia de procesos de depósitos de sedimentos, podría expresarse en pequeña escala bajo condiciones locales que favorecen la acumulación de MOPF como la presencia de condiciones de velocidad heterogénea como las reportadas en este estudio para el tramo alto.
Por otro lado, los organismos que respiran por tegumento como Tanypodinae y Chironominae también se relacionaron con el tramo alto. El incremento de este rasgo en el tramo alto coincide con lo reportado por Statzner, Dolédec & Hugueny, (2004) y Reynaga & Dos Santos, (2012) acerca de la relación entre este tipo de respiración y la mayor captación de oxígeno en las partes altas de las quebradas por la reareación producida por la acción de la corriente. Por otro lado, Motta et al. (2017), reportan mayor abundancia de este rasgo en sitios con concentraciones bajas de oxígeno en un río tropical de la Isla de Providencia.
Así mismo, en los tramos altos en los muestreos de diciembre (E2) y febrero (E4), hubo mayor cantidad de organismos con tamaño pequeño (5-10 mm); se sugiere que puede estar relacionado con la alta velocidad del flujo, que se ve reflejada en una menor inversión en el crecimiento por parte de los organismos como estrategia para buscar refugios y evitar ser arrastrados por la corriente (Townsend & Hildrew, 1994; Brooks et al., 2005; Beche, Mcelravy, & Resh, 2006; Reynaga & Dos Santos, 2012; Dolédec, Tilbian, & Bonada, 2017).
Los tramos bajos en los muestreos de enero y febrero (E3 y E4) se asociaron con la presencia de organismos raspadores como Neoelmis sp. (larva) que se alimentan de micrófitas, respiración por branquias como Andesiops sp., y forma del cuerpo aplanada. La predominancia de organismos con estructuras especializadas como las branquias a menores altitudes en ríos de montaña coincide con lo reportado por Tomanova & Usseglio-Polatera, (2007); Statzner & Beche, (2010); Reynaga & Dos Santos, (2012), acerca de una mayor tolerancia a menores valores de oxígeno con este tipo de respiración.
La expresión de los rasgos descrita, se refleja en los estimadores de diversidad funcional, especialmente en la riqueza (RF) y la divergencia (DF). Respecto a la mayor RF en el tramo alto, se interpreta como un espectro dimensional más amplio en la expresión de rasgos con menor grado de redundancia entre los grupos de macroinvertebrados (Villéger, et al., 2008; Stubbington, Bogan, Bonada, Boulton, Datry, Leigh & Vander Vorste, 2017). Se destaca en el muestreo E3, una reducción en la RF en ambos tramos, lo que coincide con un periodo de menor velocidad de flujo, esto se explica en un mayor grado de redundancia en la expresión de los rasgos de los macroinvertebrados con la consecuente reducción en el espectro multidimensional funcional. La DF mayor en el tramo alto corresponde con la tendencia a una mayor dominancia de grupos como Orthocladiinae, Chironominae, Simulium sp. y Andesiops sp., que a su vez expresan rasgos relevantes y característicos en la comunidad (Villéger, et al., 2008; Menezes, Baird & Soares, 2010).
En cuanto a las asociaciones entre las variables físicas y los rasgos funcionales, se encontró que valores altos de velocidad se relacionan con estrategia de movilidad de adherencia al sustrato, endobentónico y volador. Esto se explica, teniendo en cuenta que los organismos encontrados en las rocas en ríos de montaña tienen estructuras especializadas para adherirse al sustrato (uñas anales o tarsales y ventosas) que les confieren cierta resistencia contra la fuerza de arrastre (Townsend & Hildrew, 1994; Lamouroux et al., 2004; Tomanova & Usseglio-Polatera, 2007; Reynaga & Dos Santos, 2012). Asimismo, los organismos endobentónicos pueden escapar de las altas velocidades, enterrándose en los sedimentos del lecho. Lo anterior corrobora la importancia de la velocidad en la expresión de estos rasgos de la comunidad de macroinvertebrados (Lamouroux et al., 2004; Tomanova et al., 2006; Tomanova & Usseglio-Polatera, 2007; Príncipe et al., 2010; Dolédec et al., 2017).
Se concluye que se presentaron cambios entre tramos y muestreos en las condiciones hidráulicas y grupos funcionales de los macroinvertebrados. Los tramos altos se caracterizaron por: alta velocidad de flujo, mayor abundancia de taxones como Orthocladiinae, Andesiops sp., Simulium sp. y aumento en la ocurrencia de categorías de rasgos funcionales como gremio trófico MOPF (colectores), respiración por tegumento y tamaño del cuerpo pequeño. En contraste, los tramos bajos se asociaron con la presencia de organismos como Nectopsyche sp. y categorías de rasgos como raspadores que se alimentan de micrófitas, respiración por branquias y forma del cuerpo aplanada. Se resalta el papel de la velocidad del flujo como una variable determinante en la ocurrencia del rasgo funcional de tipo de movilidad específicamente de las categorías adherido al sustrato, endobentónico y volador. Asimismo, se resalta la diversidad funcional como una expresión de los rasgos descritos, por lo cual se relaciona en los tramos altos con la dominancia de taxones como Orthocladiinae, Chironominae, Simulium sp. y Andesiops sp.
Se recomienda para futuros trabajos, determinar la diversidad taxonómica y diversidad funcional en diferentes escalas espaciales evaluando diferentes variables físicas y químicas con el fin de aproximarse a una comprensión menos sesgada del funcionamiento de los ecosistemas; de igual manera evaluar los rasgos funcionales con un mayor número de muestreos en un período interanual al igual que incluir un mayor número de rasgos morfológicos, fisiológicos, ecológicos.
Declaración de ética: los autores declaran que están de acuerdo con esta publicación; que no existe conflicto de interés de ningún tipo; y que ha cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

