











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
Remote locations are colonized by long-distance dispersal (Jordano, 2017), the actual mechanism of colonization is organism-dependent (Gillespie et al., 2012). Large terrestrial animals use rafts of vegetation and their patterns of dispersal depend on surface marine current, as it has been the case for Anolis lizards in the Bahamas (Calsbeek & Smith, 2003). A similar situation happens with plants with large seeds (Nathan et al., 2008). Some terrestrial organisms disperse attached to the body or inside the guts of migratory birds (Gillespie et al., 2012). For marine organisms, sea currents play an important role in defining the direction of the dispersal, while the larval duration will determine how far the organisms can be transported (Shanks, 2009).
Small terrestrial invertebrates disperse in a passive manner through wind currents, forming what is known as aerial plankton. Winged animals get caught on air currents while flying. In the case of spiders, they actively start dispersal with ballooning (Larrivée & Buddle, 2011). The take-off required for ballooning could be initiated by different strategies as the tip-toe launch or from a suspended position at a high structure (Bell, Bohan, Shaw, & Weyman, 2005). Given some environmental cues spiders start to spin silk to the air until it gets caught by wind currents. Simulations suggest that this behavior appears to be optimal in non-ideal convection conditions, which means warm temperatures with a light breeze (Reynolds, Bohan, & Bell, 2007). Recent studies have shown that the time to perform this behavior is also favored by the detection of electric fields (Morley & Robert, 2018). Ballooning has been showed to occur in open fields (Okuma & Kisimoto, 1981) and closed forest (Larrivée & Buddle, 2011). It is not exclusive for spiders, it also has also been reported on spider mites (Acari) and larvae of moths (Lepidoptera) (Bell et al., 2005).
Indirect evidence about the groups of organisms that disperse by wind currents was provided on the seminal experimental studies to test the hypothesis of island biogeography. Indeed, transport through rafts was described not as important as air-based colonization on the early stages of colonization after the experimental defaunation (Simberloff & Wilson, 1969). Similarly, spiders were also among the early colonizers on land cleared by the volcanic explosion on the island of Krakatau (Bristowe, 1931).
Direct collections with entomological nets placed in airplanes and ships also demonstrate that there is an important amount of terrestrial invertebrates floating on the air column (Gressitt, Leech, & O’Brien, 1960; Hardy & Cheng, 1986). Surveys from ships going around Antarctica have demonstrated the presence of Araneida, Collembola, Psocoptera, Thysanoptera, Heteroptera, Coleoptera, Diptera, and Hymenoptera (Gressitt et al., 1960). Sub-Antarctic surveys combining land trapping and nets on ships have resulted in the collection of Acari, Collembola, Homoptera, Coleoptera, Diptera, and Hymenoptera (Gressitt, Leech, Leech, Sedlacek, & Wise, 1961). While, collections across the north tropical Pacific have reported Hymenoptera, Diptera, Coleoptera, Hemiptera, Thysanoptera, Arachnids (Yoshimoto & Gressitt, 1960; Holzapfel & Perkins, 1969) and even a marine bivalve (Yoshimoto & Gressitt, 1961). Surveys on the East China Sea focused on arachnids, reported that the dominant (101 out of 105 specimens) genus was Tetragnatha (Tetragnathidae). The other families found corresponded to Theridiidae, Erigonidae, and Lyniphiidae (Okuma & Kisimoto, 1981).
In the case of the Eastern Tropical Pacific, there are no previous reports of aerial plankton. Within the region, Isla del Coco is an oceanic island located 550 km from Costa Rica and 680 km from the Galápagos. Among the climatic features that characterize the island is the heavy rainfall (7 m), as a result of being in the path of the Intertropical Convergence Zone (ITCZ), and the presence of south-westerlies trade winds (Amador et al., 2016). The month of September is historically the one with the highest frequency of named tropical storms, hurricanes and major hurricanes (Amador et al., 2016). The island was originated on the Galápagos hotspot. The current emerged land has between 1.9 and 2.4 million years (Bellon, Saenz, & Tournon, 1983; Castillo et al., 1988), which suggests it corresponds to later volcanic activity. Therefore, all the terrestrial biodiversity colonized during the last couple of millions of years. As is expected for a remote island, it lacks many groups of organisms of large size present in the continent.
The direct evidence from aerial trapping and indirect evidence from the species present on remote islands provide a clear idea of which groups are the most abundant and effective components of the aerial plankton. However, the amount of available data is geographically sparse and reduced. Increasing the number of locations will give a better picture of which are the most common groups at a global level or how do they compare between geographic regions. Also, better sampling in places like Isla del Coco will provide data to regionally understand the process of long-distance colonization. Here, we characterize the aerial plankton present on the transect Puntarenas - Isla del Coco on 2 roundtrip voyages during July 2017 and August 2018. This study corresponds to the first report for this part of the tropical Pacific.
Materials and methods
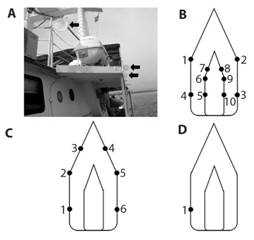
Collections were performed during 4 travels on the transect Puntarenas - Isla del Coco: (1) 08th -10th July 2017, (2) 23rd-25th July 2017, (3) 12th-14th August 2018 and 22nd-24th August 2018. On trip number 1, an array with 10 entomological nets (approx. 50 cm diameter each) was placed around the boat (Fig. 1A, Fig. 1B). A similar arrangement was used on trip number 2, but with only 6 nets (Fig 1C). Nets were checked every 6 hours starting from the departure from Puntarenas or the island. On trips number 3 and 4, only one net was placed on towards the bow on the stern (Fig. 1D). The net was checked every 8 hours. The nets were cleaned and checked before being placed in the boat in order to ensure that all insects collected corresponded to aerial plankton. Whenever nets were checked the location was recorded with a GPS. Specimens collected were preserved in ethanol 100%. Opportunistic collections on the deck were also done and GPS point recorded.
Results
On the first three trips, a total of 18 individuals were collected corresponding to 10 morpho-species from the orders: Diptera (Ceratopogonidae, Milichiidae, and a non-unidentified family of Nematocera), Hymenoptera (Chalcidoidea: Agaonidae), Hemiptera (Scutelleridae and Coreidae) and Lepidoptera (Sphingidae). The first two orders (Diptera and Hymenoptera) were collected on the nets, while the other two (Hemiptera and Lepidoptera) were found on the deck of the boat (Table 1, Table 2, Table 3). Each collection event corresponds to a single individual, except for the Sphingidae months, which numbers are specified on the tables. On the trip from Isla del Coco to Puntarenas departing on August 22nd, 2018 nothing was collected.
TABLE 1 Collections of aerial plankton on transect Puntarenas - Isla del Coco, during July 2017 onboard Okeanos I
| Point | Date | Time (hrs.) | Coordinates | Distance to nearest continental land (km) | Collections |
| PUNTARENAS | July 8th, 2017 | 14:25 | -84.835352°, 9.980183° | 0 | - |
| AP1 | 15:25 | -84.784242°, 9.869021° | 6 | Net #5: Diptera: Nematocera, probably Scatopsidae Net #6: Diptera: Nematocera: Ceratopogonidae | |
| AP5 | 20:25 | -85.104346°, 9.221228° | 35 | Net #4: Diptera: Nematocera: Ceratopogonidae | |
| AP8 | July 9th, 2017 | 2:25 | -85.513157°, 8.454209° | 130 | - |
| AP9 | 8:25 | -85.921614°, 7.682497° | 226 | - | |
| AP10 | 14:25 | -86.341958°, 6.886541° | 325 | Net #9: Hymenoptera: Chalcidoidea: Agaonidae | |
| AP11 | 21:00 | -86.799227°, 6.030168° | 432 | - | |
| ISLA DEL COCO | July 10th, 2017 | Approx. 8:00 | -87.055579°, 5.544034° | 492 | - |
TABLE 2 Collections of aerial plankton on transect Isla del Coco - Puntarenas, during July 2017 onboard Okeanos II.
| Point | Date | Time (hrs.) | Coordinate | Distance to nearest continental land (km) | Events |
| APR1 | July 23rd, 2017 | 14:45 | -86.977001°, 5.641371° | 479 | - |
| APR2 | 20:45 | -86.587383°, 6.395585° | 385 | - | |
| APR3 | July 24th, 2017 | 3:54 | -86.103414°, 7.323246° | 270 | - |
| APR4 | 8:45 | -85.744103°, 7.989793° | 187 | Deck: Diptera: Nematocera: Ceratopogonidae | |
| (AP4.1R) | 14:25 | -85.334508°, 8.761253° | 92 | Deck: Diptera: Acalyptratae: Milichiidae | |
| AP5R | 14:52 | -85.318124°, 8.794723° | 87 | Deck: Diptera: Nematocera: Chironomidae or Ceratopogonidae | |
| AP6R | 20:45 | -84.938296°, 9.518006° | 18 | Deck: Hemiptera: Scutelleridae “shield-backed bug“ | |
| (AP6.1R AP6.2R) | 22:12 | -84.861155°, 9.660273° -84.844015°, 9.692802° | 12 | Near lights on deck: Lepidoptera: Sphingidae, Nyceryx ericea(Druce, 1888) (5 individuals) | |
| PUNTARENAS | July 25th, 2017 | 5:55 | -84.835352°, 9.980183° | 0 | Net #4: Diptera Net #5: Diptera |
TABLE 3 Collections of aerial plankton on transect Puntarenas - Isla del Coco, during August 2018 onboard Okeanos II
| Point | Date | Time | Coordinate | Distance to nearest continental land (km) | Events |
| PUNTARENAS | August 12th, 2018 | 14:50 | -84.835352°,9.980183° | 0 | - |
| AP1-18 | 19:56 | -84.993768°,9.382055° | 23 | Deck: Hemiptera: Coreidae “Leaf legged bug” | |
| AP2-18 | 20:45 | -85.0414°, 9.29134° | 30 | Near lights on deck: Lepidoptera: Sphingidae, Aellopos (2 individuals) | |
| AP3-18 | August 13th, 2018 | 8:12 | -85.74355°, 7.988447° | 187 | - |
| AP4-18 | 17:00 | -86.265954°,7.011534° | 309 | - | |
| AP5-18 | 21:47 | -86.561589°, 6.456803° | 378 | - | |
| AP6-18 | August 14th, 2018 | 4:37 | -87.012202°, 5.610432° | 484 | - |
Discussion
Cross oceanic dispersal of small arthropods occurs through wind currents. The composition and geographic distribution of the aerial plankton are highly understudied, yet essential to understand island colonization processes. Here, we characterize the aerial plankton on the transect Puntarenas - Isla del Coco. This portion of the Eastern Tropical Pacific has not been previously surveyed and represents a high interest for the historical biogeography of the oceanic islands present on the region (Revillagigedo archipelago, Clipperton, Isla del Coco, Malpelo, and Galápagos).
Regardless a small sampling size of 4 trips (36 hrs. each) on the transect, with half of them using only one sampling net, it was possible to collect representatives of 3 different families: Ceratopogonidae, the unidentified Nematocera (Diptera) and Agaonidae (Hymenoptera). In terms of the total amount of specimens, this result is similar to previous studies across the Tropical Pacific where they collected around 1 or 2 individuals per day (Yoshimoto &Gressitt, 1960). Moreover, other 5 families (Diptera: Milichiidae; Hemiptera: Scutelleridae and Coreidae; and Lepidoptera: Sphingidae) were collected on the deck of the vessel.
The predaceous midges Ceratopogonidae (Diptera) were the most commonly collected in the aerial nets. They have a worldwide distribution and are associated with aquatic environments. This family has been recorded on Isla del Coco (Hogue & Miller, 1981; Gutiérrez-Fonseca, Ramírez, Umaña, & Springer, 2013). The one collected on this study was found 35 km from the continent, most likely it was carried from the mainland by the wind. Ceratopogonidae have been also found on collection trips on the transect California - Philippines (Yoshimoto &Gressitt, 1960), Japan (90 km from Kyushu (Yoshimoto & Gressitt, 1961), Tokyo Bay and Tanegashima (Holzapfel & Perkins, 1969), the Philippines (3 km from Luzon, 5 km from Mindoro, 10 km from Samar; Holzapfel & Perkins, 1969), 540 km from Wake Island, Guam, Taiwan and Vietnam (Holzapfel & Perkins, 1969). They have been even recorded on Antarctic waters (Gressitt et al., 1960; Gressitt et al., 1961).
The other Diptera family identified was Milichiidae, which correspond to a very specialized group with many species with kleptoparasitic behaviors (Albu, 2006). The one from this study was collected on the deck at 92 km from the mainland. Previously on 1959, the U.S.N.S Barrett recorded the family Milichiidae on different locations across the transect San Francisco-Guam-Japan-Okinawa-Korea-Japan-Honolulu. One specimen was collected at latitude 33°N at 200 km from Amami-Oshima (Japan). Two days after, on the bridge of the same vessel, more than 50 Milichiidae were found alive at 100 km from South Korea and 500 km from the China coast (Yoshimoto &Gressitt, 1960). Other records from the family appear at 50-100 km from South Korea and at 100 km from Kauaʻi (Yoshimoto& Gressitt, 1961). A piece of thorax identified as potentially Milichiidae was recorded on Antarctic waters in the latitudinal transect between 43°- 47°S in the south of New Zealand (173°E) (Gressitt et al., 1961).
There are also other two specimens of non-identified families of Nematoceran flies. One of those individuals corresponds potentially to Scatopsidae, and the other to either Chironomidae or Ceratopogonidae. The family Chironomidae has been previously reported as part of the aerial plankton (Gressitt et al., 1960; Gressitt et al., 1961; Yoshimoto & Gressitt, 1961), yet not Scatopsidae. The Chironomidae is the second most abundant family on freshwater ecosystems on the island (Gutiérrez-Fonseca et al., 2013).
The Hymenoptera family Agaonidae (superfamily Chalcidoidea) was less frequent on the reviewed literature. This superfamily is present on Isla del Coco (Hogue & Miller, 1981) and the one from this study was collected 325 km from the mainland on the way to the island (167 km from the island). Therefore, most likely it was carried by wind currents. There is only one other record, 5 km from the coast of Japan (34°30’N, 145°18’E) (Holzapfel & Perkins, 1969). These wasps are Ficus (Moraceae) pollinators, and on the island there is one species of these trees (Ficus petusa) (Trusty et al., 2006).
The Coreidae (Hemiptera) reported on this study was found on the deck at 23 km from the coast. Its proximity to the continent and almost absent presence on other surveys make it very likely that the insect was carried on the boat from land. The only other record of this group corresponds to a head and an abdomen potentially identified as belonging to Coreidae found in Antarctic waters in the latitudinal transect between 43°-50°S in the south of New Zealand (173°E) (Gressitt et al., 1960). This family is absent from Isla del Coco (Hogue & Miller, 1981).
The other family of Hemiptera recorded, Scutelleridae or shield-backed bug, has not been previously reported on this kind of aerial collecting. On this study, it was collected at 18 km from the mainland on the way back from the island at sunset time. Therefore, probably the insect was attracted to the lights of the boat. This record provides evidence of the minimum distance that a shield-backed bug can be dispersed to the open sea.
The Sphingidae moths were found at 30 km and 12 km, from the continental coast at nighttime. They correspond to 2 different species one in the genus Aellopos, and Nyceryx ericea (Druce, 1888), respectively. Most likely they were attracted to the lights of the boat powering themselves to that point. Long distance migrations on Lepidoptera have been widely documented and many of them reach continental scales as the Monarch (Danaus plexippus; Urquhart & Urquhart, 1978) in North America or the Danainae in India (Euploea and Tirumala; Kunte, 2005), so it is not unexpected for a robust month to cover long flight distances. This record presents evidence of the potential home range of these moths, implying that regularly they do offshore incursions of several kilometers. This family of Lepidoptera is present on the island: Agrius cingulata, Erinnyis obscura, Pachylia ficus, and Xylophanes tersa (Hogue & Miller, 1981; Montoya, 2015), yet not the genera collected.
No spiders were found on this collecting regardless they have been found in other trans-oceanic trips (Gressitt et al., 1960; Yoshimoto & Gressitt, 1960; Gressitt et al., 1961; Yoshimoto & Gressitt, 1961; Holzapfel & Perkins, 1969) and they are very well known of dispersing through ballooning. Most likely this absence is due to small sampling time.
The present research shows that regardless of the small sample set, the aerial plankton of the Eastern Tropical Pacific includes similar taxa to some of the ones recorded for the rest of the tropical Pacific. Moreover, one family (Ceratopogonidae, Diptera) and one Superfamily (Chalcidoidea, Hymenoptera) of the specimens collected by nets have representatives on Isla del Coco, which could be considered as evidence for a potential colonization mechanism by wind dispersal. Here, we also reported insects which most likely reached the boat by their own means providing evidence of their dispersal abilities. However, due to the small sampling size, this must be considered only as a preliminary result. Adding more replicates on different seasons and additional maritime routes will give a more complete view.
The combination of these field observations with our knowledge on the island community will allow us to answer more general questions about the island colonization process. In particular, does island colonization occurs due to a constant low probability over time, or due to rare events as hurricanes or storms? If the favored mechanism is the first one, we would predict to find representatives of most of the island native species as part of the aerial plankton. While, in the second case, it will require to invoke rare events that we might have not witnessed in historical times.
Ethical statement: authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.

