











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
Currently, there is a global concern regarding the state of populations of sharks and rays (Subclass Elasmobranchii), which are threatened across different geographic regions as a consequence of overfishing (Davidson, Krawchuk, & Dulvy, 2015) and degradation of marine ecosystems (Lotze et al., 2006). As exploitation increases globally, shark conservation and population management has become crucial for fishery researchers and administrators (Heithaus, Frid, Wirsing, & Worm, 2008). Sharks are highly vulnerable to overfishing because of their biological characteristics such as slow growth rates, late maturity, and low fecundity (Hoenig, & Gruber, 1990), which in turn causes these species to have a low rate of intrinsic population increment (Barker, & Schluessel, 2005). Sharks are also top predators that play an essential function for the balance of trophic chains and marine ecosystems (Myers, Baum, Shepherd, Powers, & Peterson, 2007).
The smalleye smooth-hound shark, Mustelus higmani, is a relatively small tropical species of demersal habitats that lives in the margins of continental and insular platforms (Springer, & Lowe, 1963). Its distribution is restricted to the Western-central Atlantic Ocean, from the Gulf of Mexico to Southeastern Brazil, including Colombia, Venezuela, and Trinidad and Tobago (Compagno, Dando, & Fowler, 2005). Mustelus higmani is a viviparous placentotrophic species with an annual reproductive cycle; the litter size varies between 2 and 9 pups, and feeds primarily on crustacea, small bony fish and squid (Compagno et al., 2005; Tavares et al., 2010). In Venezuela, this species is particularly abundant in the Northeastern region, and is the most frequently captured in artisanal shark fishery (40.8-36.2 %, Tavares et al., 2010; Márquez, Tavares, & Ariza, 2019). Despite the commercial importance of M. higmani, this species has not been well studied in the region, and there is a lack of information regarding its biology and fishery, which has hindered the stock assessment. According to the International Union for Conservation of Nature (IUCN), this species is globally listed as “Least Concern” (LC) (Faria, & Furtado, 2006); however in Venezuela, it is classified as “Vulnerable” (VU) (Tavares, 2015) due to the decreasing pattern in abundance and excessive fishing without any management and conservation measures.
Information about age structure, growth parameters, maturation processes and longevity are vital for stock assessment and prediction of population changes over time (Goldman, Cailliet, Andrews, & Natanson, 2012). To date, there are no studies on growth biology of M. higmani; however, age and growth have been evaluated in other species of the genus Mustelus, such as M. californicus and M. henlei in the Western coast of the United States (Yudin, & Cailliet, 1990), M. asterias in the Northeastern Atlantic (Farrell, Mariani, & Clarke, 2010), and M. mustelus in the Northwestern Atlantic (Conrath, Gelsleichter, & Musick, 2002). In this sense, the purpose of our study was to evaluate the age and growth of M. higmani from the Northeastern region of Venezuela, on the basis of the optical analysis of vertebrae and quantification of growth rings.
Materials and methods
Study area: The study area comprises a wide geographic extension of the Northeastern region of Venezuela (10° 35’-10° 38’ N & 65° 50 - 61° 51’ W; Digital Appendix 1), which corresponds to the zone explorer by the artisanal fishing fleets based in Margarita Island. The fishing area extends from the continental coastline of the Northeastern region of Venezuela to approximately 50 km North of Margarita Island and from the surrounding area of the La Tortuga Island to the limits with Trinidad and Tobago. The artisanal shark fishing fleets from Margarita Island are composed mainly of wooden boats propelled by outboard or central motors, and uses mainly gillnets and long lines as fishing gears (Tavares et al., 2010).
Size structure: In order to characterize the size composition of M. higmani specimens caught in the study area, we analyzed the shark fishing database (period 2007-2017) organized by the Instituto Nacional de Investigaciones Agrícolas (INIA) from Margarita Island. Size distribution by sex was described by creating a frequency histogram, grouping specimens in classes of 2 cm of total length (TL). Sex proportion (M:F) was evaluated statistically by using a c2-test in R (R Development Core Team, 2016). The mean birth size was estimated based on all neonates recorded that could be easily identified by the presence of an open umbilical scar.
Age and growth: Between August 2016 and July 2017, M. higmani specimens were examined in the fishing port of Juan Griego, one of the most important fishing markets in Margarita Island. For each specimen, the total length (TL cm; taken with the caudal fin in natural position) and sex were recorded at the time of samples collection. A vertebral segment (containing 6-8 vertebrae) was removed from under the origin of the first dorsal fin and the sections were taken to the laboratory and stored at -15° C. Preparation of the vertebrae, cleaning, cutting sections and quantification of the growth rings were performed following the procedures described by Goldman et al. (2012). Dried vertebrae were sagitally sectioned through the focus with a low-speed Isomet rotary diamond saw (Buehler). Vertebrae sections 0.5 mm thick, were then polished with wet fine-grit sand paper (No. 1 200) to a thick of 0.3 mm. The optical examination of the vertebral sections and measurements were performed based on digital images generated from a binocular dissecting microscope (Carl Zeiss 150C) with an integrated camera (Sony DCR-VX 2100). Growth patterns of the vertebrae consist of a wide translucent band followed by a narrow opaque band; together, they represent a growth ring. Visualization of the growth rings was achieved by modifying the brightness and contrast in the digital images by using the software Adobe Photoshop CC.
The periodicity of growth ring formation was evaluated using the index of Relative Marginal Increment (RMI; Natanson, Casey, & Kohler, 1995):
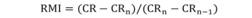
where, CR is the radius of the vertebral centrum, CRn is the distance between the focus and the last ring formed, and CRn-1 is the distance between the focus and the penultimate ring formed. Monthly averages of RMI were plotted against the size of the specimens (TL) to verify whether they showed a yearly growth pattern. Small individuals (neonates) were excluded from the analysis because they do not have a completely formed growth ring. We applied an analysis of variance (ANOVA; R Development Core Team, 2016) to statistically compare the monthly averages of RMI.
Growth parameters for M. higmani were estimated for both sexes by applying the von Bertalanffy growth function (VBGF; von Bertalanffy, 1938):
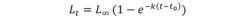
where, L t is the expected length at age t, L ∞ is the theoretical asymptotic length, k is the growth rate, t is the time in years; and t 0 is the theoretical age at which Lt would be zero. Additionally, the parameterized VBGF proposed by Ogle & Isermann (2017) was chosen in order to estimate the age at maturity for both sexes, and on the basis of the lengths at maturity (males: 51.4 cm TL; females: 50.0 cm TL) reported by Macias (2017). The parameterized VBGF is as follows:
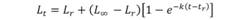
where, L r is the length required for fish in a population to reach a specific age (t r ). In this study t r represent the age at maturity for the lengths of L 51.4 (males) and L 50.0 (females). Fitting of models was conducted by using a least square method in an Excel spreadsheet with support of the Solver tool (Microsoft® Office Excel, 2015). Comparison of growth curves between the sexes was conducted by applying the analysis of the residual sum of square (ARSS; Chen, Jackson, & Harvey, 1992). This method is based on the Fisher distribution, and the F-statistic is calculated using the following equation:
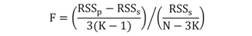
where, RSSp is the residual sum of square of each VBGF fitted by pooled growth data, RSSs is the sum of the RSS of each VBGF fitted to growth data for each individual sample, N is the total sample size; and K is the number of samples in the comparison. The theoretical longevity for males and females was calculated according to the equation proposed by Fabens (1965), which is defined by the time it takes to reach 95 % of L ¥and is denoted as t 0.95 = 5.0 (ln 2)/k.
Results
Size structure: Analysis of the shark fishery database (2007-2017) showed that M. higmani captures consisted of 2 820 males (22.1-62.8 cm TL) and 2 837 females (24.8-72.5 cm TL) (Fig. 1). Size composition indicated that captures are dominated by males between 48 and 56 cm TL and females between 46 and 60 cm TL. Results also showed that females have large amplitude in size intervals, with maximum lengths greater than males. However, the general M-F proportion observed (1:1.1) does not differ significantly from the expected proportion (1:1) (c2-test; P = 0.67). In total, 59 neonates (22.1-31.9 cm TL) were recorded in the study area, and the estimated average size at birth was 26.8 ± 4.2 s.d. cm TL.
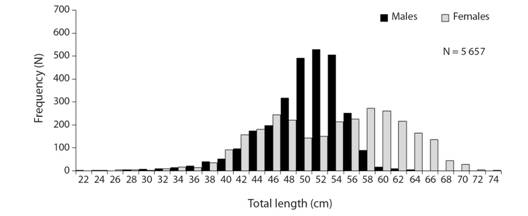
Fig. 1 Size structure for male and female Mustelus higmani caught in the artisanal shark fishery from the Northeastern region of Venezuela, period 2007-2017.
Age and growth: Optical analysis and quantification of the growth rings of M. higmani was based on 238 vertebral samples. Of these, 86 were from males (24.1-59.5 cm TL) and 152 were from females (24.4-69.5 cm TL). Analysis of RMI showed a high seasonal variability but with an apparent increasing trend from January to October, followed by a fall in RMI averages during the end of the year, from October to December. Then, RMI data suggested a yearly ring-formation pattern with October-December being the months when ring formation is completed (Fig. 2). Statistical analysis indicated a highly significant difference between monthly averages of the RMI (ANOVA; F = 3.67, P < 0.000). There was no detected overlap in the last growth rings of the vertebral sections from the largest individuals of both sexes, probably due to the low number of animals with sizes near the maximum length observed. In the vertebral sections examined, time of birth was determined by an angle change on the margin of the corpus calcarium. Formation of rings before birth was only identified in a few examined specimens (7.0 %).
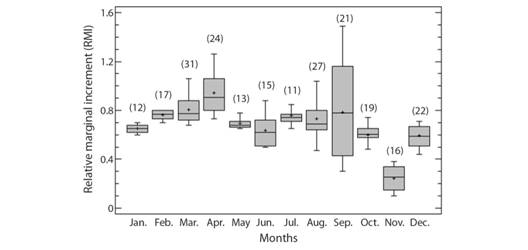
Fig. 2 Monthly averages of the Relative Marginal Increment (RMI) of Mustelus higmani from the Northeastern region of Venezuela. Numbers in parenthesis indicate sample size.
Taking into account the growth pattern observed in this study, the ages of M. higmani ranged between 0 and 5 years in males and between 0 and 6 years in females (Fig. 3). VBGF parameter estimates were L ¥ = 60.4 cm TL, k = 0.53 years-1 and t 0 = -1.02 years for males, and L ¥ = 71.1 cm TL, k = 0.38 years-1 and t 0 = -1.17 years for females. The ages at maturity (t r) derived from the parameterized VBGF were 2.6 and 2.1 years for males and females, respectively (Table 1). A highly significant difference was found between growth curves of males and females (ARSS test; F = 42.41, P < 0.000). Comparative information of the growth parameters reported for several species of the genus Mustelus from other geographic regions is shown in Digital Appendix 2. Longevity estimates were 6.6 years for males and 9.0 years for females. Based on the length/age at maturity estimates, we determined that commercial captures of M. higmani during 2007-2017 consisted of high proportions of immature specimens in both sexes (males: 59.9 %; females: 37.2 %).
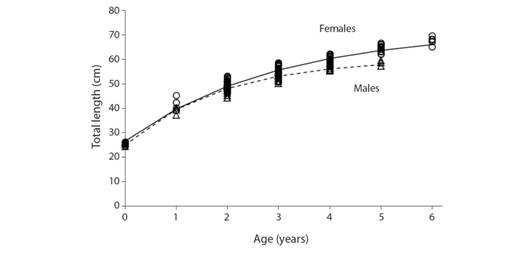
Fig. 3 Growth curves (VBGF) for male and female Mustelus higmani from the Northeastern region of Venezuela.
TABLE 1 Estimated VBGF parameters and derived variables for male and female Mustelus higmani from the Northeastern region of Venezuela
| Model | Sex | Parameter/Variable | RRS | ||||
| L ∞ | k | t 0 | L r | t r | |||
| VBGF | Males | 60.4 | 0.53 | -1.02 | - | - | 201.42 |
| Parameterized VBGM | 60.4 | 0.53 | - | 51.4 | 2.6* | 201.42 | |
| VBGF | Females | 71.1 | 0.38 | -1.17 | - | - | 506.47 |
| Parameterized VBGM | 71.1 | 0.38 | - | 50.0 | 2.1* | 506.47 | |
*Predicted ages at maturity (t r ) corresponding to the lengths (L r ) of 51.4 (L 51.4) and 50.0 cm (L 50.0).
Discussion
The size structure of M. higmani reported in the present study is similar to the pattern observed previously for this species in the Northeastern region of Venezuela (Macias, 2017; Márquez et al., 2019). The differential pattern by sex in relation to maximum sizes (i.e. females larger than males) as observed for M. higmani is common in species of the genus Mustelus (Conrath, & Musick, 2002; Sidders, Tamini, Pérez, & Chiaramonte, 2005; Farrell et al., 2010; Márquez et al., 2019), as well in other groups of sharks (Hoenig, & Gruber, 1990; Cortes, 2000). Regarding results on the general sex proportion of the specimens captured in artisanal fishery, we did not find a significant difference between sexes; however, a previous seasonal analysis by Macias (2017) revealed the presence of sexual segregation during some periods of the year. Sex segregation has also been widely described for elasmobranchs, and its origin may be related to several factors such as reproductive strategies, philopatric behavior, migratory patterns, nutritional requirements, among others (Sims, 2005; Wearmouth, & Sims, 2008; Heist, 2012). Analysis of size composition also revealed that specimens with sizes between 22 and 40 cm TL are not well-represented in commercial captures because this group was out of the action of fishing gears. The birth size estimated in this study (26.8 cm TL) is comparable to that previously reported (25.0 cm TL) by Tavares et al. (2010).
The formation of wide translucent and narrow opaque bands reflects periods of rapid and low growth, respectively (Loefer, & Sedberry, 2003) and generally, the mineralization pattern is associated with seasonal environmental changes and food availability (Goldman, 2004; Smith, Miller, & Heppell, 2013). The RMI analysis suggested an annual periodicity in the formation of growth rings of M. higmani in the study area. This finding agrees with the yearly ring-formation periodicity reported for other species of the genus Mustelus (Yudin, & Cailliet, 1990; Conrath et al., 2002; Farrell et al., 2010). Likewise, results showed that annual growth rings complete their formation by the end of the year, between October and December. This period precedes to the coastal up-welling season that affects the Northeastern coast of Venezuela, and extends throughout the first semester of the year (Castellanos, Varela, & Muller-Karger, 2002). This oceanographic phenomenon contributes to the increase of primary productivity and abundance of marine resources, thus increasing food availability for the sharks. Our results suggest that this time period of high prey abundance would correspond to the fastest growth phase of M. higmani in the study area. Although the births of M. higmani can be observed throughout the year, data on the seasonal development of embryos and follicles, and the numbers of captured neonates indicate that time of parturition and mating still occur during a main period of several months, between November and February (Tavares, unpublished data). The pattern related to the embryo development and time of parturition could explain the lack of growth rings before birth in most of the examined specimens because only those embryos that were in the early development process during the months of the main birth period would have pre-birth growth rings.
Based on the vertebrae analysis, the maximum ages of M. higmani were 5 years for males and 6 years for females. Because of the low number of specimens analyzed with sizes near of the known maximum lengths, it will be necessary to complement the growth studies of this species to validate our results and to avoid an underestimation of maximum age as a consequence of overlap of the last growth rings formed in larger animals. The overlap of growth rings happens because larger individuals increase in age, but their growth rates gradually decrease with time, resulting in the superposition of the rings, thus affecting age quantification.
Regarding VBGF parameters, the asymptotic length values for males (L ∞ = 60.4 cm TL) and females (L ∞ = 71.1 cm TL) are comparable to the maximum sizes registered for M. higmani in the study area. The growth rate estimates for males (k = 0.53 years-1) and females (k = 0.38 years-1) can be considered high even among species of the genus Mustelus. The growth rates reported for other Mustelus species ranged from 0.20 and 0.44 years-1 in males and from 0.15 and 0.29 years-1 in females (Yudin, & Cailliet, 1990; Conrath et al., 2002; Farrell et al., 2010; see Digital Appendix 2). According to Branstetter (1990), Mustelus species are within the group of sharks with small sizes (< 100 cm) and fast growth (k>0.1). In relation to the difference in growth rates between sexes (generally favoring males), this is a biological characteristic among shark species (Hoenig, & Gruber, 1990). The growth rate and the asymptotic length are variables negatively correlated, indicating that larger individuals will likely have a lower growth rate. These lower growth rates displayed by females may be explained by the need for females to attain larger sizes to support pups and to a smaller investment in growth in favor of reproduction (Cortes, 2000).
Our results showed that the maturation process of M. higmani is relatively rapid and similar for both sexes, with an age at maturity of 2.6 in males and 2.1 in females. These estimations are similar to those reported for M. californicus (males: 1.5 years, females: 2.5 years; Yudin, & Cailliet, 1990) and slightly lower than those obtained for M. henlei (males: 3.0 years, females: 2.5 years; Yudin, & Cailliet, 1990) and M. canis (males: 3.0 years, females: 4.5 years; Conrath et al., 2002). The longevity estimates for M. higmani were 6.6 years in males and 9.0 years in females. Considering the longevity values for elasmobranchs provided by Cailliet and Goldman (2004), M. higmani is among the species with the shortest longevity, even among sharks of the genus Mustelus.
Artisanal shark fisheries in Venezuela represent a traditional commercial activity that is conducted along all coasts and islands, and as the most Caribbean nations that harvest this fishing resource, there is an uncertainty in relation to fishery data and biological information of most shark species. Mustelus higmani is one the shark species with greatest commercial importance in the Northeastern region of Venezuela, and in spite of this, it does not have any management and conservation plans. Because of this, it is necessary to contribute with knowledge and scientific data to evaluate the population and design the conservation measures. Despite the high mortality levels to which M. higmani is subjected in the study area, that includes large numbers of immature individuals, its population has probably not collapsed because of the biological characteristics of this species such as fast growth, early maturity and annual reproductive cycle. To conclude, this is the first study that evaluates age and growth of M. higmani through its distribution range. However, it is necessary to continue and complement the studies regarding the most crucial aspects of this species, for example growth process, age structure and reproductive biology, among others. Likewise, it is recommended that government institutions with responsibility in fishery administration and conservation implement measures that lead to the sustainable exploitation of this species, such as the establishment of minimum capture sizes based on sexual maturity, and changes in the characteristics of fishing gears, mainly gillnets.
Ethical statement: authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.

