











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
The United Nations Convention on Biological Diversity defines “indigenous” as those people who have historical continuity with pre-invasion and pre-colonial societies, that have developed on their own territories, and who consider themselves distinct from other sectors of society now prevailing in those territories. To raise awareness of their importance for humanity, the United Nations designated 2019 as the “International Year of Indigenous Languages.”
Biocultural diversity encompasses the link between biological diversity and humankind’s cultural diversity, and identification of ecological conservation hotspots and linguistics are the two pillars of biocultural diversity analyses. Trends in biological diversity and cultural/linguistic diversity parallel one another (Maffi, 2001, Maffi, 2005) and there is a documented correlation between the two (Mühlhäusler, 1995; Harmon, 1996; Nettle & Romaine, 2000; Oviedo, Maffi, & Larsen, 2000; Moore et al., 2002; Sutherland, 2003; Stepp et al., 2004; Fincher & Thornhill, 2008; Gorenflo, Romaine, Mittermeier, & Walker-Painemilla, 2012). Research across both continental and regional scales have identified patterns of co-occurrence of linguistic and biological diversity around the world, and nations/areas with high biodiversity also tend to have high linguistic and cultural diversity (Toledo, 1994; Harmon, 1995, Harmon,1996; Nettle, 1996; Nettle & Romaine, 2000; Stepp et al., 2004; Loh & Harmon, 2005; Toledo & Barrera-Bassols, 2008; Gorenflo et al., 2012). The countries with the highest linguistic diversity (jointly having 54 % of all living languages) are Papua New Guinea, Nigeria, India, Mexico, Cameroon, Australia, the Democratic Republic of the Congo, and China (Harmon, 1995); and the most bioculturally diverse countries as measured by their centers of biological, linguistic, and agricultural diversity, as well by the presence of indigenous peoples, include those plus Indonesia, Brazil and Peru (Toledo & Barrera-Bassols, 2008).
The conceptual framework of biocultural diversity draws upon a common interest in understanding and preserving the relationships between biological, linguistic, and cultural variety and range. For indigenous people, biological, cultural, and linguistic diversities are intrinsically linked, as are environment and development. Biocultural diversity is often used as an index, or measure, to assess geographical regions in terms of the status or linkages between biological, cultural and linguistic diversity (Harmon, 1996; Sutherland, 2003; Harmon & Loh, 2010; Gorenflo et al., 2012). Languages, like genes, are “documents of history,” and a vast amount of information about our past is inscribed in the content and structure of the approximately 7 100 languages that are spoken today (Gray, Quentin, & Greenhill, 2018). Present-day indigenous cultures and languages are an expression of the long historical legacy of interrelationships between humans and nature (Toledo & Barrera-Bassols, 2008). Conservation practitioners should also embrace biocultural approaches for social justice, legal, and practical reasons (Gavin et al., 2013, Gavin et al., 2015). This view was convincingly articulated in the Indigenous Peoples International Declaration on Self-Determination and Sustainable Development at the 2012 United Nation’s Rio+20 Summit, which reaffirmed that cultural belief systems and worldviews of indigenous people are fundamental to biodiversity protection and sustainable development.
Species are the basic units of biodiversity, while languages serve to measure the diversity of cultures, and the striking parallels in their evolution suggest that nature and culture evolved in similar fashion (Harmon, 1996). In practical terms, there is an emphasis on language over other aspects of culture and identity because it strongly circumscribes an indigenous group. Homo sapiens displays a remarkable linguistic diversity that correlates strongly to areas of plant and animal diversity-and both may be moderated by similar environmental factors, such as temperature and rainfall. Both domains also involve the transmission of discreet heritable units: genes in biology, and socially-transmitted units such as words and morphosyntax in linguistics. Both can be altered as they pass through generations and thus, they display a hierarchical relationship over time. Languages, like biological species, are therefore related through nested patterns of descent allowing their evolutionary history to be depicted in branching patterns, or trees (Dunn, 2014). Of course, some words do enter languages by way of diffusion, or “borrowing” from other cultural sources (Campbell, 1997), and these instances are comparable to horizontal gene transmission in the scheme of biodiversity assessment and phylogenetics. Nonetheless, just as the historical evolution of species can be inferred by phylogenetic analysis, so the history of linguistic diversity can be estimated by phylogenetics (e.g., Bouckaert et al., 2012; Birchall, Dunn, & Greenhill, 2016; Gray et al., 2018). In both cases, the phylogenies can be overlain by areas of endemicity to produce dated phylogeographic reconstructions. In the case of cultures and linguistics, this amounts to what has been called “virtual archeology.”
Three types of language diversity have been recognized (Gavin et al., 2013). Language richness refers to the number of languages within a given area. Phylogenetic language diversity is the minimum total length of all branches needed to span a set of “language taxa” on a phylogenetic tree. Linguistic disparity refers to the range of expression in a language trait within a clade. Languages differ on a multitude of structural levels, including phonology, morphology, and syntax. There is a strong latitudinal gradient in both language richness and biological richness (Gavin et al., 2013, Gavin et al., 2015).
It has been proposed that human languages reached their maximum number (estimated at 12 000) at the end of the Pleistocene, directly predating the rise of agriculture (Harrison, 2007). In the non-sedentary hunter-gatherer societies of the time, the dominant force in language creation is likely to have been fissioning mechanisms (Hamilton, Milne, Walker, Burger, & Brown, 2007), although such historical “splits” might be blurred by inter-language diffusion, or borrowing, as noted above (e.g., Campbell, 1997). The number of languages has been in decline since the Neolithic as agricultural groups have spread, replacing hunter-gatherers, and population movements have tended to reduce global language diversity (Nettle, 1999a, Nettle, 1999b). A substantial body of theoretical work suggests that the spread of politically complex agricultural societies was a dominant factor in the reduction of language diversity (Renfrew, 1994).
After examining nearly 7 000 languages spoken on Earth today against the planet’s biological diversity and biodiversity hotspots, Gorenflo et al. (2012) found that more than 4 800 languages occur in regions containing high biodiversity, and 3 202 languages occur in 35 previously defined biodiversity hotspots. Of the languages found in biodiversity hotspots, 1 553 are spoken by 10 000 or fewer people, and 544 are spoken by 1 000 or fewer people.
Greater geographic heterogeneity, as measured by more-diverse habitats or higher topographic complexity, is commonly correlated with greater species diversity (e.g., Kerr & Packer, 1997). A link between geographic heterogeneity and diversity also appears to exist for languages. Environmental variables have been noted as important predictors of linguistic diversity patterns, and historical processes of co-evolution of small-scale human groups with their local ecosystems have been proposed (Harmon, 1996; Maffi, 1998).
Three narratives have been offered to explain the correlation between biological and cultural diversity (Maffi, 2001): geographic determinism (both species and languages diversify in heterogeneous landscapes with geographic barriers); ecological determinism (linguistic diversification occurs in response to high biodiversity as cultural groups encode their knowledge of rich biotas); and historic determinism (areas of high linguistic diversity today are “residual,” persisting because of their geographic isolation from extensive agricultural development, implying that now-impoverished areas were once more diverse).
Today, linguistic diversity and biological diversity face similar threats and are both in crisis because of human population growth and increasing consumption (Loh & Harmon, 2014). One of the biggest challenges for biocultural conservation is ensuring that indigenous peoples obtain recognition of their rights to the resources found on their lands and territories on which they depend on for their economic, spiritual, cultural, and physical well-being (Inter-American Commission on Human Rights, 2009), and that these translate into tangible policies and action at the national and international level. Which is, by no means, an easy task since it has been estimated that about 12-20 % of areas under human management worldwide are indigenous lands (Toledo, 2001a).
Different cultures perceive and value biodiversity in different ways because of their distinct heritage and experience. Posey (1999) argued that cultural diversity parallels ecological diversity, and local traditional adaptations are often the most environmentally sound. Through time, as local communities interact closely with their environment and modify it as they adapt to specific ecological niches, they acquire detailed knowledge of the environment and how to manage it for their long-term benefit. Languages thus carry deep eco-cultural knowledge, and embedded in indigenous languages is knowledge about habitats, plant life, animal behavior, conservation methods, and many other aspects of the natural world (Harrison, 2007).
The world’s inhabitants today speak around 7 100 languages, patterned unevenly across the Earth. But, roughly half of the world speaks only 24 languages, and have from tens to hundreds of millions of speakers, while the other half of the population speaks the remaining 7 073 languages, of which around half have fewer than 10 000 speakers (Lewis, Simons, & Fennig, 2016; Simons & Fennig, 2018). Since 1950, 230 languages have disappeared and at least 3 000 have become endangered, of which 577 are critically endangered (78 have 10-50 speakers, and 146 have fewer than 10 speakers) (United Nations Educational, Scientific and Cultural Organization [UNESCO], 2010). And, according to UNESCO, nearly 40 % of the world’s population lack access to education in a language that they speak or understand. In the next 30 years, over half of the world’s languages will likely go extinct or be spoken by only a few old people (Harrison, 2007).
While much effort has and is being spent to stem the loss of plant and animal species, the loss of languages is receiving far less attention and is actually being ignored in many parts of the world, particularly in developing countries, for a variety of reasons. As the world grows less linguistically and culturally diverse, it is also becoming less biologically diverse. Biologists estimate an annual loss of species at 1 000 times or greater than historic rates, and linguists predict that 50-90 % of the world’s languages will disappear by the end of this century (Nettle & Romaine, 2000; Gorenflo et al., 2012).
More than 70 % of the world’s biodiversity is found in only 17 countries, of which 15 are in the developing world and 6 are in Latin America (Mittermeier, Myers, Gil, & Mittermeier, 1999). The most species-rich, human-accessible environments are tropical rain forests and deciduous forests, coral reefs, and large tropical lakes (Millennium Ecosystem Assessment [MEA], 2005). Even though the world’s tropical forests occupy only 7 % of the land area, they have been estimated to contain over half the world’s species (Oviedo et al., 2000; Corlett & Primack, 2010). Of the 93 579 species of plants and animals assessed by the Red List of the International Union for the Conservation of Nature (IUCN) until 2018, more than 26 000 (28 %) are threatened with extinction, including 41 % species of amphibians, 35 % of reptiles, 25 % of mammals, 13 % of birds, 7.5 % of bony fishes and 63 % of cycads. Those estimates, however, are very conservative given that the species evaluated only represent about 4 % of the nearly two million living species described to date (Brusca, Moore, & Shuster, 2016; Primack & Vidal, 2019), and far less than the estimated total number of species on Earth (estimates range from around 10 to 100 million species of prokaryotes, protists, plants, fungi, and animals) (Mora, Tittensor, Adl, Simpson, & Worm, 2011; Brusca et al., 2016). At least 479 species of vertebrates and 116 species of plants have gone extinct in modern times, due mainly to habitat destruction and fragmentation, overexploitation, and invasive species. Although the global extent of protected land has roughly doubled in size since the 1992 Earth Summit in Rio de Janeiro, with more than 202 000 protected areas now covering 14.7 % of the world’s terrestrial area, 6 × 106 km2 (32.8 %) of that protected land globally is under intense human pressure (Jones et al., 2018).
In this paper, we test the hypothesis that a pattern of co-occurrence of biological and cultural diversity exists in Mexico. We do this by examining the available data on species diversity and language diversity across all Mexican states, and by looking for areas of overlap between regions of high diversity and extinct/threatened/endangered diversity in species and languages.
Materials and methods
Much has been written on Mexico’s biodiversity (for a review see Sarukhán et al., 2017). However, little has been published in the peer-reviewed literature on its rich linguistic diversity. We extensively reviewed the available literature, including government reports, on Mexico’s biocultural diversity with a particular focus on endangered and critically endangered species and languages to look for overlapping geographical distributions and threats. Most of the information we use regarding the languages of Mexico comes from an exhaustive appraisal of reports by Mexico’s National Institute of Indigenous Languages (Instituto Nacional de Lenguas Indígenas [INALI], 2012) and official published government records (Diario Oficial de la Federación, 2008, Diario Oficial de la Federación, 2010a). From these sources, we compile a robust set of data, though much of it is qualitative (not quantitative).
Our ranking criteria for biological species follow the IUCN, and we consider a species criticallyendangered when it faces an extremely high risk of becoming extinct, and endangered when it faces a very high risk of becoming extinct (IUCN, 2018). We follow two closely aligned criteria for defining endangered languages. The first, more detailed criterion, from INALI (2012), considers that a language has very high risk of disappearance when the total number of speakers is less than 100, the number of child speakers is less than 25 %, and the speakers are less than 30 % of the population in all localities; and a language is at high risk of disappearance when the total number of speakers is less than 1 000 but higher than 100, the number of child speakers is less than 25 %, and the number of speakers in just one locality is less than 30 %. The second criterion considers a language critically endangered when the number of speakers is less than 50, and endangered when the number of speakers is less than 250 (Loh & Harmon, 2014; Simons & Fennig, 2018). Both of these criteria are in line with the degrees of endangerment established by UNESCO (2010).
A linguistic variant has structural and lexical differences when compared with other variants in the same linguistic group. And those that speak a variant have a sociolinguistic identity that contrasts with the sociolinguistic identity of those that speak other variants within the same linguistic group (Diario Oficial de la Federación, 2008). Those differences could be sounds, words, meanings, and uses that result in speakers of one variation being unable to understand speakers of other variations. Variants are therefore often seen as equivalent to languages (INALI, 2012) and are considered as such in this paper.
Results
Biological Species
Mexico harbors 10-12 % of the world’s biological diversity and is ranked as the fourth megadiverse country after Indonesia, Brazil and Colombia (Mittermeier & Mittermeier, 1992; Mittermeier et al., 1999; Toledo, Boege, & Carrera-Bassols, 2010; Sarukhán et al., 2017). So far, 118 030 (66.44 %) of the estimated 177 641 species of animals and vascular plants of Mexico have been described (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad [CONABIO], 2008; Sarukhán et al., 2017): 23 314 species of vascular plants, 564 mammals, 1 150 birds, 908 reptiles, 399 amphibians, 2 763 fishes (2 224 marine), 11 472 non-arthropod invertebrates, 11 185 non-insect arthropods, and 66 275 insects. These estimates include terrestrial, freshwater, and coastal marine species.
As in other parts of the world, tropical regions in Mexico contain more species than temperate areas; although there are certain groups that are particularly diverse in arid zones, such as cacti, of which at least 677 species are found in the country, 518 of them being endemic (Dávila et al., 2002; Villaseñor, 2016). The states of Oaxaca, Chiapas, Veracruz and Guerrero (all in the South/Southeast and close to Guatemala and Belize) and Michoacán are home to the bulk of the country’s biodiversity (see also Toledo et al., 2001). Mammal, reptile and amphibian species richness is much higher in Oaxaca, Chiapas, Veracruz and Tabasco (CONABIO, 2008) and although it varies among different groups, tropical humid regions appear to have fewer endemic species than arid and semiarid regions (CONABIO, 2016). Oaxaca alone is home to almost half of Mexico’s known species of vertebrates, 19 % of the known invertebrates, and 40 % of the known plants (Gonzalez-Perez, Briones-Salas, & Alfaro, 2004).
At least 127 species of Mexican flora and fauna (58 % of them endemic) are known to have gone extinct through 2008 (Table 1). However, since most recorded extinctions have occurred on islands and in continental lagoons and rivers, and include only flowering plants and vertebrates, the actual number of extinctions (including other habitats and taxa) is probably much higher. There are today 1 213 Mexican species (many endemic) threatened with extinction (Diario Oficial de la Federación, 2010b; IUCN, 2018; Supplemental Material): 94 (73 endemic) species of mammals, 66 (24) birds, 97 reptiles, 219 (182) amphibians, 181 fishes, 8 mollusks, 98 other invertebrates, and 450 plants (including 12 conifers, 41 cycads, 133 cacti and 22 magnolias). Unsurprisingly -given the extent of habitat destruction and fragmentation in Oaxaca, Chiapas, Veracruz, Guerrero, Michoacán, and states adjacent to the Gulf of California, including its many islands and islets- threatened hotspots are located mostly in those regions.
TABLE 1 Mexico’s recorded extinct species of fauna and flora (CONABIO, 2008; Sarukhán et al., 2017)
| Taxa | Extinct Species | Endemic Species | States/Islands |
| Plants | 26 | 5 | Hidalgo, Veracruz, Jalisco, Isla Guadalupe |
| Fish | 38 | 20 | Nuevo León, Coahuila, Jalisco, Durango, México, Veracruz |
| Amphibians | 29 | 29 | Oaxaca, Veracruz, Guerrero, Querétaro, Hidalgo, Durango |
| Birds | 19 | 11 | México, Sonora, Coahuila, Michoacán, Colima (Isla Benedicto), Baja California (Islas Todos los Santos), Isla Guadalupe, Isla Socorro |
| Mammals | 15 | 9 | Baja California (Islas Todos los Santos, Coronado, Turner, and San José), Nayarit (Archipiélago las Marías), Sonora (Isla San Pedro), Isla Ángel de la Guarda, Isla San Juanito |
| Totals | 127 | 74 |
Languages
Today, 25.7 million (21.5 %) of Mexico’s inhabitants are indigenous peoples, and 7.4 million (6.5 % of all Mexicans older than three years) speak an indigenous language (Instituto Nacional de Estadística y Geografía [INEGI], 2010, Instituto Nacional de Estadística y Geografía [INEGI] 2015); although in some states such as Chiapas, Oaxaca and Yucatán that percentage is nearly 30 % (CONABIO, 2016). Indigenous land represents approximately 14.3 % (or 28 million hectares) of the country’s total territory (Boege, 2008, Boege, 2009) and nearly half of the most important watershed headwaters are occupied by indigenous peoples. About a third of Mexico’s protected natural areas (at the federal level) have indigenous populations living within them, and approximately 70 % of Mexico’s indigenous land is under some sort of priority for the conservation of its rich biological resources (Toledo et al., 2010). There are 64 172 named localities with indigenous people and a quarter of Mexico’s social properties are located within 4 786 ejidos (communal land, which in Mexico is land expropriated from owners of large tracts and redistributed for shared use as farmland, especially to poor populations, in accordance with the Agrarian Reform Act of 1917) and 1 258 agrarian communities in territories own by indigenous people (INEGI, 2015). The states of Oaxaca, Chiapas, Veracruz, Guerrero, Hidalgo, Estado de México, and Yucatán are home to 77 % of the Mexico’s indigenous people, while the states of Coahuila, Colima and Zacatecas have the lowest indigenous populations.
Today, with 364 living languages, Mexico is the world’s fifth most linguistically diverse country after Papua New Guinea, Indonesia, Nigeria and India (Diario Oficial de la Federación, 2008; Toledo et al., 2010; INALI, 2012). Mexico’s indigenous languages belong to 11 linguistic families and 68 linguistic groups (Diario Oficial de la Federación, 2008; INEGI, 2010; INALI, 2012). This cultural richness is, however, threatened.
Four linguistic groups comprise the largest numbers of speakers in Mexico -Náhuas (1 376 000 speakers), Maya (759 000), Mixteco and Zapoteco (> 400 000)- while 22 groups each have less than 1 000 speakers. Oaxaca, Puebla, Chiapas, Veracruz and Guerrero are the states with the highest linguistic diversity; densities of languages in Oaxaca, Veracruz and Guerrero are comparable to areas in Papua New Guinea, the Himalayas, Nigeria, and Cameroon (CONABIO, 2008). Ninety percent of the people who speak the Cochimí-Yuman family (Cucapá, Paipai, Kumiai, Ku’ahl and Kiliwa) live in Baja California and Baja California Sur, all of which are at high or very high risk of disappearance. Cochimí is not a “modern” Yuman language, but one that likely split off from a Proto-Yuman tongue long ago (Mixco, 1978).
The status of speakers among ethnic groups differs greatly throughout Mexico. There are groups in which only the elders speak the language (in Mexico), as is the case of the Ixcateco, Ayapaneco, Kiliwa, Paipái, Cucapá and Ku’ahl; and those for which adults speak the language, but children do not, such as Tlahuica, Mocho’, Tuzanteco, Teko, Awakateko, Oluteco, Ayapaneco, Texistepequeño, Chocholteco, Kaqchikel, Ixil, and Ixil chajuleño (INALI, 2012). However, Kaqchikel and Ixil are languages probably represented today in Mexico by relatively recent immigration from Guatemala, where they remain strongly represented. At least 13 languages have gone extinct in Mexico: Pericú, Solteco, Naolan, Opata, Pochuteco, Cuitlatec, Pame (Southern), Tepecano, Tubar, Chiapaneco, Eudeve, Pochuteco and Cochimí. Of the 364 languages spoken today in Mexico, 64 are in very high risk of disappearance (less than 100 speakers survive) and 43 are in high risk of disappearance (less than 1 000 but more than 100 speakers survive (INALI, 2012) (Table 2, Fig. 1). Nine languages in Mexico are also considered by Simons and Fennig (2018) to be critically endangered (less than 50 speakers survive), and 29 endangered (less than 250 speakers survive). The ones with the highest risk of going extinct in the immediate future are: Kiliwa (2 speakers survive), Awakateco (3 speakers), Tuzanteco (5 speakers), Ayapaneco (8 speakers), Ixil nebajeño (12 speakers), Zapoteco de Mixtepec (14 speakers), Ku’al (20 speakers), Ixcateco (21 speakers), Kaqchikel (35 speakers), Zapoteco de San Felipe Tejalapám (50 speakers), Ixil chajuleño (52 speakers), and Zapoteco de Asunción Tlacolulita (53 speakers). Mexico’s 56 extinct and endangered languages occur in 16 states, most (77 %) in Oaxaca (16), Baja California (7), Chiapas (6), Campeche (6), Sonora (4) and Quintana Roo (4) (Table 2, Table 3).
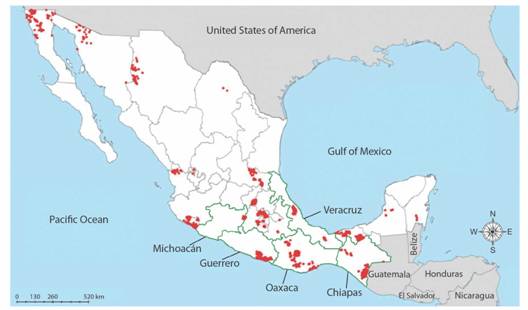
Fig. 1 Mexico’s languages at very high risk of disappearance shown in red (from INALI, 2012; coloring denotes areas in which the language is spoken), and its most biologically diverse states bordered in green.
TABLE 2 Extinct, critically endangered, and endangered languages in Mexico
| EXTINCT | |||
|---|---|---|---|
| Language (family) | Number of Speakers | State | Locality (municipality: villages) (1) |
| Pericú (2) | Baja California | Los Cabos region | |
| Solteco (Oto-mangue) (3) | None | Oaxaca | |
| Cuitlatec (4) | None | Guerrero | Balsas River |
| Naolan (unclassified) (5) | None | Tamaulipas | San Juan Naolan, near Tula |
| Pochuteco (6) | None | Oaxaca | Pochutla |
| Opata (Uto-azteca) (7) | None | Sonora | Arivechi, Bacanora, Nacori, Onavas, Sahuaripa, Suaqui. Tecoripa is the traditional area |
| Pame, sureño (Oto-mangue) | None | Estado de Mexico (possibly Hidalgo) | Jiliapan |
| Tepecano (Uto-azteca)(8) | None | Jalisco | Villa Guerrero: Azqueltán (earlier Atzqueltlán), San Martin de Bolaños on Rio Bolaños |
| Tubar (Uto-azteca) (9) | None | Chihuahua near de border with Sonora and Sinaloa | |
| Chiapaneco (Oto-mangue) | None (10) | Chiapas | El Bosque, Las Margaritas, Ocosingo, Alenque and Sabanilla municipalities |
| Eudeve (11) | None | Sonora (12) | |
| Pochuteco (Uto-azteca) (13) | None | Oaxaca | Pochutla |
| Cochimí (Cochimí-yumana) (14) | None (15) | Baja California Baja California Sur | Noth of Loreto |
| CRITICALLY ENDANGERED (less than 50 speakers and no children -5 to 14 years of age- speak the language) | |||
| Kiliwa (Cochimí-yumana) | 2 (16)* | Baja California | Ensenada: Arroyo de León (Ejido Kiliwas), Ejido San Francisco R. Serrano (Valle San Matías), Ensenada, Francisco Zarco (Guadalupe), Juntas Neji, La Zorra, Lázaro Cárdenas (Valle de la Trinidad), Licenciado Gustavo Díaz Ordaz, Parcela Número 19 (Familia Castro Ejido Nalta), Rancho las Pinzas |
| Awakateco (Maya) | 3* | Campeche | Champotón: Maya Tecún I, Maya Tecún II, Santo Domingo Kesté |
| Tuzanteco (Maya) | 5* | Chiapas | Tuzantán: Estación Tuzantán |
| Ayapaneco (Mixe-zoque) | 8 (17)* | Tabasco | Jalapa de Méndez: Ayapa, El Carmen (La Ensenada), Vicente Guerrero Primera Sección |
| Ixil nebajeño (Maya) | 12* | Quintana Roo | Othón P. Blanco: Maya Balam |
| Zapoteco de Mixtepec (Oto-mangue) | 14 (18)* | Oaxaca | San Cristóbal Amatlán: San Agustín Mixtepec |
| Ku`al (Cochimí-yumana) | 20 (19)* | Baja California | Misión de Santa Catarina, Comunidad Indígena de Santa Catarina, El Aguajito (Mat Chip) [Rancho Matchip], Rancho Escondido, Rancho Wikualpuk (El Ranchito), Ensenada |
| Ixcateco (Oto-mangue) (20) | 21 (21)* | Oaxaca | Oaxaca de Juárez, Santa María Ixcatlán |
| Kaqchikel (Maya) | 35* | Campeche Quintana Roo | Campeche: Quetzal-Edzná I, Quetzal-Edzná II, Los Laureles. Champotón: Maya Tecún II, Santo Domingo Kesté Quintana Roo. Othón P. Blanco: Kuchumatán, Maya Balam, San Isidro la Laguna |
| Zapoteco de San Felipe Tejalapám (Oto-mangue) | 50 (22)* | Oaxaca | San Felipe Tejalápam: Arroyo Central, Cañada Cera, Conejo (Rancho El Conejo), El Chilar, El Copalito, El Cucharal, El Paredón, El Roble, La Aurora, La Unión, Llano Ceniza, Llano El Coyote, Los Paderones, San Felipe Tejalapám |
| ENDANGERED (less than 250 speakers no children -5 to 14 years of age- speak the language) | |||
| Ixil chajuleño (Maya) | 52 | Campeche Quintana Roo | Campeche: Los Laureles, Quetzal-Edzná Quintana Roo. Othón P. Blanco: Kuchumatán |
| Zapoteco de Asunción Tlacolulita (Oto-mangue) | 53 | Oaxaca | Asunción Tlacolulita: Asunción Tlacolulita, Primera Sección Norte |
| Oluteco (Mixe-zoque) | 60* | Veracruz | Oluta: Correa de Abajo, El Chorro, El Mirador, El Ángel Gabriel, Los Laureles, Oluta, Tenejapa |
| Chocholteco del oeste (Oto-mangue) | 62 | Oaxaca | Ocotlán: Boquerón, San Antonio Nduayaco, Tierra Colorada, Unión Palo Solo. San Francisco Teopan: Concepción Buenavista, Santiago Ihuitlán Plumas, Tepelmeme Villa de Morelos, Tlacotepec Plumas. San Miguel Tequixtepec: Los Batos. San Miguel Tulancingo: Agua Dulce, Buena Vista, El Coatillo, El Español, Gasucho, Loma Larga, Rancho Marino Sánchez, San Miguel Tulancingo. San Pedro Nopala: San Mateo Tlapiltepec, Santa María Jicotlán. Santa Magdalena Jicotlán: San Mateo Tlapiltepec, Santiago Tepetlapa. Teotongo: El Progreso, El Tecomate, Guadalupe, La Luz |
| K`iche`(occidental) (Maya) | 65 | Campeche | Champotón: Santo Domingo Kesté |
| K`iche` (central) (Maya) | 65 | Campeche | Champotón: Santo Domingo Kesté |
| Teko (Maya) | 71* | Chiapas | Amatenango de la Frontera: Amatenango de la Frontera, Barrio Nuevo, Chiquisbil, El Porvenir, Granadillal, Las Marías, Nuevo Amatenango, Sabinalito. Frontera Comalapa: Nuevo Mazapa. Mazapa de Madero: Bacantón Altamirano Uno, Chimalapa, Mazapa de Madero, Nuevo Paraíso, Reforma, Tierra Blanca, Valle Obregón, Veracruz, Villa Hidalgo. |
| Zoque del sur (Mixe-zoque) | 84 | Chiapas | Ocozocuautla: El Edén, El Pedregal, Nuevo Chapultenango, Salina Cruz, San Antonio, San Luis las Maravillas, San Pedro, Velasco Suárez Uno. Tuxtla Gutiérrez: Copoya, Julio César Ruíz Ferro Segunda Sección, Lindavista, Los Crisantemos. |
| Pápago (Pima alto) (Yuto-nahua) | 94 | Sonora | Altar: Altar, El Cubabi, El Cumarito. Caborca: El Soñic, Heroica Caborca, Juárez, La Escondida, Las Norias (De Romero), Puerto Lobos, San Francisco Pápagos (San Francisquito), San Pedro, Santa Eduwiges (La Cachora), SPR Mayobampo (Rancho Corralero). General Plutarco Elías Calles: Ejido Luis Echeverría Álvarez, El Chamizal, El Desierto de Sonora, Quitovac, San Antonio, Santa Rosa (La Angostura), Sonoyta. |
| K`iche’ (oriental) (Maya) | 96 | Campeche Chiapas Quintana Roo | Campeche: La Libertad. Champotón: Maya Tecún II, Santo Domingo Kesté. Chiapas. Las Margaritas: San Mateo Zapotal. Marqués de Comillas: Boca de Chapul, Pico de Oro. Quintana Roo. Othón P. Blanco: Kuchumatán, Maya Balam, San Isidro la Laguna. |
| Kickapoo (Álgica) (23) | 105 | Coahuila | Múzquiz: Ciudad Melchor Múzquiz, El Nacimiento de los Kikapúes (Nacimiento de la Tribu Kikapoo) |
| Mexicano de occidente (Yuto-nahua) | 107 | Jalisco | Cuautitlán de García Barragán: Ayotitlán, Cortapico, Cuautitlán de García Barragán, Cuzalapa, Chacala, Chanquehahuil, El Chico, La Guaca, La Rosa, La Rosita, Los Encinos, Los Sauces, Los Terreros, Mojoneras, Paso Real, Plan de Méndez, Plan de San Antonio, Rancho Viejo, San Miguel, Santa Rosa, Tierras Blancas, Tierras Negras |
| Kuapá (Cucapá) (Cochimí-yumana) | 119 (24) | Baja California Sonora | Baja California. El Mayor Cucapá, Pozas de Arvizu. Mexicali: Campo Camerina (Colonia Terrenos Indios), Campo del Prado (Colonia el Mayor), Campo Flores, Campo Sonora (Colonia Terrenos Indios), Colonia la Puerta, Comunidad Indígena Cucapá el Mayor [Ejido el Mayor], Ejido Cucapá Mestizo, Ejido Doctor Alberto Mota (El Indiviso), Ejido Durango, Ejido México, Familia Regalado (Ejido Sonora 2 Campos Nuevos), La Casa de las Curvas (Colonia el Mayor), Mexicali, Sainz Domínguez (Colonia el Mayor), San Felipe. Sonora. San Luis Río Colorado: Pozas de Arvizu (La Reserva). |
| Mexicano alto de occidente (Yuto-nahua) | 127 | Nayarit | Acaponeta: Acaponeta, Agua Tendida, Buenavista (Las Paredes), El Duraznito, El Naranjo, El Resbalón, Huanacaxtle, La Guásima, La Laguna (La Lagunita), La Paloma Nueva Reforma, Mesa las Arpas, Paredes, Rancho los López, Rancho de Ernesto Aguilar, San Diego de Alcalá, San Diego el Naranjo, San Dieguito de Abajo, San José de Gracia, Santa Cruz, Saycota, Sayulilla, Unidad Habitacional, Zacatecas. Del Nayar [El Nayar]: Dolores, Los Chapiles, Santa Teresa, Tierras Cuevas |
| Mocho`(Maya) | 141* | Chiapas | Motozintla: Motozintla de Mendoza |
| Pima del norte (Yuto-nahua) | 155 | Chihuahua | Madera: Agua Amarilla, Ciudad Madera, Ejido el Lago, El Cable, El Campo Seis, El Cordón, El Cuatro, El Largo, El Pedregal, El Potrero, El Presón, El Represito, El Río Chiquito, El Táscate, El Yerbanís, Junta de los Arroyos [Junta de los Ríos], La Bolada, La Ciénega, La Nopalera, Las Espuelas, Las Lajas, Los Arbolitos, Madera, Mesa Blanca, Mesa el Tecolote, Mesa de La Simona, Mesa del Garabato, Rancho Huapoca, San Isidro, San Juan de Enmedio. Moris: Bermúdez, Los Cien Pinos |
| Pima del sur (Yuto-nahua) | 159 | Chihuahua | Ocampo: Basaseachi, Pinos Altos, Santo Domingo, Sapareachi, Tierritas. Temósachi: Agua Caliente, Casa Blanca, Ciénega Blanca, Cordón de Enmedio (Paraje Piedra Colorada), El Arbolito, El Llorón, El Nogal, El Saucito, El Terrero (Piedras Azules), Janos, La Ciénega, La Ciénega Blanca [Ciénega Blanca], La Guajolota, La Ornela, La Providencia, La Salitrera, La Vinata, Las Tierritas, Los Hornitos, Nabogame, Peñasco Blanco, Piedra Blanca, Piedras Azules, San Antonio, San Ignacio, Temósachi, Tierras de María, Yahuirachi, Yepáchic. |
| Paipai (Cochimí-yumana) | 162 (25) | Baja California (26) | Ensenada: Arroyo de León (Ejido Kiliwas), Camalu, Cañón de la Parra, Comunidad Indígena de Santa Catarina, Ejido 18 de Marzo (El Álamo), El Aguajito (El Mat Chip), El Alamar, El Pinacate [Chknan], El Ranchito [Wikwalpuk], El Sauzal, Ensenada, Ex Hacienda Sinaloa, Héroes de la Independencia (Llano Colorado), Lázaro Cárdenas (Valle de Trinidad), La Huerta, La Vinata [Wipuk], Leyes de Reforma (El Rodeo), Misión Santo Domingo, Ojo de Agua Colorada, Poblado Héroes de Chapultepec, Poblado Puerta Trampa, Pórticos del Mar, Ranchito Xonuko, Rancho Agua de Vida, Rancho el Sauco, Rancho Escondido, Rancho las Canoas, Rancho Mariscal, Rancho San Belem, Rancho Santa Martha, Real del Castillo Nuevo (Ojos Negros), Rincón de Santa Catarina, San Isidoro, Sauce Largo [Yokakgul], Sauce Solo [Yokazis], Úrsulo Galván, Valle de la Trinidad, Yokakiul |
| Zapoteco de la Sierra sur, noroeste bajo (Oto-mangue) | 164* | Oaxaca | Villa Sola de Vega: Agua del Sabino, Arroyo de la Rosa (El Aguacate), Barrio de Santa Anita (Sección Tercera), El Anís (San José El Anís), El Duraznillo, El Mangalito, El Nido, El Progreso, Guayabo, La Ciénega de la Palma, La Labor, La Nopalera, La Siempre Viva, La Singuera, Las Juntas (El Puente), Las Trancas, Llano Mazorca, Llano Monte, Llano de León, Loma de Ocote, Río Espejo, Río Húmedo, Río Manteca, Río de Boca, San Felipe Zapotitlán, San Juan Viejo, San Sebastián de las Grutas, Villa Sola de Vega |
| Chocholteco del este (Oto-mangue) | 180 | Oaxaca | San Juan Bautista Coixtlahuaca: El Capulín (Sección Primera), El Tepozón (Sección Segunda), El Zapotal (Sección Tercera), La Mulata, Santa Catarina Ocotlán. San Miguel Chicahua: Llano Seco. |
| Chocholteco del sur (Oto-mangue) | 200 | Oaxaca | San Miguel Huautla: Ocotlán. Santa María Nativitas: Barrio Nicolás, Barrio Santiago, El Mirador, El Porvenir, Loma del Tepejillo, Pie del Cordoncillo, Primera Sección (Santa Cruz), San José Monte Verde, San Pedro Buenavista, Santa María Nativitas. |
| Zapoteco de Valles, oeste (Oto-mangue) | 211 | Oaxaca | Magdalena Mixtepec: Magdalena Mixtepec, Santa Catalina Mixtepec. San Bernardo Mixtepec: Asunción Mixtepec, Barrio del Carmen, Barrio del Rosario, La Loma Grande, Rancho Teja, San Bernardo Mixtepec, San Jerónimo Tititlán. San Pablo Cuatro Venados: El Esponjado, El Jabalí, El Moral, El Poleo, El Sacatón, La Nevería, Llano Manteca, Llano de Toro, Piedra Blanca, Río Jalapillas, Río Minas, San Cristóbal, San Pablo Cuatro Venados, Santiago Clavellina. San Pablo Huixtepec: San Pablo Huixtepec. |
| Mixteco del noroeste bajo (Oto-mangue) | 220* | Oaxaca | San Bartolo Soyaltepec: Barrio del Calvario, Cerro Totocani, El Caballo Blanco, El Águila, Guadalupe Gavillera, Jataitno, La Unión Reforma, Río Verde, San Bartolo Soyaltepec, San Isidro Tejocotal, Santa Cruz Pípila. Santa María Chachoápam: San Agustín Monte Lobos, Santa María Chachoápam. Santo Domingo Tonaltepec: Cerro de Sol, Deque Inu (Cabeza de Cigarro), Dequetoto, Río Verde, San Francisco Río Blanco, Santo Domingo Tonaltepec, Vista Hermosa, Yuquini. Santo Domingo. Yanhuitlán: Santo Domingo Yanhuitlán, Tooxi, Yudayo. |
| Kumiai (Cochimí-yumana) | 221 (27) | Baja California (28) | Ensenada: El Porvenir (Guadalupe), Ensenada, Francisco Zarco (Guadalupe), La Huerta, La Misión, Rancho Dinuwa, Rancho Plazola, Real del Castillo Nuevo (Ojos Negros), San Antonio Necua (Cañada de los Encinos), San Salvador de Gálvez. Playas del Rosarito: Comunidad Indígena San José de la Zorra (San José de la Zorra) [San José la Zorra], La Zorra, Primo Tapia. Tecate: Colonia Luis Echeverría Álvarez (El Hongo), Ejido Guadalajara 2 (El Gato), Ejido Nueva Colonia Hindú, El Álamo, El Testerazo, El Venado, Encino Solo, Escuela Mescuich [Mescuich], Familia López Gaspar, Hacienda Santa Verónica, La Ciénega, Rancho Cañada Verde, Rancho Jacuin, Rancho las Auras, Rancho las Priscilas, Rancho Limón, Rancho Neji Sección C [Juntas de Neji], Rancho Puerto el Roble, Santa Elena, Tecate, Valle de las Palmas. |
| Chontal de Oaxaca bajo (Chontal de Oaxaca) | 223 | Oaxaca | Santa María Ecatepec: La Reforma, San Juan Acaltepec, Santo Domingo Chontecomatlán, Santo Tomás Teipan |
| Mixteco de San Miguel Piedras (Oto-mangue) | 243* | Oaxaca | San Miguel Piedras: Chidoco (Chidoco de Juárez), Colorado, El Fresno, El Potrerito, Guadalupe Victoria, Río Minas (Río Minas Piedras), San Miguel Piedras |
| Zapoteco de San Bartolo Yautepec (Oto-mangue) | 246 | Oaxaca | San Bartolo Yautepec: Joaquín López Mariano (Rancho), Puerto San Bartolo, San Bartolo Yautepec |
Localities are from Diario Oficial de Federación (2008), status and notes are mainly from INALI (2012), Hammarström et al. (2018) and Simons & Fennig (2018). Critically endangered (< 50 speakers survive) and endangered (< 250 speakers survive) languages as in Simons and Fennig (2018); those denoted by * are in very high risk of disappearance (< 100 speakers survive) and high risk of disappearance (< 1 000 but > 100 speakers survive) following INALI (2012).
1. Localities included are those with historic settlements.
2. Also known as edúes y coras.
3. Related to Chatino and Zapoteco languages.
4. Last speaker died in the 1960s.
5. Extinct ca. 1950.
6.Boas (1917).
7. Last speakers reportedly died around 1930, but the 1990 census lists 12 speakers.
8. Formerly spoken by a small group of people. The last speaker was born in 1895 (Holt 2001) and no remaining speakers are known after 1972.
9. As many as 100 in 1970; see also Lionnet (1978).
10. Ethnic population is 32.
11. Also known as heve and dohema; extinct since about 1940; see also Lionnet (1986).
12. In her essay “El noroeste: Sonora” (http://www.fundacionunam.org.mx/humanidades/eudeves-una-etnia-extinta/), historian Isabel Verdugo de Juárez locates the territory occupied by Eudeves as: “In the north they were found along the mid-portion of the San Miguel River (Horcasitas River) in Saracachi, Cucurpe o Opodepe. In the south they we found at the springs along the Matepe River, the banks of Moctezuma River and part of Nevoma or Yaqui Rivers. This area encompassed the historical sites of Batuco, Tepupa, Bacanora, Soyopa and Tónachi.”
13. Extinct in 1954 (Boas 1917; Knab 1980).
14. From the first contact with Europeans 300 years ago, the Cochimíes have occupied the central part of the Baja California peninsula and the state of Baja California Sur. Although there was some minor dialect variation over the vast area where Cochimí was spoken, they were mutually intelligible (T. Bowen, pers. comm.). Originally, there were no large settlements and they were basically nomads. The guamas or sorcerers had an important position in the original culture; there were no writing or formal artistic expressions. They were gatherers and fishers, who did not practice agriculture nor have livestock.
15. Ethnic population is 150; this may include Kumiai in La Huerta who call themselves Cochimí; old Cochimí is extinct (Simons & Fennig 2018).
16. A total of 15 Kiliwa people survived in 2018 and the number of speakers is given as 29 (INALI 2012); however, in February 2018 one of the last three speakers died in Ejido Tribu Kiliwas, Valle de la Trinidad, near Ensenada.
17. Ethnic population of 16 in 2016.
18. It has been reported that in 2003 there were only two native speakers (the youngest then 72 years old) and three semi-speakers; the community apparently agreed to stop speaking Zapotec in 1965 (Beam de Azcona, 2004).
19. A total of 166 elder speakers was also reported (INALI 2010).
20. Reported as Xuani-Ixcateca by Molina Cruz (2010), who described efforts to save this language using video (see also https://www.youtube.com/watch?v=lN2M-Rb7LM0).
21. A total of population of nine and only a few elder speakers (in 2009) were reported (Simons & Fennig, 2018).
22. A total of 30 speakers, all over the age of 70, were reported in 1998 (Simons & Fennig, 2018).
23. The Kickapoo arrived in Mexico in the nineteenth century when, after the Anglo-Saxon invasion of their territory, they asked the Mexican government for a place to live; in exchange the government asked them to defend Mexican residents against the frequent attacks from the Comanche. Since then, the Kickapoo of Mexico have been known in the United States as the “Texas gang.”
24.Caccavari Garza (2014) reported less than 30.
25.Caccavari Garza (2014) reported less than 50.
26. According to Caccavari Garza (2014) most people are in Comunidad Indígena de Santa Catarina, but they are also present in San Isidoro, Valle de la Trinidad and Ejido Héroes de la Independencia.
27.Caccavari Garza (2014) reported less than 50.
28. According to Caccavari Garza (2014) mainly located in two municipalities and the communities of Juntas de Nejí en Tecate, San José de la Zorra, San Antonio Necua y La Huerta, Ensenada.
TABLE 3 States in Mexico with extinct or endangered languages
| State | Number of extinct and endangered languages |
| Oaxaca | 16 |
| Baja California | 7 |
| Chiapas | 6 |
| Campeche | 6 |
| Sonora | 4 |
| Quintana Roo | 4 |
| Chihuahua | 3 |
| Jalisco | 2 |
| Tabasco | 1 |
| Veracruz | 1 |
| Guerrero | 1 |
| Tamaulipas | 1 |
| Baja California Sur | 1 |
| Mexico State | 1 |
| Coahuila | 1 |
| Nayarit | 1 |
As a stark reminder, one language and one species, both located in Baja California, will probably go extinct within the next very few years. With only two speakers alive in late 2018, the Kiliwa is Mexico’s most endangered language and is condemned to extinction (INALI, 2012; Caccavari Garza, 2014). And, with a population of fewer than two dozen individuals in 2018 (Comité International para la Recuperación de la Vaquita [CIRVA], 2019), probably even fewer today, the vaquita porpoise (Phocoena sinus), Mexico’s only endemic and the world’s most endangered marine mammal, is being driven to extinction by illegal fishing for the endangered and endemic totoaba (Totoaba macdonaldi) (Vidal, 1995; Brusca, Álvarez-Borrego, Hastings, & Findley, 2017; Thomas et al., 2017; Rojas-Bracho et al., 2018).
Discussion
Places with high species diversity, especially tropical forests, tend to have high linguistic diversity, and areas with low species diversity, such as tundra and deserts, have low linguistic diversity (Sutherland, 2003; Loh & Harmon, 2005). For instance, Papua New Guinea covers less than 1 % of the world’s land area but is home to the world’s third largest tropical rainforest (after the Amazon and the Congo) (Bryan, Shearman, Ash, & Kirkpatrick, 2010), and is also home to 6-8 % of the Earth’s animal and plant species (two thirds of them endemic) and 12 % of the world’s living languages (Papua New Guinea’s Fifth National Report to the Convention on Biological Diversity, 2014; Simons & Fennig, 2018).
Indigenous people are custodians and landowners of much of the biodiversity worldwide. The world’s 370 million indigenous people make up less than 5 % of the total human population, yet they manage or hold tenure over 25 % of the world’s land surface that supports about 80 % of the global biodiversity (Raygorodetsky, 2018).
There are increasing numbers of examples of non-industrial people living in harmony with their natural environment, such as rural communities in Hawaii; the Kayapo Indians of Middle Xingu Valley in Brazil (Posey, 1999); the Dai, an indigenous ethnic group in Southwest China (Shenghi, 1999); and the Sápara of Ecuador (Raygorodetsky, 2018). Numerous studies have proven how traditional ecological knowledge and practices have effectively served to protect and maintain natural environments (e.g., Posey, 1999; Berkes, 2001; Cunningham, 2001; Wiersum, 2004; Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services [IPBES], 2018). In addition, biocultural conservation can help secure the rights of indigenous and local people and help maintain a focus on social justice.
The current rate of extinction of languages worldwide, and loss of knowledge they contain, has no parallel in human history (Harrison, 2007). Language loss in some areas, such as the Americas, has reached 60 % over the last 35 years (Harmon & Loh, 2010). Indeed, the most rapid losses in linguistic diversity have occurred in this region, where 60 % of languages are threatened or have gone extinct since 1970. And this is not limited to developing countries. Policies in the United States, particularly between the 1870s and 1930s, greatly suppressed Native American languages and culture. It was only after years of activism by indigenous leaders that the U.S. Native American Languages Act was passed in 1990, which allowed for the preservation and protection of indigenous languages.
Languages can go extinct either because the entire population of speakers dies out, or because the speakers shift to a different language and forget their mother tongue due to the loss of intergenerational transmission. Much of the decline in linguistic diversity is a result of language shift away from small indigenous languages toward national or regional languages (Loh & Harmon, 2014). Along with the disappearance of languages, most of the rich traditional knowledge of these indigenous cultures is largely lost forever, including knowledge of the natural history of the world in which the language resided.
Languages are a critical measure of the cultural diversity of nations, while species richness is a measure of their biological diversity. Mexico offers an important case on the challenges to conserve both. With at least 118 species of economically important plants totally or partially domesticated by indigenous pre-Hispanic farmers, Mexico has been a global center of plant domestication. More than 15 % of edible vegetable species consumed in the world originated in Mexico (CONABIO, 2008; Perales & Aguirre, 2008; Sarukhán et al., 2017). In addition, between 3 000-4 000 medicinal plant species are regularly used by Mexicans, and indigenous people in Mexico use 5 000-7 000 plant species in various cultural activities (Boege, 2008). It has been suggested that almost every species of plant and animal, type of soil, landscape, and mountain in Mexico has its match in a linguistic expression, knowledge category, practical use, mythical or religious meaning, or a known individual or collective experience (Toledo et al., 2001a, Toledo et al., 2001b).
Mexico’s early cultures relied heavily on natural resources, including the harvesting of coastal and estuarine resources in the Holocene. In the Baja California-Gulf of California region this included fishing and intensive hunting of dolphins and sea turtles beginning at least 10 000 years ago (Bowen, 1976, Bowen, 2000, Bowen, 2006, Bowen, 2009; Felger & Moser, 1987; Felger, Nichols, & Seminoff, 2005; Fujita, 2006; Marlett, 2014). By around the year AD 1000, four important centers of indigenous socio-economic and cultural concentration surged in this region, which reflected changing patterns of harvesting of marine resources. First, during the early 16th and 17th centuries, the Pericú, who lived along the deserts of Los Cabos region in the South of the peninsula, were hunters and shell gatherers (concheros), and disappeared (together with their language, its phylogenetic classification remaining unknown) in the second half of 18th century (Fujita, 2006). The exploitation of marine mammals, the associated stone tools, and the construction of enormous shell mounds by the Pericú display striking similarities to the Chumash people of the Channel Islands in Southern California in the United States (Beer, Gonzalez, Huddart, Rosales-Lopez, & Lamb, 2008). Second, the Guaicuras and the Cochimís, the latter inhabiting the center and North of the Baja California peninsula, were also shell gatherers and fishers. Third, the Seris on the (“mainland”) coastal Sonoran Desert were (and still are) fishers and gatherers. And fourth, the Yumas, Pimas Altos and Papagos in North-Central Sonora and the tip of the peninsula were desert gatherers-farmers (Nolasco, 1982). The central desert area of the Baja California peninsula had the least linguistic diversity (and probably the lowest biological diversity; T. Bowen, personal communication).
Loss of biocultural diversity in Mexico (and the Americas in general) began as soon as Europeans arrived. Although estimates of the Native American population size of Mexico upon first arrival of Europeans in 1519 vary greatly (from less than 3 million to over 52 million), most researchers put it at around 20 million (Koch, Brierley, Maslin, & Lewis, 2019). At the time, large areas of land were under cultivation with maize, cacao, and fruit orchards (Whitmore & Turner, 1992). This indigenous population quickly began to collapse from warfare, slavery, and the introduction of pathogens unknown in the Americas (e.g., influenza, smallpox, bubonic plague). The most devastating epidemic in Mexico occurred in 1520, when a single smallpox outbreak killed an estimated 30 to 50 % of the indigenous population (Cook & Borah, 1960; Dobyns, 1993; McCaa, 1995). Following the first comprehensive census in 1568, the population of central Mexico had already declined to 2.7 million (Sanders, Pasons, & Santley, 1979), which corresponds to an approximate decline of 87 % within the first 50 years of European arrival (based on a contact population of 20 million) (Whitmore & Turner, 1992). A rapid population collapse of up to 90 % is plausible for the most populous parts of the country (Koch et al., 2019). One can only speculate how many indigenous languages were lost or put on a path toward extinction during this period. In Northern Mexico many groups of hunters and gatherers who spoke unknown languages perished because of the extermination campaigns launched by Spanish conquerors (Borah & Cook, 1963).
Today, there is strong geographical overlap between the states that harbor the bulk of Mexico’s biological and linguistic diversity, with Oaxaca, Puebla, Chiapas, Veracruz, Guerrero and Michoacán standing out. Furthermore, Mexico’s biodiversity hotspots closely mirror its language hotspots: areas with the highest number of endangered plant and animal species overlap with those where the endangerment of languages is greatest. Most of Mexico’s dry forests, tropical rain forests, and temperate rain forests, which are home to high levels of biological and linguistic diversity, belong to indigenous communities, and almost a third of the country’s federally protected areas are within indigenous territories (Sarukhán et al., 2017). It has been estimated that nearly 90 % of Mexico’s indigenous population live in forested areas, while the rest live in arid and semi-arid regions with shrub or grasslands (Toledo et al., 2001a, Toledo et al., 2001b).
Oaxaca is a good example of how the geography of languages and biodiversity overlap and have evolved together. Its complex geological history produced an elaborate topography and highly diverse range of ecosystems, from tropical coastal areas to temperate pine-oak forests, thorn scrub and cloud forests, which in turn favored adaptive radiation, speciation, and a high diversity of flora and fauna (Ordoñez, 2004). At least 8 431 species of plants (nearly 40 % of Mexico’s known flora) and 4 542 animal species (half of the country’s vertebrates and 19 % of its known invertebrates) are present in Oaxaca (García-Mendoza, Ordoñez, & Briones-Salas, 2004; CONABIO, 2008). For more than 10 000 years, its indigenous peoples have dispersed and evolved within the state’s diverse ecosystems, and today 157 languages (43 % of Mexico’s 364 languages) are spoken in more than 4 000 indigenous communities (de Ávila Blomberg, 2004).
The most insidious threats to Mexico’s biodiversity are habitat destruction and fragmentation (mostly by deforestation for agriculture and livestock), overexploitation, invasive species, and climate change. The country has already lost about 70 % of its forest cover, mainly the tropical forests of the Southeast (Sarukhán et al., 2017). Prieto-Amparán et al. (2019) analyzed non-tropical land use in Mexico and found the area of primary forest reduced from 55.8 % in 1990 to 37.7 % in 2017 in their study region (which included temperate primary and secondary forest, human settlements, unvegetated areas, and water bodies). In just one year (2016), 253 000 hectares of Mexico’s forests disappeared (Global Forest Watch, 2018). The effects of climate change will exacerbate all those threats (Peterson, Tian, Martínez-Meyer, Soberón, & Sánchez-Cordero, 2005).
The largest areas of forest and wildlands in Mexico are usually communal lands (Stoleson et al., 2005), and at least 70 % of its forested areas are held by ejidos (Segura, 2000; Molnar & White, 2001). However, this is rapidly changing as ejidos sell their land to private landowners (Stoleson et al., 2005), typically to non-campesinos, and the cultural erosion and loss of traditional knowledge that ensues encourages loss of biodiversity (Laird, 2002).
Languages disappear for a variety of reasons: all speakers may die; speakers may succumb to pressures to speak a different language and forget their mother tongue; speakers may choose to stop speaking their native tongue because of interests in participating in national education, finding jobs that pay salaries and could come with various benefits, or marry into a family with speakers from another language. The threat to Mexico’s languages includes a combination of factors such as the reduced number of speakers, their geographical dispersion, predominance of adult speakers, and trends for abandoning transmission of languages to new generations (INALI, 2012). All of this is compounded by a lack of interest or even neglect by authorities, which has led to exclusion of indigenous languages from public and institutional spaces, mass media communications such as radio and television, and their diminishing use among communities and families. Some indigenous people seem to have accepted that their languages will disappear, and they will soon be able to communicate only in Spanish (INALI, 2012), while others desire to reverse the extinction trend. And many more simply do not know what to do to save their (Mexico’s) cultural heritage.
Traditional peoples have accumulated vast amounts of ecological knowledge, and that knowledge is embodied in their languages. Thus, as languages go extinct, associated traditional ecological knowledge is also lost (Oviedo et al., 2000). This happens because, in most traditional cultures, knowledge is not recorded in writing but is passed on orally to other groups or new generations. In such cases, the loss of local languages means the loss of the traditional means of knowledge transmission.
The science of biocultural diversity is in its infancy. Although we do not yet fully understand it, there is evidence of ancient and profound connections between biological and linguistic diversity. When species go extinct entire biological communities and ecosystems can be disrupted or permanently altered. Regardless of their economic, scientific and aesthetic value to humans, plant and animal species have a value of their own because of their unique evolutionary history, genomic diversity, and, ultimately, because of their very existence. When we allow languages to disappear, we squander the culture of humanity, the millennial knowledge of the natural world, and a part of our own past. The IPBES released its definitive new global synthesis of “Assessing nature’s contributions to people” (Díaz et al., 2018). It is the first such report since the landmark MEA (2005) was published, and the first ever that is fully intergovernmental. It is also the first assessment to systematically include indigenous and local knowledge at a global scale.
For decades, scientists and conservationists have advocated for the establishment of protected areas to conserve ecosystems and biological diversity (Vidal, López-García, & Rendón-Salinas, 2014). But, as has been advocated for more than two decades, the world’s biodiversity will only be effectively preserved by also protecting the diversity of human cultures, and vice versa (Oviedo et al., 2000; Maffi, 2001; Toledo, 2001b). Protecting areas that are implemented under participatory and co-management schemes is the most effective strategy for conserving biodiversity while improving the economic and social conditions of local communities. Conservation area networks comprising priority areas should serve as the baseline for identifying the most suitable strategies depending on the areas’ socio-economic contexts and specific characteristics.
We have shown that Mexico’s biocultural diversity is at crossroads. Federal, state and municipal governments, businesses, conservation organizations, philanthropists, and multilateral agencies that care about biodiversity need to realize that their resources and efforts will only be effective in the long-term if they simultaneously support protection of indigenous cultures and traditional knowledge. Given that the regions harboring Mexico’s highest biocultural diversity are considered strategic for the country’s water, environmental and food security, as well as to ensuring the rights of indigenous peoples, it has recently been proposed that they should be an important component of the 2019-2024 national development plan (Luque & Ortiz Espejel, 2019). We hope this policy initiative results in concrete action to protect Mexico’s unique cultural and natural heritage. Biological and cultural diversity are two sides of the same coin. Since endangered languages and endangered species strongly overlap geographically in Mexico, it makes sense to combine efforts to protect both. But time is of the essence. All constituents need to urgently reinforce efforts and significantly augment investments if we are to rescue and preserve the country’s unique biocultural diversity and traditional knowledge for the benefit of present and future generations.
Ethical statement: authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.
Supplemental Material
