











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Existen diferentes humedales definidos por la interacción del agua, el suelo hídrico y la vegetación hidrófita, que junto con la geomorfología y el clima resultan en distinta composición y estructura de la vegetación (Wheeler, Money, Shaw, Perrow, & Davy, 2002; Mitsch & Gosselink, 2007). El interés por entender su funcionamiento ha aumentado debido al reconocimiento de los servicios ecosistémicos que proveen y su papel clave en la mitigación y adaptación al cambio global (Costanza et al., 1997; MEA, 2005; de Groot et al., 2012). Sin embargo, el cambio climático y el cambio de uso de suelo están modificando sus características bióticas y abióticas, alterando procesos funcionales como el almacenamiento de agua, la productividad y el almacenamiento de carbono (Bergh et al., 2001).
La vegetación, es un componente de los humedales, y los patrones de la distribución de las especies que la componen están relacionadas con los factores ambientales como el tipo de suelo, la disponibilidad de nutrientes, el hidroperiodo, la topografía que han sido estudiados por diversos autores (Zeilhofer & Schessl, 1999; Elmore, Mustard, & Manning, 2003; Crain, Silliman, Bertness, & Bertness 2004; King, Richardson, Urban, & Romanowicz, 2004; Givnish et al., 2008; Crase, Liedloff, Vesk, Burgman, & Wintle, 2013). En los humedales costeros, las características hidrogeomorfológicas definen las interacciones entre el ambiente terrestre y oceánico (de la Lanza Espino, Pérez, &Pérez, 2013) que condicionan las variaciones de los factores ambientales como la salinidad y la inundación, considerados los principales factores que determinan la composición, estructura y diversidad de la vegetación (Brinson, Bradshaw, & Jones, 1985; Carreño, Esteve, Martínez, Palazón, & Pardo, 2008; Anderson & Lockaby, 2011; Courtwright, & Findlay, 2011).
El hidroperiodo del humedal es el patrón estacional del nivel del agua, resultado del balance entre las entradas y salidas de agua, la geomorfología y el suelo, donde los procesos biogeoquímicos, como la reducción del hierro, magnesio y bióxido de carbono, se rigen por las reacciones de óxido-reducción (redox) influenciados por el tiempo de inundación (Mitsch & Gosselink, 2007; Vepraskas & Faulkner, 2001). La salinidad en los humedales costeros está determinada por el hidroperiodo, las fluctuaciones de la marea, la salinidad del agua del mar, los cambios en el nivel del mar, los aportes de agua dulce de los ríos, la precipitación y el agua subterránea (Zedler & Callaway, 2001; Barendregt & Swarth, 2013).
La relación entre los humedales costeros se ha evaluado con base a la conectividad hidrológica (Zedler & Callaway, 2001; Davis, Childers, Lorenz, Wanless, & Hopkins, 2005). Poco se sabe sobre la conectividad estructural (vegetación), la mayoría de los estudios están referidos a un tipo de humedal (e.g. manglar, selva inundable) (Agraz-Hernández, García-Zaragoza, Iriarte-Vivar, Flores-Verdugo, & Moreno-Casasola, 2011; Infante-Mata, Moreno-Casasola, Madero-Vega, Castillo-Campos, & Warner, 2011; Moreno-Casasola et al., 2010) y reportan la diversidad alfa, con excepción de algunos trabajos que estiman la diversidad beta para conocer el recambio de las especies y la conectividad que puede existir entre estos humedales heterogéneos donde existe un gradiente ambiental que se refleja en un cambio en la estructura y composición de la vegetación (Rickert, Fichtner, van Klink, & Bakker, 2012; Socolar, Gilroy, Kunin, & Edwards, 2016; Aponte, 2017). En general la diversidad vegetal de los humedales costeros es baja y dominada por algunas especies, debido a que son pocas las plantas con adaptaciones morfológicas y fisiológicas, para crecer en zonas inundadas y bajo influencia salina (Hackney & Avery, 2015; Lee, Jones, Diele, Castellanos-Galindo, & Nordhaus, 2017).
En México, Chiapas ha perdido el 57 % de humedales, principalmente los costeros de agua dulce (Landgrave & Moreno-Casasola, 2012; Valderrama-Landeros et al., 2017). Las actividades antrópicas a lo largo de las cuencas costeras afectan a la planicie, causando el deterioro del hábitat, la modificación de los procesos funcionales (e.g. productividad, almacenamiento de carbono) y disminuyendo la conectividad entre los humedales costeros de Chiapas (Contreras-Espinosa & Warner, 2004; Calva, Pérez, & Márquez, 2006; de la Fuente, Lisle, & McKean, 2007; Alonso-Eguialis et al., 2013).
La planicie costera de Chiapas es angosta, con un ancho entre 15 y 35 km, con un relieve bajo que se eleva abruptamente a casi 2 000 msnm. La precipitación anual es de 1 500 a 2 000 mm a lo largo de la planicie y de 2 100 a más de 4 000 mm en la parte alta. Las características hidrogeomorfológicas de los ríos (pendiente, topografía, caudal, sinuosidad, entre otros), determinan la distribución, la dimensión y la morfología de los humedales costeros (Lisle, De la Fuente, Suarez, & Hernández, 2009; de la Fuente et al., 2007; Pérez, 2013). Los humedales que se observan del mar hacia tierra adentro son: (1) el manglar, que cubre una gran extensión y se conecta horizontalmente a través de los canales y los esteros con los principales cuerpos lagunares (Valderrama-Landeros et al., 2017); (2) la selva inundable, que ocupa una extensión mucho menor que el manglar, y forma parches remanentes de pocas hectáreas en el Sistema de Humedales el Castaño (SHC) y la desembocadura del río Huixtla (Breedlove, 1981; Rincón, 2014); (3) los humedales herbáceos, tulares y popales, que ocupan también grandes extensiones y mantienen conectividad horizontal a través de canales (Rzedowski, 1978; SEMARNAP, 1999); y (4) humedales modificados como los pastizales inundables, que son áreas ocupadas para uso ganadero (Liscovsky, Parra, Balente, Huerta, & Ramos, 2009).
En este trabajo se aborda el estudio del SHC para entender el funcionamiento (e.i. la forma en que interactúan los factores bióticos y abióticos, su relación con la distribución de la vegetación y el mantenimiento de los procesos ecológicos) de este tipo de sistema de humedales costeros. El objetivo fue caracterizar las variables del hidroperiodo, las características fisicoquímicas del agua y el suelo para determinar su relación con la distribución de la vegetación de los humedales. Se plantean las siguientes hipótesis: i) Se espera encontrar un gradiente ambiental definido por la inundación y salinidad de mayor a menor influencia marina al aumentar los aportes de agua dulce del continente, que se reflejará en un cambio en la estructura y la composición de la vegetación; ii) la diversidad alfa variará de menor a mayor al disminuir la inundación y la salinidad; la diversidad beta (recambio de especies) será mayor entre las comunidades vegetales con mayor diferencia de inundación y salinidad.
Materiales y métodos
Área de estudio: El SHC forma parte de la región hidrológica prioritaria para la conservación 32, de acuerdo con la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO) (SEMARNAP, 1999; Tovilla et al., 2007), se localiza en la planicie costera del Pacífico Sur, en donde desembocan los ríos San Nicolás, Sesecapa, Madre Vieja y Bonanza-Camargo y forma parte del humedal El Castaño-Barra Zacapulco-Laguna Campón y Chantuto, al NW de la zona núcleo de la Reserva de La Biósfera La Encrucijada (REBIEN) (15°16’51.93” N & 92°57’55.02” W) entre las coordenadas 15°17’7.72” N & 92°56’57.76” W y 15°15’40.56” N & 92°57’31.47” W (Fig. 1). El clima es cálido-húmedo, con abundantes lluvias en verano (Am) (García, 2004), la precipitación anual es de 1 567 mm y temperatura media anual de 28.2 °C con un mínimo de 19.2 °C y un promedio anual máximo de 36.5 °C (CONAGUA-SMN, 2018). La vegetación es característica de la REBIEN y está formada por selva baja caducifolia, vegetación de dunas costeras, palmares, popales, tulares, manglares, selvas inundables, vegetación flotante y subacuáticas (Rzedowski, 1978; Breedlove, 1981), así como potreros y cultivos diversos.
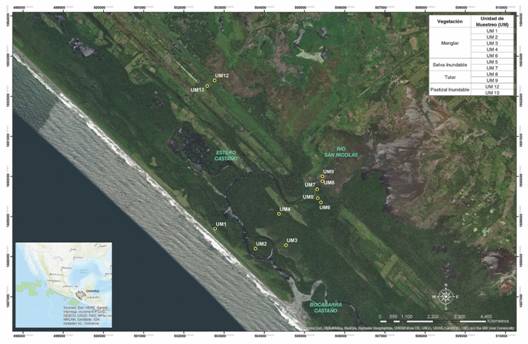
Fig. 1 Fig. 1 Área de estudio, Sistema de Humedales El Castaño, Chiapas, México. Study site, El Castaño Wetlands System, Chiapas, Mexico.
Método: Con base al conocimiento previo de la zona y con apoyo del visualizador Google Earth se propusieron los sitios de muestreo. Se realizaron dos recorridos de verificación para ubicar los tres tipos de humedales naturales (manglar, selva inundable y tular), y un humedal modificado por el uso ganadero (pastizal inundable). Se establecieron en total 11 unidades de muestreo (UM) permanentes por humedal: cinco en el manglar, dos en la selva inundable, dos en el tular y dos en el pastizal inundable. En el manglar se establecieron cinco UM porque la superficie que cubre es mayor (Carranza & Molina, 2003) y presenta una mayor heterogeneidad (Romero-Berny, Tovilla-Hernández, Torrescano-Valle, & Schmook, 2019). El muestreo es intencional y dirigido como lo sugiere Fourqurean et al. (2014). En todas las UM se caracterizó la vegetación (al inicio del trabajo) y en cada parcela se muestreó mensualmente de mayo 2016 a octubre 2017, el tiempo (meses) y el nivel de inundación; y de noviembre 2016 a octubre 2017, los parámetros fisicoquímicos del agua (superficial, intersticial y subterránea) y del suelo.
Hidroperiodo (nivel y tiempo de inundación): Se midió el nivel del agua en 52 piezómetros, construidos e instalados considerando el método de Yetter (2004) y Peralta-Pélaez, Infante-Mata y Moreno-Casasola (2009). Para conocer el hidroperiodo se graficaron por piezómetro los valores mensuales con respecto al nivel del suelo. El nivel de inundación de cada piezómetro es independiente de otro, ya que no fueron calibrados con respecto a un punto de referencia común, como el nivel del mar.
Parámetros fisicoquímicos: Se midió la salinidad y el pH del agua con un multiparámetro YSI Mod. 85 y un sensor de pH eco Testr modelo pH2 (Oakton ™). Los datos de agua superficial se tomaron directamente del espejo de agua, el agua intersticial entre 15-20 cm del suelo y el agua subterránea a 150 cm del suelo, siguiendo los métodos sugeridos por Infante-Mata, Peralta-Pélaez y Arrocha (2009).
Se recolectaron muestras de los primeros 8 cm de suelo con un cilindro de cobre (190.07 cm3). Siguiendo el método de Wilke (2005) se obtuvo la densidad aparente y el porcentaje de humedad. El potencial redox (Eh) se midió utilizando tres electrodos de platino construidos con el método de López-Rosas y Tolome (2009), que incluye la calibraron y ajuste a los cálculos.
Estructura y composición de la vegetación: Para la caracterización de la vegetación se utilizaron los métodos sugeridos por Moreno-Casasola y Warner (2009) y Kauffman, Donato y Adame (2013). En el manglar y la selva inundable se establecieron cinco parcelas circulares de 12.6 m de radio sumando 2 500 m2 por UM. En el tular y el pastizal inundable fueron cinco parcelas rectangulares de 25 x 20 m cada una, sumando 2 500 m2 por UM; esta parcela se dividió en subparcelas de 5 x 5 m y se muestrearon 10 subparcelas. En cada UM se identificaron las especies en campo y en caso de no definirla se identificó posteriormente con apoyo de guías y ejemplares de herbario de ECOSUR-Tapachula.
En la vegetación arbórea se registró el diámetro a la altura del pecho (DAP) de los individuos mayores de 2.5 cm. Para los árboles multicaules cada tallo fue considerado como un individuo. Para cada especie de la vegetación arbórea y herbácea, se estimó la cobertura usando la escala de cobertura-abundancia de Westhoff y van der Maarel (1978).
Análisis de la diversidad, estructura y dominancia: Se obtuvo la curva de acumulación de especies y dos pruebas no paramétricas de Chao2 y Jack-knife 1 para tener dos referencias al evaluar la representatividad del esfuerzo de muestreo de la riqueza. Para la diversidad alfa se obtuvo el índice de Shannon-Wiener (H’) y el índice de Simpson (D-1), puesto que son los más utilizados para comparar la diversidad entre estudios, además del índice de equidad de Pielou (J’). La diversidad beta, se evaluó con el índice de Sorensen. Se elaboró un histograma de frecuencia con las clases diamétricas de la vegetación arbórea de manglar y selva inundable. Se calculó el Índice de Valor de Importancia (IVI) considerando la suma de la frecuencia relativa, densidad relativa y dominancia relativa para la vegetación arbórea y en el caso de la vegetación herbácea la frecuencia relativa y la cobertura relativa.
Análisis de datos ambientales: Para verificar la normalidad de los datos se utilizó la prueba de Kolmogorov-Smirnov. Para las comparaciones de las variables entre los cuatro humedales y por temporada se utilizó la prueba de Kruskal-Wallis (H), ya que los datos no presentaron una distribución normal. Para las variables que presentaron diferencias significativas (P < 0.05) entre los humedales se realizaron pruebas de comparaciones múltiples de Tukey no paramétrica. Se realizaron las mismas comparaciones para la temporada de secas (noviembre 2016 a abril 2017) y lluvias (mayo a octubre de 2017). Para identificar diferencias entre el hidroperiodo y la precipitación se utilizó la prueba de correlación de Spearman.
Para establecer la forma en que se ordena espacialmente la vegetación de los sitios de muestreo e identificar las variables ambientales, que explicarán el gradiente ambiental, se hizo un análisis de escalamiento multidimensional no paramétrico (NMDS), que se considera el más adecuado ya que no asume linealidad, y se uso la distancia de Bray-Curtis, queda a las especies comunes y raras pesos relativamente similares. La matriz que se utilizó para esta prueba fue la de cobertura-abundancia por especie y parcelas. Para la mayoría de los análisis se utilizó el programa PAST v 3.2 (Hammer, Harper, & Ryan, 2001), las comparaciones múltiples y los análisis de Spearman se realizaron con el programa R (R Core Team, 2019).
Resultados
Hidroperiodo (nivel y tiempo de inundación): Las fluctuaciones de nivel de agua observadas en los humedales, siguen el patrón de la precipitación de la región (Fig. 2A, Fig. 2B). Los niveles aumentan en junio, al inicio de las lluvias, y bajan al disminuir la precipitación en noviembre. Cada humedal presenta una variación diferente en nivel y tiempo de inundación. En el nivel de inundación hay diferencias significativas (H = 62.16, P < 0.05, Tukey, P < 0.05), el pastizal inundable presenta los niveles más bajos en secas, pero en lluvias alcanza un nivel similar al del manglar y tular, siendo la selva inundable la de menor inundación (H = 36.15, P < 0.05). El manglar y el pastizal permanecieron inundados el mismo tiempo. En el manglar en las UM1, UM3 y UM4 se observaron parcelas con inundación permanente. En relación al tiempo de inundación el manglar, el tular y el pastizal inundable permanecen mayor tiempo inundados con respecto a la selva inundable (kw = 24.39, P < 0.05, Tukey, P < 0.05) (Apéndice Digital 1, Tabla A, Tabla B, Tabla C).
En el tular y el pastizal inundable, el aumento del nivel del agua se relaciona con la precipitación, (Spearman, P > 0.05). El manglar y la selva inundable presentaron un patrón diferente con la precipitación (Spearman, P < 0.05). En el manglar de mayo a octubre (lluvias), la inundación es generalizada y en noviembre (secas), disminuye, llegando al nivel más bajos en el mes de abril (Fig. 2C). En la selva inundable, aún cuando el nivel del agua subterránea aumenta en lluvias, no hay agua superficial en la mitad de las parcelas, y a partir de noviembre el agua llega a -90 cm por debajo del suelo (Fig. 2D). En el tular las fluctuaciones en la temporada de lluvia son similares en todos los sitios; el agua superficial se mantuvo hasta diciembre, y en los siguientes meses baja a -95 cm del suelo (Fig. 2E). En el pastizal inundable, el agua superficial sube hasta 120 cm y se mantiene en algunas parcelas hasta febrero; en los siguientes meses el nivel del agua desciende a -120 cm por debajo del suelo (Fig. 2F).
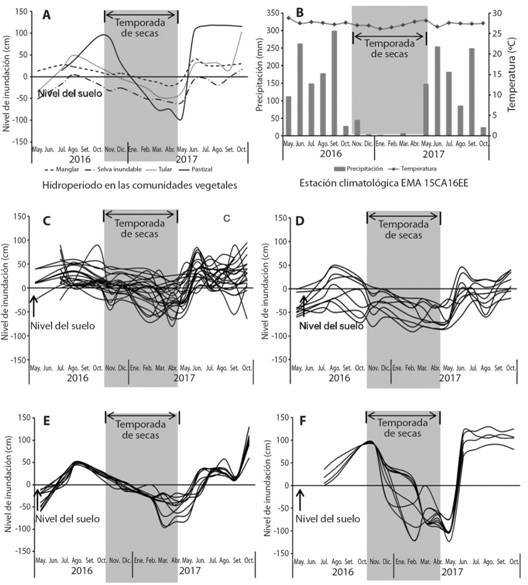
Fig. 2 Fig. 2 Hidroperiodos. A. Hidroperiodo general de humedales; B. Precipitación y temperatura C. Manglar; D. Selva inundable, E. Tular y F. Pastizal inundable, en el Sistema de Humedales el Castaño. Cada línea de las gráficas corresponde a un piezómetro. Hydroperiods. A. General wetland hydroperiod; B. Precipitation and temperature C. Mangrove; D. Freshwater swamp, E. Freshwater marsh and F. Flooded grassland, in El Castaño Wetland System. Each line of the graphs corresponds to a piezometer.
Parámetros fisicoquímicos del agua: Los humedales del SHC difieren en los parámetros fisicoquímicos del agua. La salinidad del agua superficial del manglar, la selva inundable y el tular son mayores a la del pastizal inundable (H = 52.42, P < 0.05, Tukey, P < 0.05). En el agua intersticial y subterránea, la salinidad, del manglar es mayor a las otras comunidades. Entre la selva inundable y el tular la salinidad es igual y la del pastizal es menor que todas, durante todo el año. La salinidad del agua intersticial del tular durante la temporada de lluvia es diferente de la selva inundable, y el pastizal inundable. Con respecto al pH, en el agua superficial del manglar y en el pastizal inundable, es diferente a la de la selva inundable y la del tular. No hay diferencias del pH del agual intersticial y subterránea, entre la del manglar y la de la selva inundable. El pH del tular es diferente al manglar en los tres tipos de agua. Durante la temporada de lluvias el pH del agua subterránea solo es diferente en el pastizal inundable (Apéndice Digital 1, Tabla A, Tabla B, Tabla C).
Parámetros fisicoquímicos del suelo: De las variables del suelo, el porcentaje de humedad de cada humedal fue diferente (H = 54.20, P < 0.05, Tukey, P < 0.05); los valores más altos se registraron en el manglar (90.76 %) y los más bajos en el pastizal inundable (7.7 %). En la temporada de secas la humedad del suelo de la selva inundable y el pastizal inundable, fueron similares (H = 26.83, P < 0.05, Tukey, P < 0.05). Con respecto a la densidad aparente del suelo solo el pastizal inundable y la selva fueron diferentes (H = 16.24, P < 0.05, Tukey, P < 0.05). El potencial redox, considerando el promedio anual, para el manglar fue diferente a los otros humedales (H = 40.84, P < 0.05, Tukey, P < 0.05). En secas el potencial redox del manglar es igual al de la selva inundable pero difieren con respecto a los humedales herbáceos (H= 25.22, P < 0.05, Tukey, P < 0.05). En temporada de lluvias el potencial redox de la selva inundable es mayor con respecto al de los otros humedales (H= 21.15, P < 0.05, Tukey, P < 0.05) (Apéndice Digital 1, Tabla A, Tabla B, Tabla C).
Composición y diversidad de la vegetación: Se registraron 78 especies pertenecientes a 33 familias. El pastizal inundable tiene la mayor riqueza (50), seguido por la selva inundable (18), el tular (16) y el manglar (14). De las 78 especies registradas, 53 son hierbas, 12 son árboles, seis pueden ser encontradas en forma de árboles o arbustos, hay una trepadora, dos pueden crecer como arbusto y trepador, tres como hierba y trepador y una epífita. Según la CONABIO (2016), 45 especies tienen hábitats terrestres, 16 dulceacuícolas, 11 en dulceacuícolas y terrestres, tres en dulceacuícolas y salobres, y tres en dulceacuícolas, salobres y terrestres (Apéndice Digital 2).
A nivel nacional, considerando la base de datos de la CONABIO (2016), se registraron dos especies endémicas (Hampea trilobata y Sabal mexicana), y seis especies exóticas Cucumis anguria, Eichhornia crassipes, Elaeis guineensis, Brachiaria decumbes, Cynodon dactylon y Panicum maximum, siendolas últimas tres pastos.
Las especies del manglar (R. mangle, L. racemos, A. germinans y C. erectus), tienen categoría de amenazadas (A) por la NOM059 en México. También se identificaron 21 especies evaluadas por la Unión Internacional para la Conservación de la Naturaleza (UICN) en categoría de bajo riesgo, preocupación menor (LR(lc)) (Apéndice Digital 2) (UICN, 2017).
La curva de acumulación de especies no llegó a la asíntota (Apéndice Digital 3), pero se obtuvieron los porcentajes de representatividad de las especies esperadas con los estimadores de Chao2 y Jack-knife1 (Tabla 1). Con el primer estimador la representatividad fue de 93 % para manglar, 95 % la selva inundable, 84 % el tular y 75 % el pastizal inundable. Con el segundo estimador, los porcentajes observados fueron, 78 % en el manglar, 82 % la selva inundable, 80 % el tular y 75 % el pastizal. Con los resultados de ambos estimadores se confirmó la observación mínimo del 75 % de las especies esperadas.
TABLA 1 TABLE1 Área muestreada por sitio, especies observadas y estimadas según los índices Chao2 y Jack-knife1 Area sampled by site, species observed and estimated according to the Chao2 and Jack-knife1 indexes
| Vegetación | Área (m2) | Especies observadas | Estimada Chao2 | Estimada Jack-knife1 |
| Manglar | 12 000 | 14 | 15 | 18 |
| Selva Inundable | 5 000 | 18 | 19 | 22 |
| Tular | 5 000 | 16 | 19 | 20 |
| Pastizal Inundable | 5 000 | 50 | 67 | 67 |
Para la diversidad alfa el mayor valor se obtuvo en el pastizal inundable (H: 3.44, D-1: 0.96, J´0.88), seguido por la selva inundable (H: 2.18, D-1:0.85, J´0.75), el tular (H: 1.92, D-1: 0.76, J´0.69) y el menor el manglar, (H: 1.66, D-1:0.74, J´0.63). El índice de Sorensen, indica un menor recambio de especies entre los humedales arbóreos (el manglar y la selva inundable), al presentar una mayor similaridad de 0.56 compartiendo nueve especies. Entre los humedales herbáceos (el tular y el pastizal inundable) la similaridad fue de 0.21 el recambio es mayor, se comparten siete especies. Entre los humedales arbóreos y herbáceos el cambio de especies es mayor, entre el manglar y el pastizal inundable la similaridad es de 0.06, solo comparten dos especies. Hay tres especies que se comparten en tres humedales: Zygia conzattii y Cissus sicyoides que se encuentra en el manglar, selva inundable y pastizal inundable, y Acrostichum aureum, que se encuentran en el manglar, selva inundable y tular.En general cada tipo de humedal tiene especies vegetales características que los definen, y no hay especies que se encuentren en los cuatro humedales (Tabla 2, Apéndide Digital 1).
TABLA 2 TABLE 2 Índice de diversidad beta de Sorensen Sorensen beta diversity index
| Selva Inundable | Tular | Pastizal Inundable | |
| Manglar | 0.56 | 0.07 | 0.06 |
| Selva Inundable | 0.12 | 0.09 | |
| Tular | 0.21 |
Estructura de la vegetación: En el manglar, se registraron 1 945 ind ha-1 y 20.50m2 ha-1 de área basal. Las especies con el mayor IVI fueron L. racemosa y R. mangle con 117.91 % y 116.14 %, respectivamente. En la selva inundable se estimó una densidad de 3 256 ind ha-1 y un área basal de 23.64m2 ha-1. La especie con el mayor IVI es P. aquatica (Apéndice Digital 4, Tabla A). La distribución de las clases diamétricas de individuos del manglar y selva inundable siguió una tendencia similar, disminuyendo hacia las clases de mayor diámetro. En ambos casos son más frecuentes los individuos con diámetros entre 2.5 y 7.49 cm (Fig. 3).
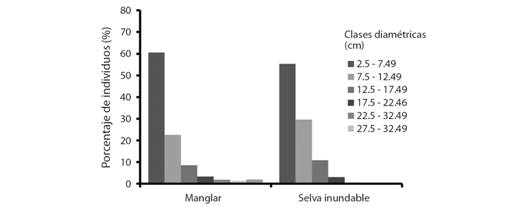
Fig. 3 Fig. 3 Porcentaje de individuos en cada clase diamétrica del manglar y de la selva inundable del Sistema de Humedales el Castaño. Percentage of individuals in each diametric class of the mangrove and freshwater swamp of the El Castaño Wetlands System.
En los humedales herbáceos, T.domingensis domina en el tular con 85.2 % de IVI; los valores del IVI están por debajo del 20 % en el resto de las especies. En el pastizal inundable P. conjugatum es la especie dominante con 21.57 % de IVI seguida por Solanum hirtum con13.62 % y el resto; de las especies tiene valores de IVI menores al 10 % (Apéndice Digital 4, Tabla B y Tabla C).
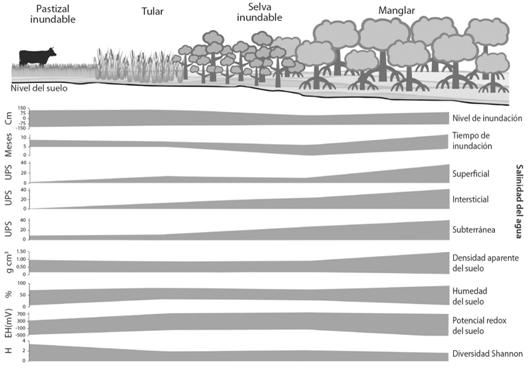
Distribución de la vegetación y su relación con las variables ambientales. Los resultados del análisis de escalamiento multidimensional con un stress 0.3136, muestran que la composición de las especies es diferente entre las comunidades vegetales por la distribución de las UM a lo largo del eje 1 (R2 = 0.5284), ordenando, al manglar, la selva inundable, el tular y al pastizal inundable a lo largo de un gradiente definido por la salinidad del agua subterránea e intersticial y potencial redox (Fig. 4).
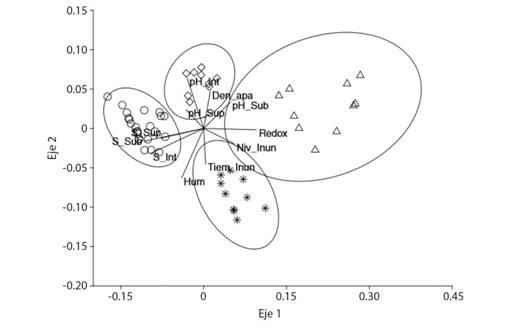

Fig. 4 Fig. 4 Análisis de escalamiento multidimensional no paramétrico de las especies de plantas usando distancia de Bray-Curtis que muestra la relación entre las comunidades vegetales y las variables ambientales. Salinidad superficial (S_Sup), salinidad intersticial (S_Int), salinidad subterránea (S_Sub), pH superficial (pH_Sup), pH intersticial (pH_Int), pH subterránea (pH_Sub), porcentaje de humedad (Hum), densidad aparente (Den_apa), nivel inundación (Niv_Inun), tiempo de inundación (Tiem_Inun), Potencial Redox (Redox). Non-metric multidimensional scaling analysis of plant species using Bray-Curtis distance that shows the relationship between plant communities and environmental variables. Surperficial salinity (S_Sup), interstitial salinity (S_Int), groundwater salinity (S_Sub), surperficial pH (pH_Sup), interstitial pH (pH_Int), groundwater pH (pH_Sub), humidity percentage (Hum), bulk density (Den_apa), level flood (Niv_Inun), flood time (Tiem_Inun), Redox Potential (Redox).
El manglar, dominado por R. mangle, L. racemosa y A. germinans, ocupa el extremo izquierdo de la ordenación con los mayores valores de salinidad. La selva inundable, dominada por P. aquatica, con elementos de R. mangle y Z. conzattii, y el tular, compuesto por T. domingensis, como especie principal, y con menor presencia, Sesuviummaritimun y A. aureum, en el centro de la ordenación; y el pastizal inundable a la derecha con la menor salinidad. Las tres principales especies que lo componen son P. conjugatum, Solanum hirtum y B. decumbens. Por lo tanto, el eje 1 muestra un gradiente de salinidad y sobre el eje 2 (R2 = 0.2606) se establece un gradiente definido por el pH intersticial, y el porcentaje de humedad del suelo. El tular tiene los valores más bajos de pH y el más alto porcentaje de humedad del suelo, y el tiempo de mayor inundación se registra en el manglar y en el pastizal inundable (Fig. 4).
Distribución de las comunidades vegetales en el SHC a lo largo del gradiente ambiental definido principalmente por la salinidad (Fig. 5).
Discusión
El gradiente ambiental del SHC está definido por la salinidad, de mayor a menor, principalmente del agua subterránea, a través de los humedales de la costa hacia tierra adentro durante todo el año. El nivel de inundación, no presentó el gradiente como se esperaba, debido a que el manglar y el pastizal inundable, los humedales más alejados entre sí, mantienen el mismo tiempo de inundación y el nivel del agua durante la temporada de lluvias. Esto se explica por el origen de las aportaciones de agua al sistema. En el pastizal, el humedal más alejado del mar, la influencia marina es poca, el agua llega por la precipitación, el desbordamiento de los ríos y el agua subterránea, por lo que permanece inundado durante la temporada de lluvias y parte de la temporada de secas por los aportes del agua subterránea que continúan fluyendo. Por lo que la primera hipótesis se cumple parcialmente, puesto que sólo la salinidad presenta un gradiente de mayor a menor al aumentar los aportes de agua dulce. Con respecto al potencial redox, el pH, la densidad aparente y el porcentaje de humedad en el suelo, el gradiente que se observa está sujeto a cambios según la temporada del año.
La diferencia en la salinidad del agua entre los humedales es uno de los factores que limita la distribución, establecimiento y crecimiento de las especies. El agua intersticial y subterránea es la que tienen disponible las plantas. En el manglar la zonación de las especies depende de la salinidad intersticial, y cada especie tiene diferente rango de tolerancia y adaptaciones que les permite vivir hasta en 80 ups (López-Portillo et al., 2011). Los valores en el SHC para el manglar están dentro de los reportados para Veracruz (Flores-Verdugo et al., 2007) y Tabasco (Torres, Infante-Mata, Sánchez, Espinoza-Tenorio, & Barba, 2017). En el caso del tular y la selva inundable la salinidad es mayor a la observada comúnmente en estos humedales en Veracruz (Yetter, 2004; Flores-Verdugo et al., 2007), aunque se han reportado algunos valores más altos, como es el caso del tular por Moreno-Casasola et al. (2010) y las selvas inundables con presencia de R. mangle (Infante-Mata et al., 2011; Infante-Mata, Moreno-Casasola, & Madero-Vega, 2014) en la que la salinidad está dentro del rango registrado en el SHC. Estas diferencias en el Castaño se relacionan con la distancia en que se ubican los tulares y los remanentes de la selva inundable con respecto al mar y durante la temporada de secas es más evidente al presentar los mayores valores de salinidad que llegan por arriba de 15.00 ups. Es por tanto importante considerar la fragilidad de estas comunidades ante los procesos de intrusión salina como efecto del cambio climático (Herbert et al., 2015; Zhai et al., 2018).
El pastizal inundable es el humedal con menor influencia marina, la salinidad es poca, y presenta la mayor diversidad, con la menor diversidad le sigue la selva inundable y el tular y al final el manglar con mayor influencia marina y menor diversidad. Esto significa que al reducirse el estrés por salinidad la diversidad aumenta. El recambio de especies, es mayor entre el manglar y el pastizal inundable, los humedales más alejados y con mayor diferencia de salinidad, y entre el pastizal inundable y la selva inundable con la mayor diferencia de inundación. Por lo que la segunda hipótesis también se cumple parcialmente ya que se esperaba que el tular fuera más diverso que la selva. En el SHC el gradiente de mayor a menor salinidad del manglar hacia los pastizales inundables, es uno de los factores ambientales que están diferenciando la distribución de la vegetación. Por lo que los valores más altos de similitud están entre las comunidades vecinas, en este caso el manglar y la selva inundables, y el tular y el pastizal inundable.
Sin embargo, en el pastizal inundable la alta diversidad también está relacionada con las especies estacionales en los cuerpos de agua temporales durante las lluvias y el efecto del disturbio por el uso ganadero (Grime, 1973; Cingolani, Noy-Meir, & Díaz, 2005). En el pastizal inundable el ganado pasta durante 6 a 8 meses (sequía) evitando que la población de las especies nativas dominantes aumente y permitiendo que las poblaciones de otras especies crezcan (Cingolani, Noy-Meir, Renison, & Cabido, 2008) y al retirar el ganado durante al menos seis meses por la inundación, permite al suelo recuperarse (Moreno-Casasola, López-Rosas, & Rodríguez-Medina, 2012b; Rodríguez-Medina, Moreno-Casasola, & Yañez-Arena, 2017), lo que al final aumenta la diversidad. Por lo que mantener el hidroperiodo permite la eliminación y control de ciertas especies, evitando que las poblaciones de especies no nativas se expandan (Moreno-Casasola et al., 2018).
En relación con la distribución, el humedal más cercano al mar es el manglar, en el que se encuentran cuatro de las seis especies de mangle registradas para Chiapas, R. mangle y L. racemosa son las dos especies dominantes. La densidad (1 945 ind ha-1) y área basal (20.5 m2 ha-1) son bajas con respecto a lo observado por Tovilla-Hernández y Romero-Berny (2012) y Romero-Berny et al. (2019) para Chiapas. Considerando solo el área de la Encrucijada Tovilla-Hernández, Aguilar-López, Gordillo-Solís, Rojas-García y Vazquez-Lule (2010) reportan una densidad similar, aunque un área basal mayor. Comparando con los manglares del Golfo de México la dominancia, la densidad y el área basal es diferente. En Alvarado y la Mancha, Veracruz, dominan Avicennia germinans y L. racemosa, con una densidad menor y un área basal mayor (Agraz et al., 2011; Rodriguez-Zuñiga, Ramírez-García, & Gutiérrez-Granado, 2011) y en la Laguna Mecoacán, Tabasco, domina A. germinans, con una densidad mayor y menor área basal que en el SHC (Torres et al., 2017). Las diferencias de estructura entre los manglares de Chiapas, se relacionan con las variables ambientales como la salinidad, el hidroperiodo, los nutrientes y los impactos antrópicos, en el caso de las diferencias con el Golfo de México también se tienen que considerar la ubicación latitudinal, el clima, y la geomorfología (López-Portillo & Ezcurra, 2002; López- Portillo et al., 2011; Romero-Berny et al., 2019).
Las especies registradas en el manglar del SHC, coinciden con las reportadas por Tovilla et al. (2007), excepto por el mangle amarillo, Rhizophora harrisonii de distribución limitada en la Encrucijada y Phyllanthus elsiae. También se tienen especies que no están reportadas para el manglar como: C. sicyoides, C. oaxacana, Hippomane mancinella, aunque se distribuyen en La Encrucijada, (SEMARNAP, 1999). La mayor parte de las especies que acompañan al manglar se establecen en los suelos elevados o en los sitios con influencia de agua dulce.
Siguiendo con la distribución de los humedales, atrás de los manglares están las selvas inundables dominadas por P. aquatica, que difieren en estructura con las estudiadas en Veracruz y Tabasco, la densidad es mayor, el área basal es menor, la riqueza y la diversidad son menores (Ascencio, 1994; Infante-Mata et al., 2011; Rincón, 2014). Con respecto a la composición Chiapas y Veracruz comparten 10 especies. Resalta la presencia de Zygia conzattii en Chiapas, con un valor alto de importancia y su ausencia en Veracruz. En ambas selvas hay palmas, en el SHC, S. mexicana y en Veracruz: Attalea liebmannii y Roystonea dunlapiana. Las selvas inundables de México son afines a las de Centro y Sur América, por los géneros y especies en común, (Infante-Mata et al., 2011). Moreno-Casasola, Infante-Mata y López-Rosas (2012a), consideran que también son diversas en composición, debido a la mayor heterogeneidad ambiental de las costas de México (Contreras-Espinoza & Warner, 2004; López-Portillo et al., 2011).
En el SHC los tulares son dominados por T. domingensis, ocupando amplias extensiones que colindan con los manglares y las selvas inundables en la zona del delta del río San Nicolás y tierra adentro colindan con los humedales modificados como los pastizales inundables, tierras de cultivo temporal, o por asentamientos humanos. Al igual que los tulares de Veracruz, dominados por T. domingensis, la composición de especies es baja. Algunas de las especies que comparten son C. giganteus, P.foetida y F. clausum (Peralta-Peláez &Moreno-Casasola, 2009; Moreno-Casasola et al., 2010).
Los pastizales inundables son los de menor influencia marina en el SHC, son humedales modificados para el uso ganadero. En éstos se observan parches remanentes de vegetación original y colindan principalmente con los tulares. Tienen la mayor riqueza y diversidad de los humedales de estudio y también con respecto a los pastizales inundables de Veracruz estudiados por Travieso-Bello, Moreno-Casasola y Campos (2005) y Rodríguez-Medina et al. (2017). Es una comunidad de herbáceas, dominada por el pasto nativo P. conjugatum, especie frecuente en sitios inundados. Hay otros pastos nativos como P. virgatum, Hymenachneamplexicaulis, y tres especies de pastos exóticos, siendo B. decumbens, el de mayor presencia. En SHC, en los pastizales inundables se mantiene la hidrología y no se desecan, se ha intentado sembrar pasto para forraje, sin embargo las condiciones permiten que siga dominando P. conjugatum.
A parte del pastizal inundable en los otros humedales también se registraron especies no nativas, como plántulas y juveniles de la palma de aceite, E. guineensis, en las selvas inundables y los suelos elevados del manglar. Esta especie es catalogada como invasora por la CONABIO (2017) e Invasive Species Specialist Group ISSG (2015). Se desconocen los efectos que puede tener en los ecosistemas costeros naturales, y su presencia va en aumento (Martínez, Castellanos, & Guerrero, 2016). En el tular se registró, B.decumbens, que consideramos no ha prosperado por su poca tolerancia a la inundación (Holman, Rivas, Argel, & Pérez, 2004).
Comparando, la costa de Chiapas en el Pacífico con el Golfo de México, este último tiene un mayor espacio de la planicie de inundación, hay una mayor heterogeneidad ambiental, que puede explicar la mayor diversidad de especies en humedales como la selva inundable, en relación con los que existen en la costa de Chiapas (Infante-Mata et al., 2011; López-Portillo et al., 2011; Moreno-Casasola et al., 2012a). La distribución de la vegetación y el funcionamiento de los humedales costeros en parte se explican por el gradiente de los factores ambientales como la salinidad, sin embargo, para entender las diferencias hay que considerar las características hidrogeomorfológicas de cada sitio, que se reflejan en la extensión y el ensamblaje de los ecosistemas costeros (Phillips, 2018). Sin embargo, en México, aún faltan estudios para terminar de comprender totalmente el funcionamiento en los humedales costeros.
En conclusión el gradiente de salinidad de mayor a menor influencia marina es uno de los factores que definen la distribución, composición, diversidad y estructura de la vegetación en el SHC. Se esperaba un gradiente similar de la inundación; sin embargo, esta condición no se cumplió por la dinámica hidrológica que presenta los mayores aportes de agua a los extremos del sistema. La diversidad de cada humedal responde a los cambios en la salinidad y el recambio de especies, diversidad beta, es evidente entre algunos humedales, por lo que existe un cambio de la vegetación que se relaciona con el gradiente ambiental. Finalmente la distribución de la vegetación en el SHC siguen los patrones de organización típica de los humedales costeros tropicales, (Zedler & Callaway, 2001; Moreno-Casasola et al., 2012a) del litoral hacia tierra adentro se observan los manglares, las selvas inundables y humedales herbáceos, en este caso los tulares y pastizales inundables.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.
See Digital Appendix at: / Ver Apéndice digital en: revistas.ucr.ac.cr
