











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Hasta la fecha se han descrito alrededor de 270 especies del género Liolaemus (Abdala & Quinteros, 2014; Gutiérrez et al., 2018; Uetz, 2019). Dentro del género se reconocen dos subgéneros: Eulaemus y Liolaemus “sensustricto”. A su vez, dentro de estos se reconocen grupos divididos en varios subgrupos, incluido el grupo de L.montanus (Abdala, Acosta, Cabrera, Villavicencio, & Marinero, 2009; Abdala & Juárez-Heredia, 2013; Abdala & Quinteros, 2014). Actualmente, en Perú se conocen 21 especies de este género, de las cuales 13 pertenecen al grupo antes mencionado (Gutiérrez et al., 2018). Particularmente, en el departamento de Arequipa se conocen solo cinco especies de Liolaemus (Cei & Péfaur, 1982; Villegas, 1990; Laurent, 1998; Aguilar, Wood, Belk, Duff, & Sites, 2017). Sin embargo, este número podría incrementarse con más investigaciones en la zona, debido a la falta de conocimiento de la biodiversidad (Zeballos, López, Villegas, Jiménez, & Gutiérrez, 2002; Gutiérrez, Villegas, López, & Quiroz, 2010; Valladares, 2011; Gutiérrez et al., 2018).
Actualmente, en Perú se han descrito las siguientes especies: Liolaemus annectens, Liolaemus etheridgei, Liolaemus insolitus y Liolaemus tacnae en el departamento de Arequipa; Liolaemus williamsi, Liolaemus polystictus, Liolaemus melanogaster, y Liolaemus wari en el departamento de Ayacucho; Liolaemus signifer en el departamento de Puno. La mayoría de estos organismos pertenecen al grupo de Liolaemus montanus, con excepción de L. tacnae, L. walkeri y L. wari, las cuales pertenecen a los subgrupos L.alticolor - bibroni y L. ornatus del grupo L. boulengeri (Etheridge, 1995; Abdala, 2007; Lobo, Espinoza, & Quinteros, 2010; Quinteros, 2012; Quinteros, 2013; Aguilar et al., 2013; Abdala & Quinteros, 2014).
Teniendo lo anterior en cuenta, la lagartija que describiremos es asignada al grupo de Liolaemusmontanus por presentar las sinapomorfias propuestas por Etheridge (1995) para este grupo, particularmente una hipertrofia del músculo tibialis anticus (Abdala, Abdala, & Tulli, 2006), originada por un proceso posterior distal de la tibia en forma de navaja. En general, del total de especies del grupo de Liolaemus montanus conocidas en el sur de Perú y norte de Chile, solo L. insolitus, L. poconchilensis y L. reichei se distribuyen en zonas costeras en alturas menores a los 1 000 m.s.n.m. (Valladares, 2004; Gutiérrez et al., 2018; Valladares-Faundez, Etheridge, & Simón-Abdala, 2018).
Aunque estas últimas son fenéticamente similares con la especie que se describe en este trabajo, los análisis moleculares indican que no hay relación cercana. Asimismo, existen unas formas aún no descritas de Liolaemus para la zona de Nazca en el departamento de Ica, y en el norte del departamento de Arequipa (Aguilar et al., 2017), las cuales también difieren molecularmente de la nueva especie. De igual manera, la nueva especie de Liolaemus, posee una combinación de caracteres que lo distinguen del resto de los Liolaemus descritos.
A fin de poner a prueba la hipótesis que le dé validez a la especie que se describe en este trabajo, utilizamos el concepto general o unificado de especie. Este lo define como entidades que representan linajes históricos independientes o linajes divergentes de meta-poblaciones (De Queiroz, 1998). Nuestro criterio operacional para inferir el límite entre especies está basado en la elaboración de árboles filogenéticos moleculares y la caracterización morfológica, al igual que otras descripciones de nuevos Liolaemus (Ávila et al., 2017).
Materiales y métodos
Característicasmorfológicas: La descripción se basa en especímenes depositados en las colecciones herpetológicas del Museo de Historia Natural de la Universidad Nacional de San Agustín de Arequipa (MUSA, Arequipa, Perú), en el Museo de Historia Natural de la Universidad Mayor de San Marcos (MUSM, Lima, Perú) y en el Museo de Biodiversidad del Perú (MUBI; Cusco, Perú). Se revisaron un total de 18 ejemplares para la obtención de la descripción de la especie propuesta desde las instalaciones del Museo de Historia Natural de la Universidad Nacional de San Agustín de Arequipa. Los ejemplares estaban fijados en formol al 10 % y preservados en etanol al 70 %. Las coordenadas del sitio de recolecta fueron tomadas con un GPS Garmín eTrex. Mientras tanto, los datos georreferenciados para la preparación del mapa se manejaron con el programa QGIS (QGIS Development Team, 2017).
En general, los caracteres descritos fueron los comúnmente utilizados para Liolaemus citados por Laurent (1986), Villegas (1990), Lobo y Espinoza (1999), Pincheira-Donoso y Núñez (2005), Abdala (2007), Abdala y Juárez-Heredia (2013). Las medidas fueron tomadas en gabinete con un calibrador digital de precisión 0.01 mm. Los conteos y observaciones de escamas fueron realizados a través de un estereoscopio con magnificación 4X. Mientras tanto, la descripción de coloración fue realizada en los organismos vivos en campo y con ayuda de fotografías. La terminología del patrón de coloración del cuerpo sigue a Lobo y Espinoza (1999) y Abdala (2007).
Extracción de ADN, PCR, secuenciación y alineamiento de secuencias: Las muestras de tejido muscular fueron obtenidas de lagartijas fijadas en etanol al 96 %. Para la extracción de DNA, se utilizó el Kit IBI © (Larson, 2015) siguiendo el protocolo del fabricante. El gen mitocondrial Cyt-b fue amplificado a partir del ADN total extraído utilizando los primers IguaCytob F2 (5’-CCACCGTTGTTATTCAACTAC-3’) e IguaCytob R2 (5’-GGTTTACAAGACCAATGCTTT-3’). Las condiciones del PCR fueron las siguientes: 94 °C durante cinco min luego de 35 ciclos; 94 °C durante 30 s; 50 °C durante 30 s; 72 °C durante 90 s; y finalmente 72 °C durante cinco min (Corl, Davis, Kuchta, Comendant, & Sinervo, 2010; Aguilar et al., 2017).
Posteriormente, los amplicones fueron secuenciados por MACROGEN - USA.
Para este proceso, las condiciones del PCR fueron las siguientes: 5 min de precalentamiento a 94 °C; 35 ciclos por 30 s de desnaturalización a 94 °C; 30 s de hibridación a 50 °C; 90 s de elongación a 72 °C; finalmente, se le adicionó 5 min de extensión final a 72 °C (Corlet al., 2010; Aguilar et al., 2017). Los amplicones obtenidos fueron secuenciados por la empresa MACROGEN - USA.
Análisis filogenético: Las secuencias obtenidas (Tabla 1) fueron comparadas con las secuencias homólogas reportadas en el GENBANK (Tabla 2) mediante la herramienta BLAST (http://blast.ncbi.nlm.nih.gov/Blast), según lo sugerido por Molina, Barreto & Giraldo (2014). Se consideraron principalmente las especies del género registradas para el departamento de Arequipa y la zona sur del Perú y utilizadas en los últimos estudios Aguilar et al. (2017). Las 48 secuencias nucleotídicas usadas en este análisis fueron alineadas usando la extensión MUSCLE (Edgar, 2004) del programa MESQUITE versión 3.2© (Maddison & Maddison, 2017). Por otro lado, la elección del mejor modelo de evolución de nucleótidos fue realizada en el programa JModelTest Version 2.1.4 ©” (Darriba, Taboada, Doallo, & Posada, 2012), considerando los índices de BIC y AIC.
TABLA 1 TABLE 1 Secuencias obtenidas de los organismos mediante la técnica PCR y posterior secuenciación Sequences obtained from organisms using the PCR technique and sequencing
| Especie | Voucher | GenBank |
| Liolaemus balagueri sp. nov. | MUSA 5578 | MK568538 |
| Liolaemus balagueri sp. nov. | MUSA 5575 | MK568539 |
TABLA 2 TABLE 2 Secuencias homólogas obtenidas en GenBank para realizar comparación Homologous sequences obtained in GenBank for comparison purposes
| Especie | Voucher | GenBank |
| L. ornatus | MUSM31438 | KX826632.1 |
| L. poconchilensis | MUSM31545 | KX826637.1 |
| L. insolitus | MUSM31490 | KX826627.1 |
| L. insolitus | BYU50462 | KX826626.1 |
| L. ortizi | MUSM 31514 | KX826634.1 |
| L. ortizi | MUSM 31513 | KX826633.1 |
| L. thomasi | BYU50469 | KX826680.1 |
| L. thomasi | MUSM31516 | KX826681.1 |
| L. williamsi | MUSM31485 | KX826687.1 |
| L. williamsi | BYU50464 | KX826685.1 |
| L. williamsi | BYU50463 | KX826684.1 |
| L. williamsi | BYU50143 | KX826682.1 |
| L. melanogaster | MUSM31472 | KX826630.1 |
| L. melanogaster | BYU50151 | KX826628.1 |
| L. sp. Abratoccto | MUSM31468 | KX826672.1 |
| L. sp. Abratoccto | BYU50431 | KX826664.1 |
| L. sp. Abratoccto | MUSM31465 | KX826671.1 |
| L. sp. Abratoccto | MUSM31374 | KX826667.1 |
| L. sp. Abratoccto | MUSM31371 | KX826665.1 |
| L. sp. Abratoccto | MUSM31481 | KX826660.1 |
| L. sp.AbraApacheta | BYU50148 | KX826659.1 |
| L. sp.AbraApacheta | BYU50145 | KX826658.1 |
| L. polystictus | MUSM31451 | KX826642.1 |
| L. polystictus | MUSM31446 | KX826641.1 |
| L. robustus | MUSM31508 | KX826648.1 |
| L. robustus | MUSM31505 | KX826647.1 |
| L. robustus | BYU50483 | KX826643.1 |
| L. robustus | MUSM31504 | KX826646.1 |
| L. signifer | MUSM31447 | KX826657.1 |
| L. signifer | MUSM31434 | KX826654.1 |
| L. signifer | BYU50357 | KX826651.1 |
| L. signifer | BYU50350 | KX826649.1 |
| L. signifer | BYU50355 | KX826650.1 |
| L. signifer | MUSM31443 | KX826656.1 |
| L. signifer | BYU50444 | KX826652.1 |
| L. signifer | MUSM31437 | KX826655.1 |
| L. signifer | MUSM29110 | KX826653.1 |
| L. etheridgei | BYU50497 | KX826662.1 |
| L. etheridgei | BYU50494 | KX826620.1 |
| L. etheridgei | BYU50495 | KX826621.1 |
| L. annectens | BYU50491 | KX826617.1 |
| L. annectens | BYU50486 | KX826615.1 |
| L. annectens | BYU50489 | KX826616.1 |
| L. sp.Nazca | MUSM31524 | KX826677.1 |
| L. sp.Nazca | MUSM31523 | KX826676.1 |
| L. sp.Nazca | BYU50472 | KX826673.1 |
| L. sp.Nazca | BYU50508 | KX826675.1 |
| L. sp.Nazca | BYU50507 | KX826674.1 |
Por su parte, el análisis de inferencia bayesiana (IB) fue corrida en Mr. Bayes v.3.2.6©” (Huelsenbeck & Ronquist, 2001). Todos los anteriores se realizaron dos veces durante diez millones de generaciones cada uno. Se obtuvo una desviación estándar promedio final de las frecuencias divididas por debajo de 0.05, muestreando árboles cada 1 000 generaciones y utilizando cuatro cadenas simultáneas (una fría y tres calientes) en cada ejecución. Las convergencias de las cadenas a la distribución estacionaria se confirmaron utilizando Tracer v1.6 (Rambaut, Suchard, Xie, & Drummond, 2014) y se descartaron los primeros mil árboles muestreados que no estaban dentro de la distribución estacionaria de las probabilidades de registro. Los árboles y las probabilidades posteriores se resumieron utilizando el método de consenso de la “mayoría del 50 %”(Huelsabeck & Ronquist, 2001; Wilcox, Zwickl, & Heath, 2002).
Establecimiento de la distribución potencial y hábitat:Para la determinación del área de distribución potencial de la especie de estudio, se realizó el modelado de nicho ecológico a partir de variables ambientales y los registros de presencia de las especies. En cuanto a las variables ambientales, se utilizaron las 19 del programa Worldclim (Fick & Hijmans, 2017). También se emplearon variables adicionales como radiación solar, índices de vegetación (NDVI) y mapa de cobertura vegetal, las cuales fueron construidas a una resolución de 100 m.
Previamente, se realizó un análisis de correlación entre las variables por utilizar en el modelo con la herramienta NicheToolBox (Osorio-Olvera, 2016), la cual se ejecuta bajo la plataforma de R (R Core Team, 2016) que, asimismo, integra el software de modelación de máxima entropía - MAXENT (Phillips, Anderson, & Shapire, 2006). Los resultados para el modelo fueron evaluados utilizando medidas de desempeño sobre los valores de AUC (área bajo la curva ROC) con valores entre 0.5 (no diferencia del azar) a uno (puntuación mejor que el azar; Fielding & Bell 1997). Además, se realizó una evaluación de las variables más importantes para determinar cuál de estas predice mejor el nicho fundamental de la especie, para lo cual se utilizaron valores de AUC entre cero (no importante) y uno (importante).
Resultados
Liolaemus balagueri sp. nov.
(Tabla 3, 4 y 5; Figs. 2 y 3)
Análisis filogenético: El análisis de inferencia bayesiana se fundamenta en un data set final compuesto por 45 muestras de especies del grupo Liolaemus montanus, 2 muestras de la nueva especie y 1 muestra de L. ornatus del grupo “boulengeri” como “outgroup”. Se tuvo como modelo de evolución nucleotídica GTR + G + I. Esta filogenia bayesiana (Fig. 1) muestra a la especie descrita, Liolaemus balagueri sp. nov. (MUSA 5575, MUSA 5578), como clado monofilético (pp = 1), taxón hermano de la población de Liolaemus sp. en Nazca (pp = 1). Además, separa a Liolaemus balagueri sp. nov. de las poblaciones de L. insolitus y L. poconchilensis - a pesar de su similitud fenética - así como de las demás poblaciones altoandinas de Liolaemus. Por tanto, se considera que L. balagueri sp. nov. es una especie diferente a las anteriormente descritas para el grupo de L. “montanus” (pp = 1).
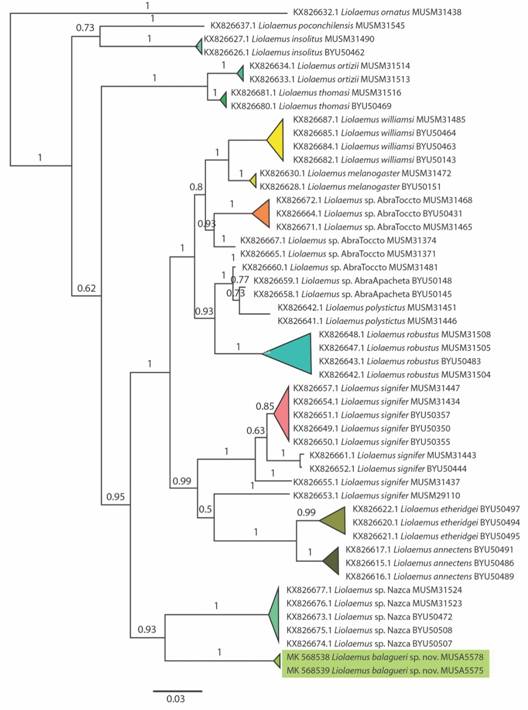
Fig. 1. Fig. 1 Árbol filogenético del grupo deL.montanus, incluido Liolaemus balagueri sp. nov. (MUSA 5575, MUSA 5578) Phylogenetic tree of the L. montanus group, including Liolaemus balagueri sp. nov. (MUSA 5575, MUSA 5578).
Análisis morfológico: Las diferencias morfológicas entre las poblaciones analizadas y el resto de las especies de Liolaemus, están plasmadas en el diagnóstico de la nueva especie y se detallan en la Tabla 3.
La combinación de caracteres de lepidosis, las medidas morfométricas y, principalmente, el patrón de coloración de machos y hembras, nos permiten realizar un diagnóstico claro y fundamentado. Además, aporta evidencia suficiente a favor de la descripción de esta nueva especie para la ciencia.En las Fig. 2A, Fig. 2B y Fig. 3A, Fig. 3B se pueden apreciar las vistas frontales y laterales de los machos esta especie (Liolaemus balaguerisp. nov.) y las hembras paratipo.

Fig. 2. Fig. 2 A. Vista lateral de un macho adulto de Liolaemus balagueri sp. nov. en su hábitat natural. B. Vista ventral del mismo ejemplar. A. Side view of a male adult of Liolaemus balagueri sp. nov. in the natural habitat. B. Ventral view of the same specimen.

Fig. 3. Fig. 3 A. Ejemplar paratipo hembra (MUSA 5578) en vista lateral. B. Vista ventral del mismo ejemplar A. Side view of the female paratype (MUSA 5578) in side view. B. Ventral view of the same specimen.
TABLA 3 TABLE 3 Principales medidas y variaciones entre especies del grupo de L. balagueri sp. nov. Main measurements and variations between the species of the group L. balagueri sp. nov.
| L. audituvealtus | L. sp nov. | L. reichei | L. insolitus | L. poconchilensis | L. torresi | |
| (N = 2) | (N = 18) | (N = 3) | (N = 10) | (N = 4) | (N = 8) | |
| Largo hocico-cloaca | 47.2 (47.5) 47.8 | 50.9 (59.1) 66.5 | 41.5 (47.7) 50.8 | 50.3 (59.4) 65.6 | 47.17 (51.7) 53.83 | 53.8 (58.1) 64 |
| Longitud de cola | 45.3 (46.3) 47.5 | 48.7 (59.4) 80.0 | 35.7 (39.5) 43.1 | 45.4 (51.7) 61.9 | 42.66 (47.2) 53.65 | 58.8 (57.4) 74 |
| Largo de la cabeza | 11.4 (11.6) 11.7 | 12.4 (14.1) 17.1 | 10 (10.9) 11.5 | 11.6 (13.6) 14.9 | 11.34 (13.1) 14.16 | 13 (13.6) 14.5 |
| Ancho de la cabeza | 9.6 (9.8) 10.3 | 9.8 (11.2) 13.6 | 8.3 (8.5) 9.7 | 9.5 (11.5) 12.4 | 9.51 (10.9) 11.53 | 10.3 (11.0) 11.7 |
| Longitud de la extremidad anterior | 27.1 (28.6) 28.7 | 21.0 (22.9) 25.1 | 21.4 (22.6) 24.4 | 26.1 (29.1) 31.5 | 22.2 (24.8) 27.5 | 26.3 (28.8) 30.5 |
| Longitud de la extremidad posterior | 37.1 (38.8) 40.6 | 29.1 (32.4) 35.9 | 30.1 (31.7) 33.4 | 37.4 (41.2) 44.3 | 31.15 (34.1) 32.62 | 38.6 (40.1) 42.2 |
| Escamas supralabiales | 7 (8) 9 | 5 (6.0) 7 | 9 (9) 9 | 7 (7.5) 9 | 12 | 10 |
| Escamas Infralabiales | 5 (5.5) 6 | 5 (5.9) 7 | 6 (6.7) 8 | 5 (6.5) 7 | 8 | 6 (6.6) 7 |
| Escamas Lorilabiales | 7 (8) 9 | 5 (6.9) 8 | 8 (8.7) 9 | 7 (8.2) 10 | 14 | 8 (8.4) 9 |
| Escamas alrededor del medio cuerpo | 74 (74.5) 75 | 52 (54.0) 56 | 43 (45) 47 | 51 (55.4) 58 | 57 (59.4) 62 | 64 (66.6) 72 |
| Escamas dorsales | 78 (82.5) 87 | 50 (55.1) 68 | 50 (51.7) 54 | 56 (62.9) 68 | 57 (61.2) 64 | 70 (74.4) 80 |
| Escamas ventrales | 86 (90.5) 95 | 65 (73.4) 79 | 70 (73.9) 78 | 77 (79.6) 84 | 86 (90.6) 97 | |
| Poros precloacales en hembras | 0 | 0 | 0 (1.3) 3 | 0 (1.3) 2 | 0 (1.5) 2 | |
| Poros precloacales en machos | 5 | 3 (5.08) 7 | 4 | 5 (6.2) 7 | 4 (4.6) 5 | 3 |
Holotipo: Se trató de un macho adulto (MUSA 5575) recolectado a 3.17 km, al oeste de la carretera Majes - Camaná antes de llegar a la quebrada del Toro (16°34`23.94” S & 72°33`43.02” W) a 1 058 m de altitud, Distrito de Quilca, Provincia Camaná, Departamento de Arequipa, Perú. Fue almacenado por Luis Villegas, Ling Huamaní y César Luque el 23 de julio 2017.
Paratipos: Se emplearon tres machos (MUBI 13206, MUSA 1772 - 1773) y cuatro hembras (MUSA 1774, MUBI 13207-13209), recolectados en la misma localidad del holotipo en las coordenadas 16°34’23” S & 72°33’43” W por Roberto Gutiérrez, Aarón Quiroz y Luis Villegas el 11 de agosto de 2010. Un macho (MUSM 39195) fue recolectado en la misma localidad que el holotipo, particularmente las coordenadas 16°34’11.96” S & 72°34’00.53” W por César Luque, Luis Villegas y Anthony Pauca el 02 de abril del 2017. Asimismo, otro macho (MUBI 16483) fue recolectado en la misma localidad que el holotipo en las coordenadas 16°34’19.21” S & 72°33’53.95” W por Luis Villegas, Ling Huamaní y Cesar Luque el 02 de abril de 2017.
De la misma manera, cinco machos (MUSA 5576-5577, MUBI 16484, MUSM 39193-39194) fueron recolectados en la misma localidad que el holotipo por Luis Villegas, Ling Huamaní y César Luque el 23 de julio de 2017, así como una hembra (MUSA 5578) recolectada por Luis Villegas, Ling Huamaní y César Luque el 08 de septiembre de 2017. En las lomas de La Chira, Distrito de Camaná, Provincia de Camaná en el departamento de Arequipa, se recolectaron dos machos (MUSA 5579 y MUSM 39192) por Luis Villegas, César Luque, Raquel Ruiz y Anthony Pauca, el 16 de diciembre de 2017.
Diagnóstico: La especie identificada fue asignada al grupo o serie de Liolaemus montanus (Etheridge, 1995; Schulte, Macey, Espinoza, & Larson, 2000) porque presenta un proceso posterior distal en la tibia - en forma de navaja - asociado a la hipertrofia del músculo tibialis anticus (Etheridge, 1995; Abdala, Abdala, & Tulli, 2006). Fue denominada como Liolaemus balagueri sp. nov. Dentro de la serie o grupo de L. montanus, se diferencia de las especies del grupo de L. boulengeri (Abdala, 2007; Schulte et al., 2000) por tener escamas de igual tamaño en la parte posterior del muslo. Por un lado, en L. montanus, se diferencia de L. annectens, L. aymararum, L. cazianiae, L. chlorostictus, L. dorbignyi, L. fabiani, L. filiorum, L. forsteri, L. foxi, L. gracielae, L. huayra, L. huacahuasicus, L. igneus, L. inti, L. jamesi, L. juanortizi, L. melanogaster, L. montanus, L. multicolor, L. nigriceps, L. orientalis, L. pantherinus, L. pachecoi, L. patriciaiturrae, L. pleopholis, L. polystictus, L. puritamensis, L. robustus, L. scrocchii, L. signifer, L. tacora, L. thomasi, L. vulcanus, L. vallecurensis, y L. williamsi por ser especies de mayor tamaño. Por ejemplo, el LHC de L. balagueri sp. nov. oscila entre 66.50 y 75 mm.
Por otro lado, se diferencia de L. andinus, L. audituvelatus, L. cazianiae, L. chlorostictus, L. eleodori, L. erguetae, L. fabiani, L. forsteri, L. foxi, L. gracielae, L. halonastes, L. insolitus, L. islugensis, L. jamesi, L. molinai, L. manueli, L. nigriceps, L. omorfi, L. patriciaiturrae, L. pleopholis, L. poconchilensis, L. poecilochromus, L. porosus, L. reichei, L. robertoi, L. rosenmanni, L. ruibali, L. schmidti, L. scrocchii, L. tacora, L. torresi, L. vallecurensis y L. vulcanus porque tienen escamas dorsales del cuerpo yuxtapuestas y sin quilla, escamas con disposición subimbricada y presencia de una leve quilla. Estos caracteres particulares también lo diferencian de L. aymararum,L. disjunctus, L. etheridgei, L. fittkaui, L. griseus, L. huacahuasicus, L. montanus, L. orko, L. ortizi, L. pulcherrimus, L. signifer, L. thomasi y L. williamsi que tienen las escamas imbricadas y con quilla evidente.
El número de escamas alrededor del cuerpo en Liolaemus balagueri sp. nov., varía entre 52 y 56 ( = 54.3), carácter que lo diferencia las especies del grupo que tienen más de 60 escamas, como L. andinus, L. audituvelatus, L. cazianiae, L. chlorostictus; L. duellmani, L. eleodori, L. erguetae, L. forsteri, L. foxi, L. gracielae, L. griseus, L. halonastes, L. inti, L. islugensis, L. manueli, L. molinai, L. multicolor, L. nigriceps, L. orientalis, L. orko, L. ortizi, L. patriciaiturrae, L. pleopholis, L. poconchilensis, L. poecilochromus, L. polystictus, L. porosus, L. pulcherrimus, L. robustus, L. robertoi, L. rosenmanni, L. ruibali, L. signifer, L. torresi, L. schmidti, L. vallecurensis y de L. reichei que tiene menos de 50 escamas alrededor del cuerpo.
El número de escamas ventrales entre la mental y el borde de la cloaca en la nueva especie varía entre 65 y 79 mm ( = 72.4). El mayor tamaño se ha identificado en L. ortizi (53-65 mm, = 61.1). También, esto lo diferencia de otras especies que tienen más de 80 escamas ventrales como L. andinus, L. annectens, L. audituvelatus, L. cazianiae, L. chlorostictus, L. eleodori, L. erguetae, L. foxi, L. gracielae, L. hajeki, L. halonastes, L. huayra, L. inti, L. islugensis, L. multicolor, L. nigriceps, L. pachecoi, L. patriciaiturrae, L. pleopholis, L. poecilochromus, L. porosus, L. robertoi, L. rosenmanni, L. signifer, L. torresi y L. vallecurensis.
En general, la combinación de los estados de caracteres presentes en el patrón de coloración dorsal y ventral lo diferencian claramente de todas las especies del grupo de L. montanus. Sobresale particularmente el patrón de coloración dorsal y sus grandes manchas paravertebrales y laterales de forma circular o de ocelos, con borde oscuro evidente y centro claro o verde. Estos caracteres son únicos en todo el género (Fig. 2A, Fig. 2B, Fig. 3A, Fig. 3B).
Diagnosis: This species is assigned to the group or series called L. montanus (Etheridge, 1995; Schulte, Macey, Espinoza, & Larson, 2000) because it presents a posterior distal process in the tibia (in the form of a knife) associated with the hypertrophy of the tibialis anticus muscle anticus (Etheridge, 1995; Abdala, Abdala, & Tulli, 2006). Within this group, it differs from the species L. boulengeri (Abdala, 2007; Schulte et al., 2000) for having scales of the same size on the back of the thigh. Within the overall group, it differs in size (they are larger) from L. annectens, L. aymararum, L. cazianiae, L. chlorostictus, L. dorbignyi, L. fabiani, L. filiorum, L. forsteri, L. foxi, L. gracielae, L. huayra, L. huacahuasicus, L. igneus, L. inti, L. jamesi, L. juanortizi, L. melanogaster, L. montanus, L. multicolor, L. nigriceps, L. orientalis, L. pantherinus, L. pachecoi, L. patriciaiturrae, L. pleopholis, L. polystictus, L. puritamensis, L. robustus, L. scrocchii, L. signifer, L. tacora, L. thomasi, L. vulcanus, L. vallecurensis and L. williamsi. For example, the LHC of L. balagueri sp. nov. ranges from 66.50 to 75 mm.
On the other hand, it differs from L. andinus, L. audituvelatus, L. cazianiae, L. chlorostictus, L. eleodori, L. erguetae, L. fabiani, L. forsteri, L. foxi, L. gracielae, L. halonastes, L. insolitus, L. islugensis, L. jamesi, L. molinai, L. manueli, L. nigriceps, L. omorfi, L. patriciaiturrae, L. pleopholis, L. poconchilensis, L. poecilochromus, L. porosus, L. reichei, L. robertoi, L. rosenmanni, L. ruibali, L. schmidti, L. scrocchii, L. tacora, L. torresi, L. vallecurensis and L. vulcanus in their dorsal scales of the body, which are juxtaposed and without keel; their scales with sub-imbricated arrangement and the presence of a slight keel in the new species. These particular traits also separate it from L. aymararum,L. disjunctus, L. etheridgei, L. fittkaui, L. griseus, L. huacahuasicus, L. montanus, L. orko, L. ortizi, L. pulcherrimus, L. signifer, L. thomasi and L. williamsi, which have imbricated scales and an evident keel.
The number of scales around the body in Liolaemus balagueri sp. nov. varies between 52 and 56 ( = 54.3). This trait differentiates the species from the group that has more than 60 scales, which includes L. andinus, L. audituvelatus, L. cazianiae, L. chlorostictus; L. duellmani, L. eleodori, L. erguetae, L. forsteri, L. foxi, L. gracielae, L. griseus, L. halonastes, L. inti, L. islugensis, L. manueli, L. molinai, L. multicolor, L. nigriceps, L. orientalis, L. orko, L. ortizi, L. patriciaiturrae, L. pleopholis, L. poconchilensis, L. poecilochromus, L. polystictus, L. porosus, L. pulcherrimus, L. robustus, L. robertoi, L. rosenmanni, L. ruibali, L. signifer, L. torresi, L. schmidti, L. vallecurensis and L. reichei. All of them have less than 50 scales around the body.
The number of ventral scales between the mental and the edge of the cloaca in varies between 65 and 79 ( = 72.4), higher than L. ortizi (53-65, = 61.1). Additionally, this number of scales is lower than in species that have more than 80 ventral scales such as L. andinus, L. annectens, L. audituvelatus, L. cazianiae, L. chlorostictus, L. eleodori, L. erguetae, L. foxi, L. gracielae, L. hajeki, L. halonastes, L. huayra, L. inti, L. islugensis, L. multicolor, L. nigriceps, L. pachecoi, L. patriciaiturrae, L. pleopholis, L. poecilochromus, L. porosus, L. robertoi, L. rosenmanni, L. signifer, L. torresi and L. vallecurensis.
The combination of characteristics observed in the dorsal and ventral coloring pattern clearly differentiate it from all the species of the L. montanus group. Particularly, the dorsal coloring pattern, large paravertebral and lateral spots of circular or ocelli shape, with border dark evident and light or green center stand out. These traits are unique in the whole genus (Fig. 2A, Fig. 2B, Fig. 3A, Fig. 3B).
Descripción del holotipo: Consiste en un macho adulto (MUSA 5575). El ejemplar es de tamaño mediano (largo hocico-cloaca = 63.9 mm), figura esbelta, con la cabeza un poco más ancha que el cuello. Presenta el pliegue auricular y longitudinal. Esta zona también está compuesta de escamas granulares que se conectan a las escamas granulares de la parte dorsal en la inserción de los miembros superiores al cuerpo. El cuerpo es semicilíndrico y deprimido dorsoventralmente.
La cabeza es regular con el perfil triangular ligeramente convexo, un largo de 15.9 mm, ancho de 13.3 mm y una altura de 11.6 mm. La distancia del tronco (axila-ingle = 29.3 mm) es 0.46 veces el tamaño del cuerpo y las extremidades anteriores y posteriores son relativamente cortas (23.9 y 35.2 mm, respectivamente). La cola es cilíndrica, está ensanchada en la base y termina en la punta con una medida 68.2 mm. El diámetro del ojo es de 3.4 mm. Por otro lado, la longitud oído-ojo es de 6.5 mm: el oído elíptico presenta una altura de 2.4 mm y un ancho de 0.8 mm. La distancia entre las narinas es de 4.2 mm, el largo de la escama subocular es de 2.8 mm y el largo de la hilera de poros precloacales es de 4.3 mm.
Por otra parte, la longitud del brazo corresponde a 8.2 mm, del antebrazo 7.0 mm y la mano 8.7 mm. La longitud del muslo es de 11.1 mm, la tibia 10.1 mm. y el pie 14.0 mm.Las escamas del dorso son de subyuxtapuestas a subimbricadas y ligeramente quilladas no mucronadas, con pequeñas escamas granulares (heteronotos) entre ellas. Las escamas de los miembros anteriores y posteriores son imbricadas, laminares y levemente quilladas en la parte dorsal, mientras que la parte ventral no presenta quillas. Asimismo, las escamas de la cola son más quilladas que el dorso del cuerpo. Por su parte, las escamas ventrales son lisas, imbricadas y ligeramente más grandes que las dorsales.
En cuanto a la cabeza, esta es de superficie rugosa, la escama interparietal irregular está rodeada de siete escamas de distinto tamaño y las placas parietales (dos) son más notorias. La escama rostral es de forma semirectangular, dos veces más ancha (2.4 mm) que alta (0.9 mm), la mental es semirectangular ligeramente más ancha (2.4 mm) que alta (1.3 mm). Las escamas supraoculares y frontal son un poco más grandes que las demás escamas cefálicas. Por otro lado, las escamas nasales se encuentran separadas de la rostral. Ellas también están divididas entre sí por cuatro escamas.
Se evidenció también una escama en el meato auditivo. Las escamas alrededor del medio del cuerpo son 54. Las escamas dorsales desde la línea imaginaria entre los bordes anteriores de los miembros posteriores hasta el occipucio también son 54 y las ventrales, desde la mental hasta la cloaca, son 74. Por su parte, las lamelas del cuarto dedo de la mano son 16 y las del pie 20.
Coloración: Presenta una cabeza dorsalmente de color marrón. Su área parietal es ligeramente más oscura que el resto de la cabeza. Las escamas son supralabiales e infralabiales de color marrón claro al igual que las escamas temporales. Presenta una línea transversal de color negro en los parpados. Ventralmente, la cabeza es de color crema con semicírculos paralelos negros que rodean a la escama mental que continúa hasta el sector gular en forma de jaspeada.
El dorso del cuerpo es de color marrón claro, con presencia de manchas paravertebrales de forma redondeada y borde oscuro seguido por una mancha posterior blanca. Lateralmente, tiene por encima de la línea ventrolateral un color marrón claro con manchas laterales verdes bordeadas por un color marrón oscuro. Por debajo de la línea ventrolateral, presenta un color crema con escamas de color verde, amarillo y naranja.
Ventralmente, el cuerpo es crema, con algunas escamas amarillas, naranjas y negras en los laterales del abdomen. Los miembros anteriores y posteriores a nivel dorsal son de color marrón claro con presencia de manchas negras. Mientras tanto, a nivel ventral presenta un color de blanco a crema. Su cola es dorsalmente de color marrón claro con presencia de unión de las manchas paravertebrales; ventralmente, es color crema. Los poros precloacales tienen una coloración anaranjado intenso.
Variación: Los patrones de coloración de los paratipos MUSA 5579 y MUSM 39192 son similares al holotipo al presentar manchas de coloración naranja al costado de las paravertebrales en el dorso. Los paratipos MUSA 39194 y MUSA 5577 muestran una coloración corporal dorsal marrón claro. Las hembras, en general, presentan una coloración más tenue (Fig. 3A). Por lo cual, puede observarse un dicromatismo sexual a nivel ventral, ya que las hembras tienen el vientre de color blanco con algunas escamas amarillas (Fig. 3B).
Con respecto a las medidas corporales, se obtuvo que las hembras son ligeramente de menor tamaño, a excepción de la longitud axila-ingle (Tabla 4). La escamación varía de 1 a 2 escamas en el conteo de lepidosis de cabeza y de 1 a 4 escamas a medio cuerpo en promedio (Tabla 5), dependiendo del sexo del individuo. Los poros precloacales solo se presentan en machos, mientras que los poros supernumerarios se presentan en ambos sexos al borde superior e inferior de la cloaca.
TABLA 4 TABLE 4 Principales medidas y variaciones entre hembras y machos de L. balagueri sp. nov. (N: número de individuos, DS: desviación estándar) Main measures and variations between females and males of L. balagueri sp. nov. (N: number of individuals, DS: standard deviation)
| Promedio machos | DS machos | Variación | N | Promedio hembras | DS hembras | Variación | N | |
| Largo hocico-cloaca | 61.12 | 4.42 | (52.31-66.58) | 13 | 57.15 | 4.24 | (50.99-62.7) | 5 |
| Largo de la cabeza | 15.12 | 1.32 | (12.98-17.14) | 13 | 13.23 | 0.74 | (12.46-14.4) | 5 |
| Ancho de la cabeza | 12.12 | 1.04 | (10.35-13.66) | 13 | 10.32 | 0.36 | (9.88-10.8) | 5 |
| Alto de la cabeza | 11.01 | 0.96 | (9.27-12.4) | 13 | 9.44 | 0.49 | (8.70-9.78) | 5 |
| Longitud cola | 65.85 | 10.12 | (53.15-80) | 8 | 53.03 | 3.70 | (48.78-55.6) | 3 |
| Base de la cola | 7.84 | 0.41 | (7.29-8.6) | 13 | 7.77 | 0.51 | (7.13-8.5) | 5 |
| Longitud axila- ingle | 26.35 | 2.37 | (21.88-30.27) | 13 | 26.24 | 3.58 | (20.85-30.9) | 5 |
| Longitud del fémur | 10.22 | 1.03 | (8.72-12) | 13 | 8.90 | 0.61 | (8.29-9.68) | 5 |
| Longitud de la tibia | 9.75 | 0.81 | (8.33-11.54) | 13 | 7.96 | 0.74 | (7.08-8.88) | 5 |
| Pie | 13.84 | 0.84 | (12.70-15.2) | 13 | 13.62 | 0.72 | (12.67-14.4) | 5 |
| Largo del brazo | 7.78 | 1.18 | (6.02-10.06) | 13 | 7.45 | 0.44 | (6.82-8) | 5 |
| Largo del antebrazo | 7.36 | 0.59 | (6.68-8.68) | 13 | 6.57 | 0.48 | (5.72-6.88) | 5 |
| Mano | 8.75 | 0.89 | (7.04-10) | 13 | 7.87 | 0.67 | (7.09-8.58) | 5 |
| Número de poros | 5.08 | 0.82 | (3.00-7) | 0 | 0 | 5 |
TABLA 5 TABLE 5 Principal conteo de escamas entre hembras y machos en L. balagueri sp. nov. (N: número de individuos, DS: desviación estándar) Main scale counts between females and males of L. balagueri sp. nov. (N: number of individuals, DS: standard deviation)
| Escamación | Promedio machos | DS machos | Escala | N | Promedio hembras | DS hembras | Escala | N |
| Escamas alrededor del medio cuerpo | 54.54 | 1.26 | (52-56) | 13 | 53.60 | 0.55 | (53-54) | 5 |
| Supralabiales del lado derecho | 6.23 | 0.59 | (5-7) | 13 | 5.80 | 0.45 | (5-6) | 5 |
| Infralabiales del lado derecho | 5.92 | 0.49 | (5-7) | 13 | 6 | 0 | (6-6) | 5 |
| Escamas en contacto con la interparietal | 6.92 | 1.25 | (5-9) | 13 | 7.20 | 1.78 | (5-10) | 5 |
| Escamas ventrales desde la mental a la cloaca | 71.23 | 2.38 | (65-74) | 13 | 75.60 | 2.60 | (72-79) | 5 |
| Escamas dorsales del occipucio al borde anterior de los miembros posteriores | 56.30 | 5.15 | (51-68) | 13 | 54 | 4.06 | (50-60) | 5 |
| Escamas auriculares diferenciadas del meato auditivo | 1.00 | 0 | (1-1) | 13 | 1 | 0 | (1-1) | 5 |
| Lamelas del dedo cuarto, pata trasera derecha | 20.69 | 0.94 | (20-23) | 13 | 20.60 | 0.55 | (20-21) | 5 |
| Lamelas dedo cuarto, pata delantera derecha | 16.31 | 0.85 | (16-19) | 13 | 15.8 | 0.48 | (15-16) | 5 |
Etimología: Dedicamos el nombre científico al Dr. Luis Balaguer Núñez (1964-2014), profesor principal e investigador incansable de la Cátedra de Ecología en la Universidad Complutense de Madrid, España. Particularmente, reconocemos su valiosa contribución al conocimiento del funcionamiento de los ecosistemas de las lomas de Atiquipa. Lamentablemente ya ha fallecido, pero dejó una gran escuela de futuros investigadores, tanto en Perú como en España.
Distribución: Los ejemplares estudiados de Liolaemus balagueri fueron encontrados en la zona alta de las lomas de Quilca (640-1 060 m) en el distrito de Quilca; en Samuel Pastor, en las lomas de La Chira (300-400 m) distrito de Mariscal Cáceres; y en las lomas de Ocoña (600-700 m.), distrito de Ocoña. Todas de las localidades pertenecen a la provincia de Camaná, departamento de Arequipa, Perú. La zona corresponde a la antigua Cordillera Costera, con altitudes entre los 400-1 000 m. (Fig. 4).
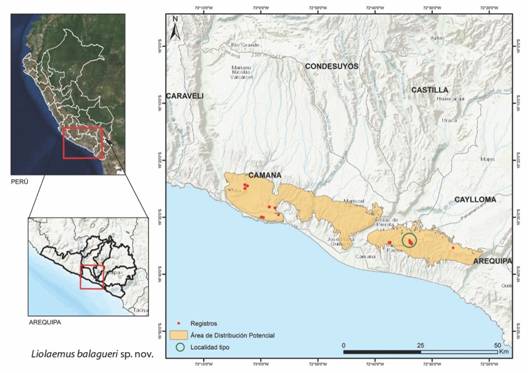
Fig. 4. Fig. 4 Distribución potencial de Liolaemus balagueri sp. nov. estimada a partir del análisis de 22 variables ambientales a 100 m de resolución y con el software Maxent. El círculo corresponde a la localidad tipo. Potential distribution of Liolaemus balagueri sp. nov. estimated from the analysis of 22 environmental variables at 100 m resolution and Maxent software. The circle corresponds to the type locality.
Se obtuvieron un total de 21 registros de la especie, los cuales fueron utilizados íntegramente en el modelo. Posterior al análisis de correlación, se determinó que las variables más significativas para la formación del modelo fueron: estacionalidad de la precipitación, radiación solar (14 000-14 500 lux), temperatura en el mes más frío (5-13 °C), precipitación anual (inferior a 10 mm) y cobertura vegetal (lomas). Esto concuerda con las características del hábitat en los que se realizaron los registros de la especie. Por ejemplo, la presencia de vegetación de lomas era común en los sitios concurridos por la especie, particularmente los bordes de la vegetación. Este hábitat depende en gran medida de la precipitación de la zona.
Por otro lado, el área estimada para la distribución de estas especies fue 711 km2, políticamente encontrados solamente dentro del departamento de Arequipa entre las provincias de Camaná y Caylloma. La variación de su altitud se encuentra entre los 300 a 1 060 m. La distribución más baja fue hacia el norte, próximo al área de Punta La Chira (Fig. 5). Los suelos de estas zonas presentan texturas limosas-arenosas y en algunas zonas hay presencia de rocas. Entre las barreras geográficas identificadas para su ampliación de distribución se encuentran la quebrada del río Quilca hacia el sur y la quebrada del río Ocoña hacia el norte.

Sobre la validación del modelo, según la prueba de AUC (área bajo la curva) se obtuvo un valor de 0.994, es decir, es un modelo mejor que el azar. Asimismo, al realizar una verificación del modelo en campo (la zona de Punta La Chira), se confirmó, mediante una visita de campo, que los sitios marcados con una alta probabilidad de encontrar a la especie presentaban individuos con facilidad.
Historia natural: El hábitat donde se encuentra Liolaemus balagueri es una zona alta de las lomas, con presencia de rocas y de sustrato arenoso-limoso con cobertura rala. La vegetación dominante está dada por pajonales de Cistanthe paniculata, Palaua dissecta, Oziroe biflora, Hierobotana inflata, Wedelia weberbaueri, Haageocereus decumbens y Plantago limensis. Las madrigueras de estos organismos se ubican en el sustrato arenoso-limoso, en el cual excavan pequeñas fosas del tamaño de su cuerpo. Ocasionalmente pueden utilizar madrigueras de alacranes y lagartijas de la especie Phyllodactylus gerrhopygus. Su coloración es críptica con respecto al suelo, lo que les permite camuflarse con facilidad; sin embargo, son fáciles de encontrar en ambientes abiertos, y con escasa cobertura vegetal (Fig. 5).
Discusión
El conocimiento de los reptiles de la costa sur de Perú es aún incipiente. Los listados publicados hasta la fecha de las especies de la familia Liolaemidae para Perú han sufrido de incertidumbres taxonómicas y grandes vacíos geográficos en las colecciones (Carrillo de Espinoza & Icochea, 1995; Aguilar et al., 2017). De modo que existen muchas otras aún sin describir (Gutiérrez et al., 2018). Por ello, la descripción de esta nueva especie contribuye al esclarecimiento de la diversidad real de las especies de Liolaemus para este país. Según los resultados, esta nueva especie (Liolaemus balagueri sp nov.) pertenece al grupo de L.montanus (Etheridge, 1995). Dentro de este compendio de organismos, los caracteres morfológicos la acercan a las especies pertenecientes o similares al ex género Phrynosaura (de donde derivan L. audituvelatus, L. insolitus, L. poconchilensis, L. reichei, L. stolzmanni, L. torresi), tal como lo indican los resultados de Etheridge (1995).
Sin embargo, los resultados moleculares presentados en este trabajo indican que L. balagueri sp. nov. no forma parte del mismo grupo monofilético al que pertenecen L. poconchilensis y L. insolitus. Por lo tanto, el ex género Phrynosaura - ahora sinonimizado con Liolaemus - puede presentarse en distintos nodos del grupo de Liolaemus montanus, como se muestran en los trabajos de Valladares, Etheridge, Schulte, Manríquez & Spotorno (2002), Aguilar et al.(2017) y Ruiz De Gamboa, Correa, Marambio-Alfaro, Riveros-Riffo, & Ortiz (2018).
Por otro lado, los resultados del nicho potencial de la nueva especie muestran cierta similitud con lo obtenido en el trabajo de Minoli, Morando & Ávila (2014), donde se resalta la importancia de la estacionalidad, la precipitación, isotermalidad y la temperatura mínima del mes más frío para el complejo de especies del género Liolaemus, en particular L.fitzingeri. No obstante, las conclusiones obtenidas en este trabajo se distinguen de Aguilar et al.(2013), donde resaltan la importancia de la precipitación en el periodo más húmedo y la isotermalidad para especies altoandinas del género Liolaemus del complejo de L.walkeri. Bajo la misma línea, Aguilar et al.(2017) propone un modelamiento del nicho potencial para algunas especies del grupo de Liolaemus montanus, el cual no presenta superposición con los nichos potenciales de Liolaemus balagueri sp. nov.
Por último, un punto a considerar es la gran cantidad de poblaciones innominadas que aún se encuentran en las lomas de las costas del centro y el sur de Perú, las cuales están relacionadas o fenéticamente similares a L. insolitus. La mayoría de estas poblaciones son asignadas en los museos como L. insolitus; sin embargo, un detallado y exhaustivo análisis morfológico podría evidenciar que son efectivamente poblaciones innominadas y seguramente tienen relación de parentesco entre sí y con las especies descritas de L. montanus. Por ello, podríanser especies sin describir formalmente para la ciencia (Aguilar et al., 2017). Lo anterior sucede a lo largo de las costas desde Arequipa hasta el sur de Perú. Además, al igual que L. balagueri sp. nov -la cual tiene un endemismo marcado- se han encontrado poblaciones cercanas a esta, cuyos datos morfológicos, moleculares (Fig. 3) y de nicho potencial (Fig. 4) pueden evidenciar la existencia de otras especies que deberían describirse formalmente. Por lo tanto, aún queda mucho trabajo taxonómico por hacer en este grupo de lagartijas de Perú.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

