











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
Coastal lagoons are enclosed or semi-enclosed shallow water bodies connected to the sea by one or more mouths (Kjerfve, 1994) through which water, particles and biota are exchanged. They are usually characterized by the presence of many habitats that show wide ranges of physical-chemical conditions, sustaining high biodiversity and productivity (Alongi, 1998). Moreover, these ecosystems are recognized for supporting life in a much wider extent than their own geographical limits, e.g. by providing nursery grounds for marine fish species, some of which are of commercial interest (Verdiell-Cubedo, Oliva-Paterna, Ruiz-Navarro, & Torralva, 2013; Tournois et al., 2017). Due to their coastal location, coastal lagoons are highly vulnerable to both natural and anthropogenic disturbances (Magalhães et al., 2019), especially in tropical zones, and are particularly affected by large-scale impacts associated with climate change, such as sea level rise, increased temperatures, or changes in sediment dynamics, among others (Pitacco, Mistri, & Munari, 2018).
The physical and chemical characteristics of the water and the sediments, and the biological populations display strong spatial gradients in coastal lagoons (Guelorget & Perthuisot, 1983; Giménez et al., 2014; Lamptey & Armah, 2008). The strong salinity spatial zonation is determined by the balance between the stratification and mixing of fresh and seawater. The water exchanges with the continent, the sea and the atmosphere modify the salinity of coastal lagoons, and tidal currents and wind are the main mixing processes. The capacity of populations to adapt to different salinity values results in a fauna arrangement partly defined by the salinity gradients (Blanchet, Montaudouin, Chardy, & Bachelet, 2005). When evaporation prevails, coastal lagoons present hypersaline conditions (salinity permanently above > 40 psu; sensuCan, Kevrekids, & Cihangir, 2009), under which salinity has strong control on the organisms’ distribution. High salinity values (ca. > 45 psu) are usually associated with low fauna density (Lamptey & Armah, 2008; Can et al., 2009; Joydas et al., 2015), unique fauna communities (Alongi, 1998; Joydas et al., 2011; Joydas et al., 2015), and the presence of opportunistic species (Marzano et al., 2010; Joydas et al., 2015). Yet, fauna distribution patterns result from complex interactions between several abiotic and biotic factors, and it is difficult to single out the effects of salinity (Thrush et al., 2003). In coastal lagoons, the heterogeneously distributed sediment characteristics and biogeochemical processes are considered as the main factors additional to salinity shaping the distribution of living organisms (e.g. Giménez et al., 2014), and environmentally and biologically distinct areas can usually be defined on the large scale (Carvalho et al., 2011). Daily and seasonal variability in the water exchange is another characteristic of coastal lagoons causing large fluctuations in the physico-chemical environment (Marzano et al., 2010). In shallow systems, atmospheric changes add up to such variability. As a consequence, the biota in coastal lagoons is characterized by a high level of adaptive evolution to stress and to environmental variations (Hernández-Alcántara, Medina-Cantú, Avilés-Díaz, & Solís-Weiss, 2014).
Macrobenthos is a dominant group in terms of abundance in coastal lagoons and participates in important ecological processes such as detritus decomposition and nutrient cycling (Carvalho et al., 2011). In such ecosystems, polychaetes often dominate the macrobenthic community (e.g. Hernández-Alcántara & Solís-Weiss, 1991, 1995; Lamptey & Armah, 2008; Carvalho et al., 2011; Hernández-Alcántara et al., 2014). Polychaetes have been identified as key prey of certain fish (e.g. Chi-Espínola & Vega-Cendejas, 2016), or decapod crustaceans (Albertoni, Palma-Silva, & Esteves, 2003; Chartosia, Kitsos, Tzomos, Mavromati, & Koukouras, 2010), and they are widely used as ecological indicators of natural or human-induced changes (Solis-Weiss et al., 2004). Thus, studies of their community structure and ecology may provide useful information on the food availability for species of importance to conservation or to fisheries, or on the conservation status of the ecosystem. Moreover, the structure of macrobenthic assemblages in estuarine systems is usually patchy (e.g. Hernández-Alcántara & Solís-Weiss, 1991; Edgar & Barret, 2002; Ysebaert & Herman, 2002; Meerhoff, Rodriguez-Gallego, Giménez, Muniz, & Conde, 2013; Magalhães & Bailey-Brock, 2014). Therefore, the identification of relevant distribution patterns will help explain the ecological processes or anthropogenic impacts structuring ecosystems (Underwood & Chapman, 1996; Ysebaert et al., 2003). In addition to salinity and sediment characteristics, polychaetes distribution is mainly driven by substrate type and food availability (Blanchet et al., 2005; Como & Magni, 2009; Carvalho et al., 2011).
This study is one of the few to address the community ecology of macrobenthic polychaetes in a coastal lagoon with a wide salinity range and with hypersaline conditions, with high taxonomic resolution. Specific aims of the study are to (1) describe the spatial distribution patterns of the macrobenthic polychaetes community in the Río Lagartos lagoon system during the rainy season, and to (2) identify the role of several abiotic variables as drivers of those patterns, with a special emphasis on salinity.
Materials and methods
Study area: Río Lagartos is a coastal lagoon laying parallel to the coast in the north-eastern Yucatán Peninsula, which opens to the SE Gulf of Mexico. It has a length of about 80 km and a total area of 98 km2. This lagoon is part of the Ría Lagartos Biosphere Reserve and is home to large numbers of species important to conservation. Besides, it serves as feeding area of many migratory birds (Luis Sauma personal observation) and the American flamingo (Phoenicopterus ruber Linnaeus, 1758), that also breeds in the lagoon (Espino-Barros & Baldassarre, 1989).
Seawater inputs to the lagoon take place through three mouths located to the west, and freshwater is only available from few small submarine groundwater point discharges typical from the Yucatán Peninsula calcareous karst substrate (Herrera-Silveira & Morales-Ojeda, 2009; Vera, Mariño-Tapia, & Enriquez, 2012), and from precipitation. Long water residence time (ca. 450 days), low depth (0.5 m), and excess evaporation over rain (Herrera-Silveira & Morales-Ojeda, 2009) result in a salinity gradient from west to east in which salinity values range from seawater levels (i.e. 34 psu) to values higher than 100 psu at the eastern limit (Valdes & Real, 2004; Chi-Espínola & Vega-Cendejas, 2016). Strong salinity changes are observed between the three different lagoon basins (i.e. from west to east: Río Lagartos, Las Coloradas and El Cuyo) (Vega-Cendejas & Hernández de Santillana, 2004), with associated changes in the composition and abundance of aquatic organisms such as fish (Vega-Cendejas & Hernández de Santillana, 2004; Peralta-Meixueiro & Vega-Cendejas, 2011) and algae (Ortegón-Aznar, González-González, & Sentíes-Granados, 2001). Hence, three main zones represented by the lagoon basins can be defined. Despite the ecological importance of the lagoon as part of a Biosphere Reserve, macrobenthos has barely been studied, and only one study including polychaetes information has been published from Río Lagartos (Kuk-Dzul, Gold-Bouchot, & Ardisson, 2012). In this study, low abundance and species richness was detected in the eastern hypersaline basin (i.e. El Cuyo).
Macrobenthos sampling: Sampling was carried out in September 2017 in the Río Lagartos coastal lagoon system, north-eastern Yucatán Peninsula (21°26’-21°38’ N; 87°30’-88°5’ W) (Fig. 1), as part of the Salinity Gradient Energy project of the Centro Mexicano de Innovación en Energía (CEMIE) Océano. Macrobenthos samples were collected from a total of 16 stations distributed along the lagoon considering the three main basins in the system: Río Lagartos to the west (Stations: 3, 6, 9, 12, 15), Las Coloradas in the middle (Stations: 20, 21, 22, 23, 25) and El Cuyo to the east (Stations: 27, 29, 30, 32, 35, 38) (Fig. 1). The distribution of the sampling stations according to the three different basins was based on previous studies of the distribution of physical-chemical parameters (Valdes & Real, 2004), fish (Vega-Cendejas & Hernández de Santillana, 2004), and algae (Ortegón-Aznar et al., 2001) in the lagoon. Those studies indicated that the three basins can be considered as environmentally and biologically distinct areas. Samples were collected at depths ranging from 0.5 to 1.5 m using a Ponar standard dredge (9” x 9” or 22.9 x 22.9 cm; 0.052 m2 grab operating area), or a cylindrical PVC core (6” diameter; 0.03 m2 operating area) when the substrate was too hard and the dredge did not penetrate the ground (Appendix 1). Two replicates per station were obtained, and sediments were sieved through a 500 μm mesh size to separate the macrofauna. The biological material was initially anesthetized with magnesium chloride, and preserved in 4 % formaldehyde. In the laboratory, samples were preserved in 70 % ethanol, polychaetes were sorted and identified, usually to species level, under a stereomicroscope (at x10-x40 Leica EZ4) or a microscope (at 20x-100x Nikon Eclipse Ci).
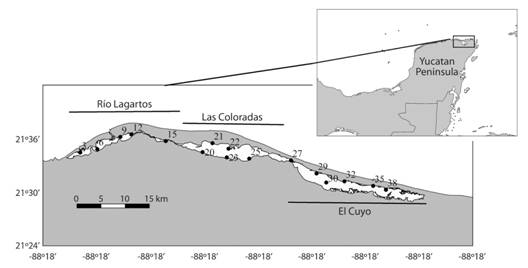
Fig. 1 Study area and sampling stations (3-38) in the Río Lagartos Lagoon, Yucatán Peninsula. The three basins are named after the names of the three fishermen villages located near each of them respectively: Río Lagartos-western basin, Las Coloradas-middle basin and El Cuyo-eastern basin.
Environmental variables sampling: At each station, temperature, salinity, dissolved oxygen, and pH were measured in the water column at depths between 0.15 and 0.35 m using a multiparameter Aquaprobe AP-5000. A refractometer was used to measure salinity when the probe values exceeded 60 psu. Sediments were sampled with the same dredge or PVC core used for biological samplings and stored at -20 °C for grain size analysis. At the laboratory, sediments were dried at 60 °C to a constant weight, and later, the proportions of gravel (> 2 mm), sand (0.063-2 mm) and mud (< 0.063 mm) were obtained by sieving sediments with a sieve agitator for 20 minutes. The proportions of different sand size classes were measured with a CAMSIZER P4 at the Instituto de Ingeniería of the Universidad Nacional Autónoma de Mexico (UNAM). Sand size classes used were: very fine sands (0.063-0.125 mm), fine sands (0.125-0.250 mm), medium sands (0.250-0.500 mm), coarse sands (0.5-1 mm), and very coarse sands (1-2 mm). The percentage contribution of each sediment size class to total sediments was obtained. Sediments from the top three cm of the bottom were obtained for analysis of total organic carbon (TOC). TOC was analyzed using an elemental analyzer Costech 4010 at the Laboratorio de Análisis de Isótopos Estables of the Unidad de Química-Sisal, School of Chemistry (UNAM). Given the large amounts of carbonates in the sediments, these were first acidified with HCl 1M. Organic carbon percentage was obtained from total sediment weight.
The vegetation collected with each macrobenthos sample was separated from the sediments after through revision under the stereomicroscope to separate the macrobenthic organisms. The wet weight of the vegetation and the sediments were obtained after drying with blotting paper, and the gravimetric contribution of vegetation (algae and seagrasses) to the whole sample (i.e. vegetation and sediments) was calculated as a proxy of the amount of vegetation in the sample.
Data analysis: The environmental gradients along the lagoon were analyzed by simple linear regression between the abiotic factors and geographical longitude.
Abundance values of polychaetes were standardized to an area of 1 m2 and percentage contribution of each species to the total density at each station was calculated. Bray-Curtis similarity indices (Bray & Curtis, 1957) were calculated on the fourth-root transformed species density per station to define the relation of the faunal composition among sampling sites (Clarke & Warwick, 2001). A one-way distance-based Permutational Multivariate Analysis of Variance (PERMANOVA: Anderson, Gorley, & Clarke, 2008) under unrestricted permutation of the raw data was used to assess the potential effects of the two sampling gear (i.e. Ponar dredge and PVC core) on the community composition (9 999 permutations at maximum), with Monte Carlo sampling (Anderson & Robinson, 2003) employed to increase the number of permutations. After that, a two-factor PERMANOVA with the factor “station” (i.e. the station number) nested in the factor “basin” (three levels: western basin: Río Lagartos; middle basin: Las Coloradas; and eastern basin: El Cuyo) was used to test the significance of the differences in community composition between the basins defined a priori and between the stations located within each basin. A distance-based test for homogeneity of multivariate dispersions (PERMDISP, Anderson, Ellingsen & McArdle, 2006), was performed on the factor “basin”. Polychaete species groupings were evaluated by group-average linkage cluster analysis of the resemblance matrix. Samples relationships were also analyzed by non-metric multidimensional scaling (nMDS: Clarke & Warwick, 2001) of the resemblance matrix, producing a 2-dimensional ordination (Clarke & Warwick, 2001). A Similarity Percentage analysis (SIMPER) was performed to determine the species characterizing each basin within the lagoon system and to measure the contribution of each species to intra-group similarity or between-group dissimilarity (Clarke, 1993). Density, number of species (S), Shannon diversity (H’) and Simpson dominance index (λ) in the different basins and at each station were calculated, and the statistical significance of differences between basins were analyzed using a Mann-Whitney U test.
The relationships between assemblage structure or species’ distributions and environmental variables were explored by means of multivariate approaches. The environmental variables used were temperature, salinity, dissolved oxygen, pH, vegetation weight percentage from the sample, percentage contribution of each grain-size class (see above) to total sediments, and TOC %. Before the analyses, a Draftsman plot was performed on the environmental dataset in order to identify correlations among the variables. Only variables that were not correlated (Pearson’s correlation, ρ < 0.75) were used for the analyses. The BIOENV routine available in PRIMER6 (Clarke & Warwick, 2001) was used to identify the subset of environmental variables that best matched the density data of the species. Canonical correspondence analysis (CCA) (ter Braak, 1986) was performed on the density data for the most representative polychaete species (i.e. species appearing in more than one station or contributing to total abundance by more than 10 %) and the environmental variables identified by BIOENV to detect relationships between the species and those variables. Density data were square root-transformed prior to CCA calculations and then a permutation test (based on 1000 permutations) was performed to evaluate whether the set of environmental variables significantly explained the observed shifts in species densities.
All statistical analyses were performed using PRIMER 6 and PERMANOVA+ (Clarke & Warwick, 2001; Anderson et al., 2008), STATISTICA 10 (StatSoftInc.), and CANOCO software (ter Braak & Smilauer, 2002).
Results
Environmental characteristics: Salinity values ranged from 36.67 to 80, with a clear pattern of increase from the sea to the head of the lagoon (west to east with regression of salinity vs geographical longitude R 2 = 0.92). A pattern of eastward increase of pH was also observed (R 2 = 0.52), but spatial variability was small (7.6 to 8.62; SD = 0.22). The rest of the variables did not show clear spatial patterns of distribution (R 2 < 0.3). The values of the environmental variables at each station are shown in Appendix 1.
Polychaetes assemblages: A total of 827 specimens belonging to 26 species and 16 families were collected. Most of them were identified to species level except for 173 specimens (21 %) that belong to four species potentially new to science (Table 1). Polychaetes were collected in 10 stations out of the 16 stations sampled. They were not present at the eastern basin, and their eastern distribution in the lagoon system was at station 25 in the middle basin (Fig. 1, Appendix 1).
TABLE 1 Average density (ind m-2) and percentage contribution of species at each station. Total density, species richness, and Shannon and Simpson indices per station are shown (± standard deviation, SD). Empty samples (stations from 27 to 38) are not shown. (A) Western basin. (B) Middle basin
| A | Western basin | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 | 6 | 9 | 12 | 15 | |||||||
| Species | ind m-2 | % | ind m-2 | % | ind m-2 | % | ind m-2 | % | ind m-2 | % | |
| Armandia agilis (Andrews, 1891) | 1050.00 | 43.15 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Syllis prolifera Krohn, 1852 | 600.00 | 24.66 | 350.00 | 7.17 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Syllis lagunae Tovar-Hernández, Hernández-Alcántara & Solís-Weiss, 2008 | 50.00 | 2.05 | 1916.67 | 39.25 | 28.85 | 37.50 | 0 | 0 | 0 | 0 | |
| Sphaerosyllis taylori Perkins, 1981 | 183.33 | 7.53 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Chaetozone sp. | 0 | 0 | 66.67 | 1.37 | 19.23 | 25.00 | 0 | 0 | 0 | 0 | |
| Ceratonereis longicirrata Perkins, 1980 | 0 | 0 | 333.33 | 6.83 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Nereis pelagica Linnaeus, 1758 | 0 | 0 | 200 | 4.10 | 0 | 0 | 48.08 | 100.00 | 288.46 | 54.55 | |
| Ceratonereis sp. | 0 | 0 | 33.333 | 0.68 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Laoenereis sp. | 16.67 | 0.68 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Leitoscoloplos sp. | 83.33 | 3.42 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Naineris setosa (Verrill, 1900) | 0 | 0 | 1950.00 | 39.93 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Scoloplos (Leodamas) rubra (Webster, 1879) | 100.00 | 4.11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Lysidice sp. | 0 | 0 | 16.67 | 0.34 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Prionospio heterobranchia Moore, 1907 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Polydora quintanarroensis Delgado-Blas, 2008 | 150.00 | 6.16 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Scolelepis squamata (Müller, 1806) | 166.67 | 6.85 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Pectinaria meredithi Long, 1973 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Hypereteone heteropoda (Hartman, 1951) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Diopatra cuprea (Bosc, 1802) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Melinna maculata Webster, 1879 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 230.77 | 43.64 | |
| Schistomeringos perkinsi (Wolf, 1986) | 0 | 0 | 0 | 0 | 9.62 | 12.50 | 0 | 0 | 0 | 0 | |
| Filograna implexa Berkeley, 1835 | 0 | 0 | 16.67 | 0.34 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Branchiomma bairdi (McIntosh, 1885) | 0 | 0 | 0 | 0 | 9.62 | 12.50 | 0 | 0 | 9.62 | 1.82 | |
| Sabellidae sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Capitella sp. | 33.33 | 1.37 | 0 | 0 | 9.62 | 12.50 | 0 | 0 | 0 | 0 | |
| Hesione sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Total density (ind m-2) (± SD) | 2 433.33 (± 518.55) | 4 883.33 (± 1249.22) | 76.92 (± 27.20) | 48.08 (± 13.60) | 528.85 (± 149.58) | ||||||
| Species richness (S) | 7 (± 0) | 7.5 (± 2.211) | 3 (± 1.414) | 1 (± 0) | 2 (± 1.141) | ||||||
| Shannon diversity index (H’) | 1.411 (± 0.089) | 1.36 (± 0.137) | 0.984 (± 0.491) | 0 | 0.181 (± 0.256) | ||||||
| Simpson index (ʎ) | 0.324 (± 0.007) | 0.321 (± 0.058) | 0.417 (± 0.194) | 0 | 0.915 (± 0.119) | ||||||
| B | Middle basin | ||||||||||
| 20 | 21 | 22 | 23 | 25 | |||||||
| Species | ind m-2 | % | ind m-2 | % | ind m-2 | % | ind m-2 | % | ind m-2 | % | |
| Armandia agilis (Andrews, 1891) | 9.62 | 9.09 | 19.23 | 8.00 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Syllis prolifera Krohn, 1852 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Syllis lagunae Tovar-Hernández, Hernández-Alcántara & Solís-Weiss, 2008 | 0 | 0 | 0 | 0 | 19.23 | 1.06 | 0 | 0 | 9.62 | 1.59 | |
| Sphaerosyllis taylori Perkins, 1981 | 48.08 | 45.46 | 96.15 | 40.00 | 76.92 | 4.26 | 221.15 | 69.70 | 0 | 0 | |
| Chaetozone sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Ceratonereis longicirrata Perkins, 1980 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Nereis pelagica Linnaeus, 1758 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 115.39 | 19.05 | |
| Ceratonereis sp. | 0 | 0 | 0 | 0 | 153.85 | 8.51 | 0 | 0 | 0 | 0 | |
| Laoenereis sp. | 0 | 0 | 38.46 | 16.00 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Leitoscoloplos sp. | 0 | 0 | 19.23 | 8.00 | 1278.85 | 70.74 | 0 | 0 | 0 | 0 | |
| Naineris setosa (Verrill, 1900) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Scoloplos (Leodamas) rubra (Webster, 1879) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Lysidice sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Prionospio heterobranchia Moore, 1907 | 19.23 | 18.18 | 38.46 | 16.00 | 182.69 | 10.11 | 96.15 | 30.30 | 38.46 | 6.35 | |
| Polydora quintanarroensis Delgado-Blas, 2008 | 9.62 | 9.09 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Scolelepis squamata (Müller, 1806) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Pectinaria meredithi Long, 1973 | 9.62 | 9.09 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Hypereteone heteropoda (Hartman, 1951) | 0 | 0 | 9.62 | 4.00 | 96.15 | 5.32 | 0 | 0 | 0 | 0 | |
| Diopatra cuprea (Bosc, 1802) | 9.62 | 9.09 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Melinna maculata Webster, 1879 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Schistomeringos perkinsi (Wolf, 1986) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Filograna implexa Berkeley, 1835 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Branchiomma bairdi (McIntosh, 1885) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Sabellidae sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 442.31 | 73.02 | |
| Capitella sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Hesione sp. | 0 | 0 | 19.23 | 8.00 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Total density (ind m-2) (± SD) | 105.77 (± 95.19) | 240.39 (± 231.17) | 1 807.69 (± 1033.46) | 317.31 (± 258.37) | 605.77 (± 856.69) | ||||||
| Species richness (S) | 3 (± 2.828) | 4 (± 2.828) | 5 (± 1.414) | 1.5 (± 0.707) | 2 (± 2.828) | ||||||
| Shannon diversity index (H’) | 0.651 (± 0.921) | 1.024 (± 0.653) | 0.836 (± 0.454) | 0.178 (± 0.252) | 0.393 (± 0.555) | ||||||
| Simpson index (ʎ) | 0.679 (± 0.453) | 0.453 (± 0.235 | 0.597 (± 0.211) | 0.897 (± 0.144) | 0.5737 (± 0) | ||||||
The one-way PERMANOVA performed to test the influence of sampling gear on the community composition revealed non-significant effect (p > 0.05). Therefore, the following analyses were performed using data from both gear. Empty samples easterly from station 25, where polychaetes were not present, were discarded from the further analyses. The nMDs (Appendix 2) performed on the density data showed grouping of the polychaete assemblages according to their location inside each basin. Samples from the middle basin were mostly at the left of the plot and those from the western basin, at the right side. No clear pattern in samples distribution was identifiable in the vertical axis, related with a high variance among samples from the western basin (Appendix 2). Cluster analysis revealed that the polychaete assemblage at station 3, the station nearest to the mouth of the system, was similar to those found in the middle basin, whilst the one found at station 25, which was the easternmost site where polychaetes were collected, grouped with stations from the western basin (Appendix 2). The two-factor PERMANOVA proved significant differences in polychaete assemblages between basins (pseudo-F 1,19 = 7.17; Monte Carlo p < 0.01) and between the stations located within each basin (pseudo-F 8,19 = 2.70; Monte Carlo p < 0.001). Pairwise comparisons between stations revealed significant differences between some stations in the western basin (Table 3) and homogenous polychaete assemblages in the middle basin (Monte Carlo p > 0.05 in all cases). PERMDISP revealed non-significant differences in multivariate dispersion between basins (t = 2.190; p = 0.06), even if values were higher in the western basin (48.32) when compared to the middle basin (39.04). SIMPER analysis revealed high dissimilarity between both basins in terms of species composition (average dissimilarity = 90.00 %) (Table 2). Yet, intragroup similarities at the western (14.96 %) and middle (35.32 %) basins were low, suggesting high heterogeneity in species composition among stations within each basin. At the western basin, the polychaete assemblage was mainly defined by the occurrence of Nereis pelagica Linnaeus 1758 (48.98 %) and Syllis lagunaeTovar-Hernández, Hernández-Alcántara & Solís-Weiss, 2008 (19.04 %), while at the middle basin, Prionospio heterobranchia Moore, 1907 (51.80 %) and Sphaerosyllis taylori Perkins, 1981 (35.83 %) were the characteristic species. These four species were also the ones that mainly contributed to dissimilarity between the two basins (38.78 %), P. heterobranchia being absent from the western one.
TABLE 2 SIMPER analysis of the macrobenthic polychaetes to show (A) average similarities and species contributions within each basin and (B) average dissimilarities and species contributions between basins. Cut-off at cumulative contributions of 60%
| A | Average abundance | Average similarity | Contribution (%) | Cumulative contribution (%) | |
|---|---|---|---|---|---|
| Río Lagartos (western basin) | |||||
| Average similarity: 14.96 | |||||
| Nereis pelagica | 2.10 | 7.33 | 48.98 | 48.98 | |
| Syllis lagunae | 2.32 | 2.85 | 19.04 | 68.02 | |
| Las Coloradas (middle basin) | |||||
| Average similarity: 35.32 | |||||
| Prionospio heterobranchia | 2.78 | 18.3 | 51.8 | 51.8 | |
| Sphaerosyllis taylori | 2.52 | 12.66 | 35.83 | 87.63 | |
| B | Average abundance | Average abundance | Average dissimilarity | Contribution (%) | Cumulative contribution (%) |
| Average dissimilarity = 90.00 | Western basin | Middle basin | |||
| Prionospio heterobranchia | 0.00 | 2.78 | 10.90 | 12.12 | 12.12 |
| Sphaerosyllis taylori | 0.74 | 2.52 | 9.61 | 10.68 | 22.80 |
| Nereis pelagica | 2.10 | 0.66 | 8.23 | 9.15 | 31.94 |
| Syllis lagunae | 2.32 | 0.77 | 6.16 | 6.84 | 38.78 |
| Leitoscoloplos sp. | 0.60 | 1.61 | 5.31 | 5.90 | 44.68 |
| Armandia agilis | 1.14 | 0.77 | 4.61 | 5.13 | 49.81 |
| Syllis prolifera | 1.85 | 0.00 | 3.86 | 4.29 | 54.10 |
| Sabellidae sp.1 | 0.00 | 0.92 | 3.70 | 4.12 | 58.22 |
| Melina maculata | 0.78 | 0.00 | 3.47 | 3.85 | 62.07 |
Table 3 PERMANOVA pairwise tests comparing the community composition between stations in the western basin based on 9999 permutations (* p < 0.05, ** p < 0.01, ns p > 0.05)
| Stations | 3 | 6 | 9 | 12 |
| 3 | ||||
| 6 | 3.552* | |||
| 9 | 2.155ns | 2.403ns | ||
| 12 | 4.065* | 8.668** | 2.414ns | |
| 15 | 2.537ns | 2.959* | 1.708ns | 1.912ns |
Density values were overall higher in the western basin (mean = 1594.10 ind. m-2, SD = 2015.30 ind. m-2) than in the middle basin (mean = 615.38 ind m-2, SD = 799.51 ind m-2) (Fig. 2). High intra-basin variance was observed within each basin, especially in the western basin, where stations with both the highest (i.e. 4833.33 ind m-2 at station 6 and 2433.33 ind m-2 at station 3) and lowest (i.e. 48.08 ind m-2 at station 12, and 76.92 ind m-2 at station 9) density of organisms were located (Fig. 2; Table 1). Density in the middle basin ranged from 105.77 (station 20) to 1807.69 ind m-2 (station 22). The total number of species collected in the western basin (4.1, SD = 2.96) was higher than in the middle one (3.1, SD = 2.18). Again, the stations with both the highest (seven species at station 3) and the lowest (one species at station 12) species richness were located in the western basin (Table 1). At the middle basin, species richness varied from three (station 20) to five species (station 22).
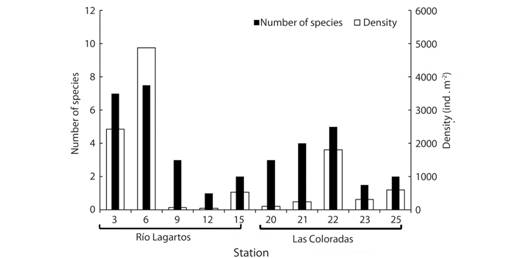
Fig. 2 Number of species and density of macrobenthic polychaetes at each sampling station in the lagoon. Basins are indicated. Empty samples (stations from 27 to 38) are not shown.
Overall, Shannon diversity (H’) was greater at the western (H’ = 0.787) than at the middle basin (H’ = 0.616), but among-stations high variability was also observed (Table 1). In the western basin, H’ was highest near the sea, at stations 3, 6, and 9, it was zero at station 12, and intermediate at station 15 (Table 1). At the middle basin, the highest H’ was observed to the west (stations 21 and 22) and H’ values were intermediate at the three easternmost stations. In general, the Río Lagartos system did not show a noticeable specific dominance (Simpson λ ranged from 0.32 to 0.91), and the patterns in λ were in general opposed to those of H’. Between-basin differences in the four community parameters were not statistically significant (Mann-Whitney U test p > 0.4 in all cases) likely owing to the high variance in all parameters within each basin.
Relationship with environmental variables: The Draftsman plot revealed that mud (< 0.63 µm) content was inversely correlated with gravel (> 2 mm) (ρ = -0.78) and coarse sand (0.5-1 µm) (ρ = -0.79) contents. Also, vegetation percentage contributions in the samples were positively correlated with fine sands’ (0.125-0.250 µm) content (ρ = 0.83).
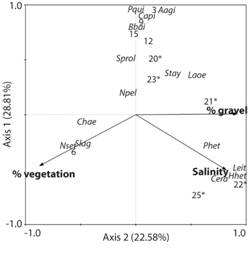
Fig. 3 CCA of macrobenthic samples based on density data for the dominant species of polychaetes. The first two axes are plotted together with explanatory variables. Station numbers are indicated: stations 3 to 15 are located within the western basin (Río Lagartos); stations 20 to 25 are located within middle basin (Las Coloradas; marked with an asterisk). Species codes: Aagi: Armandia agilis; Bbai: Branchiomma bairdi; Capi: Capitella sp.; Cera: Ceratonereis longicirrata; Chae: Chaetozone sp., Hhet: Hypereteone heteropoda; Laoe: Laoenereis sp.; Leit: Leitoscoloplos sp.; Npel: Nereis pelagica; Nset: Naineris setosa; Phet: Prionospio heterobranchia; Pqui: Polydora quintanarroensis; Slag: Syllis lagunae; Sprol: Syllis prolifera; Stay: Sphaerosyllis taylori.
The BIOENV routine (with only samples west from station 25 included) showed that the best correlation between the structure of the assemblages and the environmental variables was obtained with four variables (salinity, pH, and gravel and vegetation proportions in the samples (R 2 = 0.58; p = 0.001). Those variables were used to perform the CCA and significantly (p = 0.002) accounted for 67.0 % of its total variance, with 76.7 % of this variance explained by the first two CCA axes. Samples from the middle basin (stations 20 to 25) were distributed on the right side of axis 2 and those from the western basin (stations 3 to 15) were on the center-top or left side of the plot (Fig. 3). No clear pattern in samples distributions was observed for axis 1, associated to a high intra-group variance among samples located within the western basin. Axis 2 was highly correlated with gravel proportions (r 2 = 0.90), vegetation proportions (r 2 = 0.86), and salinity (r2 = 0.81). Axis 1 was moderately correlated with salinity (r 2 = -0.49) and vegetation proportions (r 2 = -0.45). Naineris setosa (Verrill, 1900), S. lagunae and Chaetozone sp. were associated to sites with high vegetation proportions in low salinity environments with low gravel content in the sediments. Instead, P. heterobranchia, Hypereteone heteropoda (Hartman, 1951), Leitoscoloplos sp. and Ceratonereis longicirrata Perkins, 1980 were associated to high salinity and gravel proportions in the sediments.
Discussion
The present study analysed the community structure of macrobenthic polychaetes in a coastal hypersaline lagoon with a wide range of salinity. It explored the potential effects of salinity together with other environmental variables. The main environmental drivers of assemblages’ distributions were salinity, vegetation proportions and sediment grain size. These variables have already been identified as drivers of the macrobenthic fauna distribution in Yucatán (Kuk-Dzul et al., 2012; Morelos-Villegas, Condal, & Ardisson, 2018) and worldwide (e.g. Moreira, Quintas, & Troncoso, 2006; Alves et al., 2013; Magalhães et al., 2019). Out of the 26 species reported in this study, 23 (except Ceratonereis longicirrata, Prionospio heterobranchia, Hypereteone heteropoda that were recorded by Kuk-Dzul et al. (2012)) are new records for the Río Lagartos lagoon system, highlighting the lack of knowledge of this group in the area.
Polychaetes had a very patchy distribution along the lagoon, which is a common trait for macrobenthos inhabiting such highly variable and geomorphologically complex coastal environments (e.g. Hernández-Alcántara & Solís-Weiss, 1991; Meerhoff et al., 2013; Magalhães & Bailey-Brock, 2014). The complex hydrodynamics and the strong environmental variability in Río Lagartos mainly result from the highly frictional nature of this very shallow (0.5 m depth in average) system, with an intricate morphology in a large area. Its geometric configuration restricts the communication between the inner basins, where an excess evaporation increases the salinity, and a whole set of biogeochemical processes are constantly reshaping the physicochemical characteristics and the distribution of sediments and organic matter at multiple spatial scales (Marin-Coria & Enríquez, 2016; Enríquez, Reyes, Papiol, & Aragón, 2017; Chiappa-Carrara et al., 2017). Small-scale environmental heterogeneity may in turn affect polychaetes distribution, as reflected in the high variability in species composition, density and community parameters observed among stations. Yet, some spatial patterns of distribution of polychaetes associated to the three estuarine basins of the Río Lagartos lagoon system were detected, which is consistent with the previous environmental and biological large-scale spatial zonation described for the lagoon (Ortegón-Aznar et al., 2001; Valdes & Real, 2004; Vega-Cendejas & Hernández de Santillana, 2004; Peralta-Meixueiro & Vega-Cendejas, 2011). Differences in polychaetes communities between basins are attributable to strong environmental gradients between them. Those are favoured by the physical restricted connection between the different basins by sills, and by the particular hydrological characteristics of each basin (Herrera-Silveira, Ramirez, & Zaldivar, 1998).
The most remarkable spatial gradient of the community of polychaetes was the sudden disappearance of polychaete fauna at the eastern side, which corresponds to the basin located furthest from the sea that had the highest salinity and pH values. This eastern system is inundated by seawater, but with limited connection to the ocean dynamics, and it is subject to excess of evaporation and long residence time of seawater, which increasingly promotes salinity concentration towards the head (Enríquez, Reyes et al., 2017). Polychaetes are largely represented and abundant in estuarine environments (e.g. Glasby & Tarmo, 2008; Hernández-Alcántara, Cortés-Solano, & Solís-Weiss, 2011; Carvalho et al., 2011) and a variety of species are adapted to variable salinity ranges (David & Williams, 2016), including hypersaline conditions (Por, 1972; Lardicci et al., 1993; Lamptey & Armah, 2008; Can et al., 2009; Marzano et al., 2010; Joydas et al., 2011; Joydas et al., 2015). The few available studies of macrobenthic communities in natural hypersaline lagoons with salinity values above 50 psu report the presence of low abundant species tolerant to high salinity at values higher than ca. 60 psu (Lamptey & Armah, 2008; Can et al., 2009; Marzano et al., 2010; Joydas et al., 2011; Joydas et al., 2015). In our study, polychaetes were found at salinity values up to 58.14 psu, but absent at the eastern basin, where salinity ranged from 67 to 80 psu. Although some adult polychaete species seem able to survive under salinity > 60 psu, impaired reproduction has been reported at salinity > 50 psu (Can et al., 2009), and deleterious effects on polychaete larvae were observed at salinity of 70 psu by Lyster (1965). Environmental seasonality may favour seasonal salinity decreases and associated peaks in reproduction and survival of larvae in other hypersaline lagoons (Can et al., 2009). However, salinity at the head of Río Lagartos lagoon is above 65 psu throughout the year (Peralta-Meixeiro & Vega-Cendejas, 2011), likely preventing the reproduction and establishment of polychaete populations in this area. The strong salinity gradient between Las Coloradas (middle) and El Cuyo (head) basins (Valdes & Real, 2004; this study), may represent an additional barrier for polychaetes dispersion. Variable or abrupt salinity gradients provide significant stress to bottom-dwelling aquatic organisms, and the effect is greatest for sessile macro-fauna (Telesh, Schubert, & Skarlato, 2013). Moreover, high salinity at El Cuyo has also been identified as a major cause of the absence of submerged vegetation (i.e. algae or seagrass) (Ortegón-Aznar et al., 2001). This may have impaired the ability of some taxa to colonize the basin due to the lack of this food source, shelter and reproductive habitat (Ibáñez-Aguirre & Solís-Weiss, 1986; Thrush, Hewitt, & Pridmore, 1989).
Inter-basin spatial differences were also reflected in the presence of distinct polychaete assemblages in the west (Río Lagartos) and middle (Las Coloradas) basins. Yet, density, species richness and diversity values were not statistically different between basins, likely because of a high heterogeneity between stations within each basin, especially in Río Lagartos (near the mouth). The entire Gulf of Mexico is micro tidal and the Yucatan coastal sea has a mixed tidal regime, with diurnal dominance and maximum amplitude of 40 cm during spring tides and semidiurnal tendency with variations of only a few cm (<10) during neap tides (Enriquez, Mariño-Tapia, Silva-Casarín, & Mendoza-Baldwin, 2011). Results of the ongoing CEMIE-Océano project (Enríquez, Chiappa-Carrara, Roldán, & Marín-Coria, 2017) have revealed that the small tidal range has limited capacity to force the lagoon recirculation and the modulation is rapidly diminished at the first long narrowing in the Las Coloradas strait, separating the western and middle basins. Therefore, these two basins have very different characteristics, and stronger environmental variability takes place in the western basin, resulting in heterogeneous distribution of the fauna. Pronounced patchiness can be deduced from (a) the high variability in species composition, density, species richness and diversity between the different stations within the basin and (b) the low intragroup similarity in species composition, as revealed from SIMPER analysis. High variations in the macrobenthic community parameters close to the lagoon mouth have already been reported (Kuk-Dzul et al., 2012) and in the study area coincide with pronounced changes in environmental characteristics (Valdes & Real, 2004) and with the presence of a high diversity of subaquatic vegetation (Ortegón-Aznar et al., 2001) that enhances small-scale spatial variability through the generation of microhabitats.
Polychaetes’ density values in the lagoon system were within the values reported for other lagoons in the Yucatan Peninsula (Pech, Ardisson, & Hernández-Guevara, 2007), with more extreme maximum and minimum values. Regarding species richness, Río Lagartos can be considered on the lowest range for coastal lagoons in the Gulf of Mexico (Hernández-Alcántara et al., 2011), although seasonal changes in community composition may take place (Pech et al., 2007) that would modify such values. The highest density and diversity were registered at the westernmost stations (i.e. stations 3 and 6), which are framed by the three marine inlets and featured environmental conditions most similar to marine ones (Ghinaglia, Herrera-Silveira, & Comín, 2004; Merino-Virgilio, Okolodkov, Aguilar-Trujillo, & Herrera-Silveira, 2013). Marine salinity, the potential exchange of water with the sea and the presence of considerable proportions of subaquatic vegetation that provide a broad spectrum of habitats (Giangrande, 1988; Hernández-Alcántara et al., 2011) may interact to explain such fauna patterns. The overall highest density and diversity values at station 6 are likely enhanced by the presence of dense algal mats of Avrainvillea sp., as already reported by Magalhaes & Bailey-Brock (2014) in Hawaii. Naineris setosa and Syllis lagunae contributed to 80 % of the density in station 6 (39.93 % and 39.24 %, respectively). Despite the wide distribution of N. setosa in subtropical and tropical waters in the Atlantic, the Gulf of Mexico, the Caribbean Sea and the eastern Pacific (see Blake & Giangrande, 2011 and references cited therein; Díaz-Díaz, Vanegas-Espinosa, Cárdenas-Oliva, & Liñero-Arana, 2012), its faculty to adapt to new environments (e.g., the Mediterranean Sea, where it is an invasive species: Blake & Giangrande, 2011; Belal & Ghobashy, 2012; Khedhri, Lavesque, Bonifácio, Djabou, & Afli, 2014; Atzori, López, Addis, Sabatini, & Cabiddu, 2016), and its capacity to live in different substrata (e.g., Hernández-Alcántara & Solís-Weiss, 1995; Díaz-Díaz, Bone, & López-Ordáz, 2014), in Río Lagartos it was only found at station 6, suggesting that factors additional to substrate type may have prevented its wider distribution. Syllidae was the dominant family in terms of abundance in Río Lagartos, a common pattern in estuarine systems (Omena & Creed, 2004; Hernández-Alcántara et al., 2011), especially in those with carbonate sediments (Granados-Barba, Solís-Weiss, Tovar-Hernández, & Ochoa-Rivera, 2003; Tovar-Hernández, Hernández-Alcántara, & Solís-Weiss, 2008). Together with the family Nereididae, they were also the most widely distributed family in the lagoon. A substitution of dominant syllid species between basins was observed: S. lagunae and Syllis prolifera Krohn, 1852 dominated in the western basin, and S. taylori, was more important at the middle basin. This latter species is capable of living in very different substrata (Ruiz-Ramírez & Salazar-Vallejo, 2001; Álvarez & San Martin, 2009; Surugiu & San Martin, 2017), which may favor its proliferation in unvegetated areas. Nereids comprised the highest numbers of species (i.e. four species) in the lagoon, consistent with their high diversity in coastal lagoons and estuaries from the Gulf of Mexico (Hernández-Alcántara et al., 2011).
Despite the heterogeneous distribution of the sediments, coarser sediments were more abundant in the middle basin while muddy bottoms were more important to the west. The importance of sediment grain size as a major driver of polychaetes distribution in estuarine systems is well documented. Sediment grain size can indirectly affect polychaetes distribution through the capacity of organic matter retention and oxygenation hence, different trophic groups are associated to particular sediment-size compositions (Carvalho et al., 2011). In well-vegetated areas where soft bottoms prevail, such as stations 6, 12 and 15 in the western basin, macroalgal decay and high organic matter retention in the sediments could have been thought to favor deposit feeders (Lamptey & Armah, 2008; Carvalho et al., 2011), but similar contributions of deposit feeders, such as N. setosa or Melina maculata Webster, 1879, and carnivores, such as syllids, nereids and orbiinids, were found. Greater contributions of carnivore species might be expected under the presence of coarser sediment and low vegetation (i.e. in the middle basin), but that pattern was not clear either. Moreover, the deposit feeder P. heterobranchia, dominant in the middle basin, had previously been reported from high silt and clay content bottoms in this lagoon (Kuk-Dzul et al., 2012). Yet, the high environmental variability together with the particular lifestyle of the species may have provided a particular space in Las Coloradas. Its tube-forming behavior may contribute to its distribution in barely-vegetated substrates (Liñero-Arana & Diaz-Diaz, 2005). Such behavior may also enhance salinity tolerance through saline-stress prevention (Brusca & Brusca, 2003).
The present study expanded the salinity distribution ranges for several species previously recorded in the Yucatan Peninsula, and those can be now considered euryhaline, and able to live in hypersaline conditions (e.g. S. lagunae previous records at 18.4 to 36 psu (Tovar-Hernández et al., 2008), this study at 36.6 to 58.14 psu; P. heterobranchia: previous records at 17.2 to 35.4 psu (Ibañez-Aguirre & Solis-Weiss, 1986), this study at 44.10 to 58.14 psu; N. pelagica: no previous records available, this study at 38.2 to 58.1 psu). Intraspecific geographical variations in environmental tolerance and physiological plasticity have been described among both vertebrates and invertebrates for temperature, and the scope of physiological plasticity is thought to be proportional to the magnitude of environmental variation that a species experiences in its native habitat (Barria, Bacigalupe, Lagos, & Lardies, 2018). It is therefore likely that the unstable saline conditions of this system favored environmental adaptation of some species, which represents an adaptive advantage for distribution expansion. Such may be the case of P. heterobranchia: its euryhaline character together with its capacity of living in different substrates (Ibáñez-Aguirre & Solís-Weiss, 1986; Salazar-Vallejo, 1990; Omena & Creed, 2004; Hernández-Alcántara et al., 2014) may explain its dominance in the middle basin, and its wide distribution in Mexico (Hernández-Alcántara, González-Ortíz, & Solís-Weiss, 1994; Díaz-Castañeda, de León-González, & Solana-Arellano, 2005). Adaptation to high salinity may also favor species survival, should predicted salinity increase associated to climate change occur (Curry, Dickson, & Yashayaev, 2003; Boyer, Levitus, Antonov, Locarnini, & Garcia, 2005), especially in this type of lagoons where limited water exchange and prevailing evaporation enhance salinity increases.
The Río Lagartos lagoon system harbors a highly heterogeneous polychaete community, especially near the lagoon mouth, likely associated to the strong variability of environmental characteristics. Strong gradients in environmental abiotic (salinity and sediment grain size) and biotic (abundance of subaquatic vegetation) characteristics between basins defined strong changes in species composition. Yet, the particular effect of each environmental factor cannot be singled out, and it is the interaction among them and with the particular lifestyle and biology of each species that define the distribution of each population.
Polychaetes were absent of environments with salinity > 60 psu but were well represented in hypersaline (salinity > 40 psu) areas. Many of the species in such areas had only been reported from estuarine or littoral areas, and this study contributed to establish their tolerance to wider salinity ranges. Despite the ecological importance and the extremely hypersaline character of the Río Lagartos lagoon system, there is very little knowledge on its macrofauna composition and ecology, and near 90 % of the species reported in this study are first records.
Ethical statement: authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.

