











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Los eventos geológicos y climáticos que han ocurrido a través del tiempo, han provocado que en los Andes tropicales exista una gran cantidad de hábitats terrestres. Estos ocasionan que la composición y estructura de la vegetación sean altitudinalmente variables, favoreciendo la alta heterogeneidad y diversidad de especies, que con el tiempo se han tornado propias en cada lugar (Churchill, Balslev, Forero, & Luteyn, 1995; Webster, 1995; van der Hammen, 2000; van der Hammen & Hooghiemstra, 2001; van der Hammen & Otero-García, 2007).
Colombia por su posición geográfica ecuatorial en Sudamérica, es uno de los países con la mayor diversidad biótica del mundo, posee tres cordilleras, que alojan una gran cantidad de áreas ocupadas por ecosistemas naturales que mantienen una enorme riqueza florística (Bernal, Gradstein, & Celis, 2016). Sin embargo, las acciones antrópicas como la expansión de la frontera agrícola, ganadera, quemas y erosión de suelos, han devastado estas áreas (Gentry, 1991; 1995; van der Hammen & Rangel-Ch., 1997; Serrano, 2007).
Los páramos son ecosistemas que se reconocen por su alta diversidad (Smith & Cleef, 1988; Sklenár, Hedberg & Cleef, 2014), comparados con otros ecosistemas similares en el mundo. Son propios del Neotrópico y se ubican en la parte alta de las montañas entre la franja de bosque andino y la parte más baja de las nieves (Rangel-Ch, 2000; Hofstede, Segarra, & Mena, 2003; Llambí & Cuesta, 2014). Ocupan el 2.5 % del territorio colombiano (Sarmiento, Cadena, Sarmiento, Zapata, & León, 2013) y brindan servicios ecosistémicos, como la regulación y constante provisión de agua (Rangel-Ch., 2000), albergan un alto número de especies nativas y endémicas (Hofstede et al., 2003).
Actualmente en Colombia la delimitación de la franja de transición o ecotono del bosque y páramo, ha dado origen a una extensa discusión, debido a que se han instaurado leyes (ley 99/1993) que permiten la protección y conservación de los páramos, pero no se tiene una noción clara de la delimitación de dicha franja (Llambí, 2015). La franja bosque-páramo es definida por Bader, Rietkerk, & Bregt (2007a), como una zona de transición, donde confluyen especies del bosque y la vegetación achaparrada de páramo, tal franja puede variar, debido a la alta heterogeneidad espacial de las cordilleras y los Andes en general (Bader van Geloof & Rietkerk, 2007b; Ataroff & García-Núñez, 2013). Por otra parte, van der Hammen (1998), Ramírez, Llambí, Schwarzkopf, Gámez y Márquez (2009) y Arzac, Chacón-Moreno, Llambí, y Dulhoste (2011), sostienen que esta zona se caracteriza por tener una gran variabilidad en composición florística. A su vez, Harsch & Bader (2011) definen para esta franja cuatro formas o patrones espaciales (difuso, abrupto, isla o lengüetas y “krummholz” en alemán madera doblada) basados en mecanismos de tres niveles, primero (limitación en el crecimiento, pérdida de biomasa y mortalidad de plántulas), segundo (estrés: fotoinhibición, daño por congelamiento, vientos, otros) y tercero (interacción con sus vecinos). Lo anterior, está relacionado con la dinámica de cada franja, debido a que integran factores abióticos (topografía, suelos, altitud, latitud, vientos, entre otros) y bióticos (composición de especies, respuestas fisiológicas, ente otros) (Rangel-Ch., 2000; Vargas & Pedraza, 2004; Sexton, McIntyre, Angert, & Rice, 2009). Sin embargo, llegar a un acuerdo en la delimitación de la franja no ha sido posible. Este trabajo busca conocer desde el componente florístico (composición y estructura) una aproximación a la franja de transición entre el bosque y el páramo en el flanco occidental del complejo de páramos Guantiva-La Rusia en los departamentos de Boyacá y Santander.
MATERIALES Y MÉTODOS
Área de estudio: El complejo Guantiva-La Rusia (GL) es un corredor de páramos, bosque andino y alto andino, ubicado en el flanco occidental de la cordillera Oriental de los Andes colombianos). El estudio se realizó en los municipios de Duitama, Belén (páramo de La Rusia) y Onzaga-Susacón (páramo de Güina), en los departamentos de Boyacá y Santander. En Duitama (5°56'20.4” - 5°57'1.4” N & 73°5'44.3” - 73°6'9.6” W), el páramo es el ecosistema más estudiado del complejo, y presenta alta diversidad florística (Morales et al., 2007; Martínez & Díaz, 2017), posee una extensión de 10 381 ha y presenta afectaciones humanas (Mora-Osejo & Sturm, 1994); Belén (6°1'33.8” - 6°2'50.4” N & 72°57'57” - 73°1'2.6” W), se encuentra al nor-oriente de Boyacá, con una extensión de páramo de 6 080 ha; Onzaga-Susacón (6°12'45.6” - 6°13'9.4” N & 72°46'42.2” - 72°47'7.7” W) al sur de Santander y norte de Boyacá, con un área de páramo de 13 033 ha (Fig. 1).
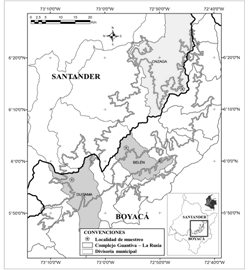
Fig. 1. Fig 1 Área de estudio, delimitación del complejo de páramos Guantiva-La Rusia. Study area, delimitation of the paramo Guantiva-La Rusia complex.
Muestreo: En cada localidad se realizó un transecto altitudinal con cuatro (Duitama, Onzaga-Susacon) y cinco (Belén) estaciones de muestreo cada 100 m (desde el bosque hasta el herbazal con presencia de frailejones). En cada estación se ubicaron tres parcelas; para evaluar el bosque fueron instaladas parcelas de 4 x 25 m [100 m2, divididas en subparcelas de 4 x 5 m (20 m2)], para arbustal y herbazal parcelas de 4 x 12.5 m [50 m2, divididas en subparcelas de 4 x 2.5 m (10 m2)] (Marín, 2013). Se evaluaron las siguientes variables: altura (m), diámetro a la altura del pecho (DAP), cobertura (Rangel-Ch. & Velásquez, 1997), abundancia y riqueza. Para las herbáceas se midió por datos porcentuales y de frecuencia en cada parcela. El material fue depositado en los Herbarios UPTC y FMB (Thiers, 2018), bajo la clasificación APG IV (2016).
Análisis de datos: Se determinó la riqueza y composición florística con base en el número de familias, géneros y especies, en cada uno de los sitios de muestreo. Seguidamente en cada estación de muestreo, se determinaron las formaciones vegetales, a partir de la cobertura relativa de los hábitos de crecimiento: árbol, arbolito, arbustillo, arbusto, hierba, roseta, macolla, entre otros (Marín, 2013). Se calculó el índice de diversidad alfa según Moreno, Barragán, Pineda, y Pavón (2011), que mide la diversidad de una comunidad integrada pori especies con igual número de individuos [p1 = p2 = p3 = ps], mediante el número efectivo de especies a nivel de formaciones vegetales dentro de cada localidad, con el uso de la diversidad de orden 1 (q1). Para establecer las diferencias en la composición de la vegetación, dentro y entre las localidades se organizó la información teniendo en cuenta la presencia-ausencia de especies por estación, y se usó para llevar a cabo el análisis de diversidad beta, a través del paquete R Betapart (Baselga, 2012), con las funciones “beta-multi” la cual permite obtener el recambio ((sim - disimilaridad de Simpson) y anidamiento ((sne - resultante de la fracción de disimilaridad de Sorensen). Además, se empleó el “beta-pairwise” que incluye las funciones para calcular matrices de distancia, con el uso de diferencias por pares ((sor) para hacer un análisis de conglomerados. Adicionalmente, se calculó el índice de Whittaker (1972) con el fin de conocer las comunidades o agrupamientos entre estaciones.
RESULTADOS
Riqueza: Se registraron 70 familias, 157 géneros y 302 especies (Apéndice). La familia Asteraceae es la más diversa con 16 géneros y 39 especies, seguida de Ericaceae (11/19), Poaceae (11/12), Orchidaceae (10/14) y Melastomataceae (7/20). El género más diverso es Miconia con 14 especies, seguido de Elaphoglossum (9), Hypericum (7), Ageratina y Pentacalia con seis cada una. La riqueza de familias y géneros por localidad no presentó mayores diferencias. La localidad con mayor número de especies fue Belén con 176, que representan el 58 % del total (Tabla 1).
TABLA 1: TABLE 1: Coberturas vegetales, riqueza florística, rango altitudinal por estación y diversidad verdadera alfa por cobertura en cada localidad, Guantiva-La Rusia Plant coverings, floristic richness, altitudinal range by season and true alpha diversity by coverage in each location, Guantiva-La Russia complex
| Localidad | Estación (cobertura vegetal) | Rango altitudinal (m) | No. Familias | No. Géneros | No. Especies | Diversidad verdadera alfa por cobertura vegetal |
| - | - | - | - | - | - | (q1) |
| Duitama | DE1 (Bosque) | 3 265 - 3 286 | 34 | 48 | 58 | 34.01 |
| Duitama | DE2 (Arbustal) | 3 394 - 3 434 | 34 | 51 | 61 | 29.09 |
| Duitama | DE3 (Arbustal) | 3 490 - 3 516 | 23 | 44 | 48 | 29.09 |
| Duitama | DE4 (Herbazal) | 3 593 - 3 615 | 20 | 31 | 38 | 13.57 |
| Duitama | Total | --- | 56 | 105 | 147 | 44.17 |
| Belén | BE1 (Bosque) | 3 275 - 3 336 | 29 | 38 | 48 | 23.56 |
| Belén | BE2 (Arbustal) | 3 411 - 3 430 | 19 | 31 | 35 | 41.21 |
| Belén | BE3 (Arbustal) | 3 499 - 3 505 | 28 | 48 | 59 | 41.21 |
| Belén | BE4 (Arbustal) | 3 642 - 3 649 | 34 | 56 | 68 | 41.21 |
| Belén | BE5 (Herbazal) | 3 766 - 3 776 | 19 | 30 | 39 | 12.63 |
| Belén | Total | --- | 54 | 108 | 176 | 66.92 |
| Onzaga- Susacón | OE1 (Bosque) | 2 944 - 3 129 | 35 | 48 | 55 | 45.49 |
| Onzaga- Susacón | OE2 (Bosque) | 3 191 - 3 200 | 37 | 50 | 60 | 45.49 |
| Onzaga- Susacón | OE3 (Arbustal) | 3 300 - 3 313 | 27 | 46 | 49 | 25.72 |
| Onzaga- Susacón | OE4 (Herbazal) | 3 444 - 3 456 | 14 | 25 | 27 | 11.66 |
| Onzaga- Susacón | Total | --- | 55 | 102 | 141 | 68.00 |
| Total, en las tres localidades | - | - | 70 | 157 | 302 | - |
Formación vegetal: Se determinó la estructura y tipo de formación vegetal, así: en Duitama, la estación uno (E1) se definió como bosque, por el predominio de árboles con 88.71 %, seguido de arbolitos (5.24 %). La estación dos (E2) es un arbustal con el 70.20 % de arbustos, seguido de arbustillos (16.05 %). La estación tres (E3) es un arbustal, con predominio de arbustillos y arbustos (46.03 y 42.82 % respectivamente). La estación cuatro (E4) es un herbazal con predominio de macollas (32.30 %) seguidas de arbustillos (23.46 %).
En Belén los hábitos dominantes en la estación uno (E1), los árboles con el 90.83%, seguida de arbustos con 3.44 %, y se definió como bosque. La estación dos (E2) es un arbustal con predominio de arbustillos y arbustos con el 38.06 y 37 % respectivamente. La estación tres (E3) es un arbustal con arbustos (78.85 %) y arbustillos (6.44 %). La estación cuatro (E4) es un arbustal con arbustos (42.24 %) y arbolitos (26.80 %), y la estación cinco (E5) es un herbazal con macollas (71.69 %) seguida de, rosetas con un solo tronco (9.78 %).
En Onzaga-Susacón, las estaciones uno y dos corresponden a bosque por sus hábitos arbóreos (E1-71.61, E2-74.05 %) y arbustivos (E1-14.67, E2-13.56 %). La estación tres (E3) es un arbustal con arbustos y arbustillos (67.91 y 9.91 % respectivamente), y la estación cuatro (E4) es un herbazal donde las macollas representan el 71.56 %, seguidas del hábito chusqueoide (7.11 %) (Tabla 1).
Según las formas propuestas por Harsch y Bader (2011), se registran para Duitama y Onzaga-Susacón, una franja de transición con forma difusa y para Belén, en islas o lengüetas. En los herbazales se destacan especies de los géneros Achyrocline, Baccharis, Espeletia, Chusquea, Hypericum, Hypochaeris, Oreobolus y Halenia, en los arbustales, Vaccinium, Gaultheria, Hypericum, Ageratina, Berberis, Bejaria, Dendrophthora, Miconia y Peperomia y en los bosques Brunellia, Clusia, Anthurium, Miconia, Quercus, Solanum, Clethra y Weinmannia; adicionalmente, se registraron 35 especies propias de los herbazales que corresponden al 42.6 % del total registrado, 93 (54.7 %) para los arbustales y 96 (73.8 %) para los bosques, junto con 41 especies compartidas entre las formaciones vegetales de bosque y arbustal, entre arbustal y herbazal se comparten 30 especies y para las tres formaciones vegetales se hallaron seis especies compartidas.
Diversidad: la mayor diversidad se registró para el transecto de Onzaga-Susacón con 68 especies efectivas, seguido de Belén (66.92) y Duitama (44.17) (Tabla 1). El bosque de Onzaga-Susacón registra 1.9 y 1.3 veces, más diversidad que Belén y Duitama respectivamente. Sin embargo, esta localidad registra el valor más bajo para el arbustal con 25.72 especies efectivas. El herbazal más diverso corresponde a Duitama con 1.07 y 1.16 veces más diverso que Belén y Onzaga-Susacón (Fig. 2).
Diversidad beta: este análisis mostro que el recambio espacial de especies ((sim = 0.85) es alto entre las estaciones evaluadas; mientras que el anidamiento fue bajo ((sne = 0.09). Se registran cinco agrupamientos que corresponden: a) estaciones de bosque, que se subdividen en dos comunidades (BE1 - DE1 y OE1 - OE2); b) estaciones de herbazales (BE5 y DE4) con una disimilitud del 67 %; c) la estación arbustal de Onzaga-Susacón (OE3) con el 70 % de disimilitud; d) una estación de arbustal de Belén (BE2) y una estación de herbazal de Onzaga-Susacón (OE4) y e) estaciones de arbustal de Duitama y Belén (BE4, DE2, BE3 y DE3. El índice de Whittaker registra cinco comunidades (5.0667), las cuales coinciden con las mencionadas anteriormente (Fig. 2).

Fig. 2. Fig. 2 Agrupamiento a partir del análisis de diversidad beta entre las estaciones (E) de Duitama (D), Belén (B) y Onzaga-Susacón (O), en el costado occidental del complejo de páramos Guantiva-La Rusia (Boyacá-Santander). Grouping based on the analysis of beta diversity between the stations (E) of Duitama (D), Belén (B) and Onzaga-Susacón (O), on the western side of the páramos Guantiva-La Rusia complex (Boyacá-Santander).
DISCUSIÓN
Las familias con mayor riqueza en géneros y especies aquí registradas, son similares a otros estudios realizados en los ecosistemas de alta montaña (Rangel-Ch., 2000; Briceño & Morillo, 2006; Lozano, Cleef, & Bussmann, 2009; Cuello, Cleef, & Aymard, 2010). A lo largo del flanco occidental se encontró un alto grado de semejanzas en la composición de familias (77 - 80 %), géneros (64 - 69 %), y menor proporción de especies (46 - 58 %).
En las localidades de Duitama y Onzaga-Susacón, la mayor diversidad de especies se encontró en las estaciones de bosque, lo que concuerda con estudios realizados por Cuatrecasas (1958), Gentry (1995), Galindo, Betancur, y Cadena (2003), quienes afirman que la diversidad vegetal es mayor en los transectos de altitudes bajas y es menor en las zonas altas. Pese a esto, los bosques entre sí, presentaron diferencias notables, lo cual indica que, aunque son formaciones vegetales estructuralmente similares poseen riqueza variable entre ellos (Galindo et al., 2003).
La alta diversidad del arbustal en Belén, se explica por los efectos de transformación de las formaciones vegetales originales, como consecuencia de la tala, quema, ganadería y cultivos (papa), que históricamente se han visto en la zona e igualmente han sido registrados por otros autores en los Andes (Serrano, 2007; Velasco-Linares & Vargas, 2008; Sarmiento & Llambí, 2011). Tales eventos, junto con la inclinación del terreno han favorecido la llegada de especies arbustivas que soportan las nuevas condiciones de las áreas transformadas. Esta modificación ha ocasionado en esta localidad que dicha franja tenga un rango altitudinal (300 m) mayor a las demás, lo que se ve reflejado en un mayor muestreo (tres estaciones) y pueda ser la causa de presentar una mayor diversidad. Lo anterior, se argumenta con la teoría del disturbio intermedio de Connell (1978), que indica, que una alta diversidad de especies es generada luego de un disturbio antrópico.
Las estaciones con dominancia de estratos arbóreos fueron: uno de Duitama (3 265 - 3 286 m), Uno de Belén (3 275 - 3 336 m), y uno - dos de Onzaga-Susacón (2 944 - 3 200 m) que corresponde a bosque altoandino, observaciones similares fueron realizadas por Acosta-Solis (1984), Bader et al., (2007a), van der Hammen (1998), Ramírez et al. (2009) y Arzac et al. (2011), quienes indican que estos bosques en su límite superior están dominados por árboles y arbustos. Por otro lado, Rangel-Ch. (2000) en sus estudios menciona que los géneros Weinmannia y Clethra son dominantes en los bosques andinos, lo cual coincide con lo encontrado en este trabajo (Weinmannia tomentosa, W. fagaroides y Clethra fimbriata), junto con Quercus humboldtii los más dominantes.
La amplitud de la franja de transición bosque-páramo en el flanco, no es uniforme; se encontró que oscila entre 3 300 y 3 649 m, contrastado con lo mencionado por autores como, Ataroff & Sarmiento (2004), Rivera & Rodríguez (2011) y Ataroff & García-Núñez (2013), quienes definen dicha franja entre el rango de los 2 700 y 3 000 m; lo anterior, se debe a factores de tipo biótico y abiótico que afectan la amplitud de los límites de cada localidad.
Monasterio (1980) señala que en la franja de transición bosque-páramo se distribuyen especies arbóreas que conforman islas de bosques, las cuales según Llambí (2015) están dominadas por familias como Myrsinaceae, Rosaceae, Clusiaceae y Asteraceae; no obstante, en el presente estudio se encontraron las mismas familias, integrando estas islas, y se hallaron también, Cunoniaceae, Melastomataceae y Symplocaceae. A su vez, se registró en estos fragmentos una diversidad menor a la línea continua del bosque, como fue afirmado por Jørgensen y Ulloa-Ulloa (1994), Wille, Hooghiemstra, Hofstede, Fehse, y Sevink (2002) y Ramírez et al. (2009), donde plantean que estas formaciones son distintas al bosque y están adaptadas a las condiciones del ecosistema donde se encuentran.
Cuatrecasas (1958), Luteyn (1999) y Rangel-Ch. (2000) afirman que el páramo propiamente dicho, está dominado por macollas, rosetas, hierbas y arbustos enanos. Sin embargo, en este trabajo se encontraron algunas especies dispersas, como arbolitos y arbustos, característicos del bosque andino; así también, se registraron especies de Hypericum, Espeletia, Espeletiopsis, Arcytophyllum, Calamagrostis y Chusquea como dominantes en los herbazales de las localidades muestreadas. Otro aspecto relevante, corresponde a la variación registrada en el límite inferior en cada localidad del herbazal así, para Duitama se encuentra en la cota de los 3 593 m, en Belén desde los 3 766 m y Onzaga-Susacón a 3 444 m. Sin embargo, Rangel-Ch. (2000) mencionó que el páramo se encuentra sobre los 3 500 m, lo cual difiere con lo registrado en este trabajo.
La diversidad verdadera reflejó que el páramo de Onzaga-Susacón fue el más diverso (68) con relación a las demás áreas de estudio; a su vez, la diversidad es mayor hacia el oriente del complejo en una proporción de 1.5 veces más diverso Onzaga-Susacón que Duitama. Esto puede estar favorecido por presentar un mayor régimen de humedad, debido a la ubicación geográfica y las lluvias orogénicas provenientes del valle del Magdalena (Morales et al., 2007).
A partir del análisis de diversidad beta, se observa que las estaciones de bosque, y los herbazales de Duitama y Belén, pueden ser definidas tanto por composición de especies como por su estructura; sin embargo, el herbazal de Onzaga-Susacón se encuentra más relacionado con las estaciones de arbustal, excepto la de su misma localidad, con la cual, se registra un 70 % de disimilitud. Lo anterior, muestra que, para el área de estudio la composición de especies se encuentra relacionada con la altitud, debido a que el transecto de Onzaga-Susacón se encuentra por debajo de las otras localidades.
A partir de los agrupamientos a nivel de familias, se encontró que la vegetación de arbustales, comparte una mayor similitud con la de herbazales, lo cual indica que hay especies del páramo que han migrado hacia la zona de transición bosque-páramo, ello sugiere que se considere como área de conservación junto con los páramos, ya que juegan un papel fundamental en la preservación de especies de la alta montaña.
La dinámica de la franja de transición entre el bosque y el páramo en la cordillera Oriental, juega un rol importante a la hora de conocer procesos bióticos que allí ocurren; conocer la diversidad de estas áreas es fundamental para que sean tenidas en cuenta para la toma de decisiones a nivel nacional, por ende, estudios de este tipo son necesarios para la protección y preservación de dichas áreas.

