











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
Feeding ecology is considered a putative driver underlying the diversification of cichlid fishes in lacustrine environments (Winemiller, 1990; Winemiller, Kelso-Winemiller, & Brenkert, 1995; Galis & Metz, 1998; Burres et al., 2013; Burres, 2015). Multiple habitats found in freshwater environments (e.g. pelagic, benthic, litoral, ...) promote ecological segregation and sympatric speciation within distribution of the range of a species as well as dietary differences (piscivory, insectivory, detritivory, …) (Horstkotte & Strecker, 2005; Joyce et al., 2005; Barluenga, Stölting, Salzburger, Muschick, & Meyer, 2006). In this way, variation in diet can lead to specialization among individuals of the same community through the differential use of resources, which in the long term can lead to the development of specialized morphologies to exploit a given resource (McKaye & Marsh, 1983; Liem, 1991; Burres, 2015). It has been documented that in cichlids, body and head shapes are determined by foraging strategies (Burress 2015, Burress, 2016). Thus, head shape is closely related with feeding apparatus which in this group of fishes widely differ in structure and dentition. Particularly, oral jaws are likely a structure driving the diversification and radiation in this group (Salzburger, 2009). Besides, in cichlid fishes, the last branchial arch is modified into a second pair of jaws, a very plastic structure that evolved in response to foraging function in closely related species (Liem, 1973; Burres et al., 2013). Hence, foraging preferences in cichlids have acted as a factor promoting changes in head morphology, particularly in oral and pharyngeal jaws as well as in speciation (Fan, Elmer, & Meyer, 2012; Tsuboi, Gonzalez-Voyer, & Kolm, 2014).
Herichthys is the most boreal genus among the Neotropical cichlids (Kullander, 2003) with relatively high number of species and with a high proportion of the various ecomorphologies found among Middle American cichlids (Říčan, Pialek, Dragova, & Novak, 2016; Peréz-Miranda et al., 2018). Still, there are few studies that deal directly with feeding habits in the genus. A few exceptions are: 1) studies performed on H. minckleyi concluding that the species is polymorphic and show different foraging strategies (Swanson, Gibb, Marks, & Hendrickson, 2003, Trapani, 2003) and 2) and evaluation of the diet through the ontogeny of H. molango (Díaz-Pardo & Guerra-Magaña 1994). For the rest of the species of the genus, there are inferences of dietary strategy based on cranial and lower pharyngeal jaw morphologies (López-Fernández, Arbour, Winemiller, & Honeycutt, 2013; Říčan et al., 2016) but direct stomach-content analyses are missing. In the present work we describe the stomach content of the species of the genus Herichthys, analyze the lower pharyngeal jaw shapes using geometric morphometrics and compare the diet among sympatric species to evaluate the existence of trophic segregation within the genus.
Materials and methods
Study sites: Includes most of the Pánuco river basin and the endorheic basin of Cuatro Ciénegas), between the coordinates range (20°3'50" - 29°30'8.6" N & 96°52'48" - 101°19'56" W) (for further details see appendix 1 or figure 1 in Pérez-Miranda et al. (2018)).
Stomach content: A total of 367 individuals from 11 species were analyzed: H. bartoni (Bean, 1892) (n = 10), H. labridens (Pellegrin, 1903) (n=10), H. pame (De la Maza-Benignos & Lozano-Vilano, 2013) (n = 18), H. steindachneri (Jordan & Snyder, 1899) (n = 12), H. pantostictus (Taylor & Miller, 1983) (n = 126), H. deppii (Heckel, 1840) (n = 20), H. tepehua (De la Maza-Benignos, Ornelas-García, Lozano-Vilano, García-Ramírez, & Doadrio, 2015) (n = 20), H. tamasopoensis (Artigas-Azas, 1993) (n = 17), H. cyanoguttatus Baird & Girard, 1854 (n = 10), H. carpintis (Jordan & Snyder, 1899) (n = 104) and H. minckleyi (Kornfield & Taylor, 1983) (n = 20). For the two widespread species H. pantostictus and H. carpintis a much larger sample covering their large distribution areas was analyzed. In the case of H. molango (De la Maza-Benignos & Lozano-Vilano, 2013), data were taken from Díaz-Pardo & Guerra-Magaña (1994). Stomach contents of H. pantostictus and H. carpintis were evaluated for each single locality due to these two species being widely distributed, while, the other species are micro-endemic. The first third of gut's anterior portion was removed from each individual and its content was mixed into a single sample for each species or locality. Food items were identified to the lower taxonomic level and the proportion of each one in the pooled sample was estimated through a volumetric method (Hyslop, 1980). To assess trophic overlapping, Pianka index of niche overlap (pi) was used (Pianka, 1974). This is a symmetric index that ranges from zero for no niche overlap to one for complete niche overlap. Niche overlapping was estimated only for localities where at least two species are sympatric; the species pair H. bartoni - H. labridens in the headwaters of the Río Verde, San Luis Potosí México; H. pame, H. steindachneri and H. tamasopoensis in the Río Gallinas, San Luis Potosí, México and finally, nine sympatric localities for H. carpintis and H. pantostictus were examined throughout their geographic distribution (Digital Appendix 1). In order to designate trophic groups and to represent species in a consumption gradient, a principal component analysis was performed; with the arc sin standardized matrix of the food items proportion to cluster each species in a single trophic group. The analysis was performed in XLSTAT (XLSTAT, 2013).
Geometric morphometrics: A total of 303 lower pharyngeal jaws (LPJ) were analyzed in the twelve species of the genus (one image of LPJ per species is available in Digital Appendix 2). Ten LPJs were analyzed for each one of the restricted distributed species (H. bartoni, H. labridens, H. pame, H. steindachneri, H. molango, H. deppii, H. tepehua, H. cyanoguttatus, H. minckleyi and H. tamasopoensis). For the widespread H. carpintis and H. pantostictus 92 LPJs from 11 localities and 111 LPJs from 13 localities were analyzed, respectively. Each LPJ was photographed with a digital camera and nine landmarks were recorded following Klingenberg, Barluenga and Meyer (2002) (Fig. 1). Photographs were digitized in tpsdig (Rohlf, 2005) and Procrustes coordinates were estimated in MorphoJ 1.03C (Klingenberg, 2011). To eliminate the allometric effect associated with growth, a multivariate regression analysis was performed, using the Procrustes distances as the dependent variable and the size of the centroid as the independent variable. The adjusted Procrustes distances were used as descriptors of the level of differences among lower pharyngeal jaw shapes between the species; the significance of the differences was evaluated using a permutation test with 10 000 iterations (Elmer, Kusche, Lehtonen, & Meyer, 2010) in MorphoJ 1.03C (Klingenberg, 2011). After it, following the criteria established by Říčan et al. (2016) species were classified into trophic morphs: H. pame, H. tepehua, H. deppii, H. labridens, H. bartoni and H. pantostictus were classified as filter-feeders or sifters; H. minckleyi, H. tamasopoensis, H. cyanoguttatus, H. molango and H. carpintis were classified as detritivorous, and H. steindachneri was classified as piscivorous. Together with the here obtained lower pharyngeal jaw shape, previously generated geometric morphometrics head shape Mejía, Pérez-Miranda, León-Romero, Soto-Galera and Luna (2015) and Pérez Miranda et al. (2018) were used to assess if and how differences in the shapes differ between observed feeding habits. An additional analysis was performed in order to compare the lower pharyngeal jaw shape of the sympatric species H. bartoni - H. labridens and H. tamasopoensis - H. steindachneri - H. pame. Above mentioned analysis were performed with the residuals of the regression in a canonical analysis. Similar to the Procrustes distances, the significance of the differences was evaluated using a permutation test with 10 000 iterations in MorphoJ 1.03C (Klingenberg, 2011). Finally, we perform a multiple stepwise regression analysis between the relative warps of the lower pharyngeal jaw and head shape against the proportion of food items as well as scatter plots of the relative warps scores in Statistica 10.0 (StatSoft, 2011) following the procedure described in Edwards, Tolley, Vanhooydonck, Measey and Herrel (2013).
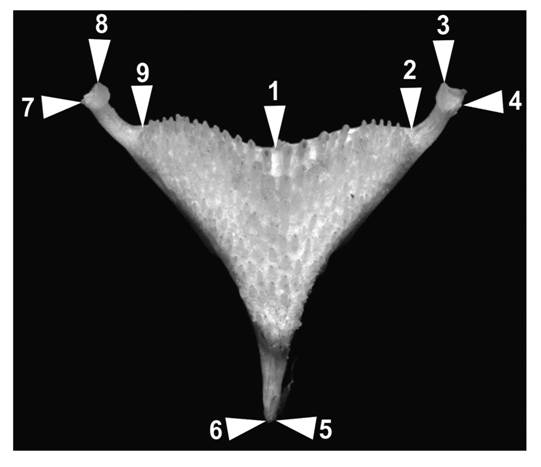
Fig. 1 Landmarks used in the geometric morphometric analysis of the lower pharyngeal jaw (LPJ). 1-middle point of the jaw; 2-last tooth lateral upper right; 3-upper point of the right muscular process; 4- lateral point of the right muscular process; 5- lower right point of the stem; 6-lower left point of the stem; 7-lateral point of the left muscular process; 8-upper point of the left muscular process; and 9- last tooth lateral upper left.
Results
Seven types of food items were recovered in the stomach content of the genus Herichthys: arthropods, non-arthropods invertebrates, snails, fishes, vascular plants, algae, and detritus. Six species showed a clear predominance of a single type of food items (H. tamasopoensis 93.7 % detritus; H. steindachneri 76.9 % fishes; H. pame 76.5 % vascular plants, H. bartoni 61.1 % algae, H. carpintis 60.6 % detritus and H. deppii 57.1 % vascular plants; Table 1). This analysis thus found one strong piscivore (H. steindachneri), one algivore (H. bartoni), two detritivores (H. tamasopoensis, H. carpintis) and two plant-eaters (H. pame and H. deppii).
The remaining six species showed more than one dominant food type; Herichthys labridens consumed mostly arthropods (57.6 %) and snails (40.7 %), H. tepehua consumed arthropods (30.4 %) and algae (24.6 %), H. cyanoguttatus consumed algae (38.5 %) and detritus (38.5 %) in equal amount, H. minckleyi fed on vascular plants (50.7 %) and detritus (33.3 %), H. pantostictus consumed arthropods (39.9 %), detritus (22.7 %) and algae (20.0 %), and H. molango consumed comparable amounts of detritus (27 %), vascular plants (26.5 %), arthropods (24 %) and algae (20.7 %) (Table 1).
TABLE 1 Proportion of diet items (%) present in the stomach contents of 12 species of the genus Herichthys
| Species | Arthropodes | Invertebrates non arthropods | Snails | Fishes | Vascular plants | Algae | Detritus |
| H. bartoni | 11.11 | 0 | 0 | 0 | 0 | 61.11 | 27.78 |
| H. labridens | 57.62 | 0 | 40.67 | 0 | 1.71 | 0 | 0 |
| H. pame | 10.20 | 0 | 5.10 | 4.08 | 76.53 | 0 | 4.08 |
| H. steindachneri | 15.38 | 0 | 0 | 76.92 | 7.69 | 0 | 0 |
| H. molango | 24 | 1.75 | 0 | 0 | 26.5 | 20.75 | 27 |
| H. pantostictus | 39.97 | 0.06 | 4.52 | 7.16 | 6.07 | 19.55 | 22.67 |
| H. deppii | 11.11 | 0 | 3.17 | 1.58 | 57.14 | 17.46 | 9.53 |
| H. tepehua | 30.43 | 10.14 | 7.25 | 7.25 | 8.69 | 24.64 | 11.59 |
| H. tamasopoensis | 2.08 | 0 | 0 | 2.08 | 2.08 | 0 | 93.75 |
| H. cyanoguttatus | 15.38 | 0 | 0 | 0 | 7.69 | 38.46 | 38.46 |
| H. minckleyi | 0.76 | 3.78 | 1.52 | 9.85 | 50.76 | 0 | 33.33 |
| H. carpintis | 14.94 | 0.32 | 3.08 | 9.55 | 8.70 | 2.85 | 60.56 |
A close inspection of the two widely distributed species showed that individuals of six localities of H. pantostictus deviate from the predominant consumption of arthropods, being detritus and algae the most abundant items. On the other hand, only two of the eleven analyzed populations of H. carpintis showed deviations from detritus consumption, with vascular plants and arthropods as the most abundant items (Table 2).
Pi values were generally low among the different performed comparisons. In the sympatric species-pair H. bartoni and H. labridens the Pianka index value was 0.13. In the three sympatric species of the Río Gallinas, values among comparisons were also low, between H. tamasopoensis and H. pame the index value was 0.08; between H. tamasopoensis and H. steindachneri was barely 0.03; and between H. steindachneri and H. pame the values was 0.17, suggesting low levels of trophic overlapping between sympatric species.
Among the nine sympatric localities of H. carpintis and H. pantostictus, Pianka's index of niche overlap showed values that ranged from no overlapping in localities two and nine (Pi = 0.05) to high diet overlapping in localities one (Pi = 0.72), four (Pi = 0.76) and eight (Pi = 0.97) (Table 2).
TABLE 2 Proportion of the distinct diet items found in stomach contents and Pianka´s index of niche overlap for the populations of H. pantostictus (Hp) and H. carpintis (Hc) collected in sympatric and allopatric conditions
| Sympatry | Allopatry | ||||||||||||||||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 10 | 11 | 12 | 13 | |||||||||||||||
| Diet items | Hc | Hp | Hc | Hp | Hc | Hp | Hc | Hp | Hc | Hp | Hc | Hp | Hc | Hp | Hc | Hp | Hc | Hp | Hc | Hc | Hp | Hp | Hp | Hp | |||||
| Vascular plants | 14.3 | 3.45 | 0 | 0 | 6.45 | 7.14 | 75 | 40.8 | 0 | 0 | 0 | 21.4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6.25 | 0 | 0 | 0 | |||||
| Arthropodes | 0 | 6.9 | 3.57 | 41.7 | 6.45 | 42.8 | 6.25 | 40.8 | 18.2 | 100 | 27.8 | 14.3 | 7.69 | 62.5 | 27.3 | 40 | 9.09 | 26.1 | 50 | 8 | 62.5 | 11.8 | 40 | 30.3 | |||||
| Snails | 0 | 0 | 0 | 0 | 0 | 14.3 | 0 | 0 | 0 | 0 | 11.1 | 28.6 | 0 | 0 | 0 | 0 | 22.7 | 0 | 0 | 0 | 0 | 0 | 16 | 0 | |||||
| Detritus | 57.1 | 48.3 | 89.3 | 0 | 64.5 | 14.3 | 0 | 8.16 | 81.8 | 0 | 55.6 | 7.14 | 76.9 | 37.5 | 72.7 | 60 | 68.2 | 0 | 20 | 80 | 18.7 | 0 | 40 | 60.6 | |||||
| Fishes | 28.6 | 3.45 | 3.57 | 25 | 3.22 | 0 | 18.7 | 10.2 | 0 | 0 | 5.56 | 28.5 | 15.3 | 0 | 0 | 0 | 0 | 8.69 | 30 | 0 | 12.5 | 0 | 4 | 0.75 | |||||
| Algae | 0 | 37.9 | 0 | 33.3 | 19.3 | 21.4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 65.2 | 0 | 12 | 0 | 88.2 | 0 | 7.57 | |||||
| Invertebrates non artrhopods | 0 | 0 | 3.57 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.75 | |||||
| Pianka´s index | 0.72 | 0.05 | 0.46 | 0.76 | 0.22 | 0.41 | 0.59 | 0.97 | 0.05 | ||||||||||||||||||||
The first two axes of the principal component analysis explained 41.57 % of the variation (Fig. 2A). In the first component, the items with the major loadings were detritus and arthropods, while in the second component the major loadings corresponded to arthropods and vascular plants. The analysis based on the first two axes recovered four distinct groups of species. The first group included detritivores (H. tamasopoensis, most of the localities of H. carpintis, and two of the localities of H. pantostictus); the second group included invertivore species that consume arthropods and snails (H. labridens, H. tepehua, most of the localities of H. pantostictus, and one locality of H. carpintis); the third group included herbivores (H. deppii, H. pame, H. minckleyi and one locality of H. carpintis). The fourth group is centrally located in the multivariate space and includes the strong algivore H. bartoni, the algivorous-detritivorous H. cyanoguttatus, and three localities of H. pantostictus with a high proportion of algae in their diet. This group is better recovered in the PCA analysis of axes one and three (Fig. 2B). Finally, H. steindachneri was recovery isolated from all of the above-mentioned groups due to the almost exclusively consumption of fishes. Results from the PCA ordination were consistent with the classification based on the predominant food-type (Table 1).
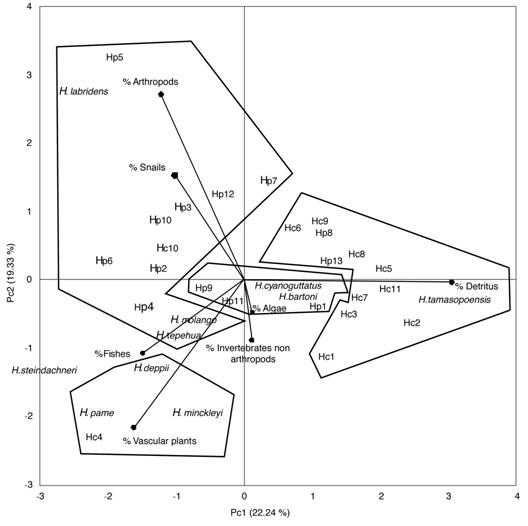
Fig. 2 Principal component analysis of gut contents for all species of the genus Herichthys. A. PC1 vs PC2. B. PC1 vs PC3.
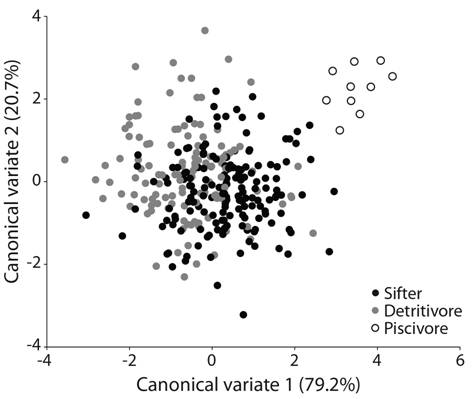
Fig. 3 Canonical variate analysis of the shape of the lower pharyngeal jaw (LPJ) for the three different foraging modes present in the genus Herichthys.
The geometric morphometric analyses of the lower pharyngeal jaw shape (Fig. 3) and head shape (Fig. 4) found in both cases the highest Procrustes distances between detritivorous and piscivorous species (d = 0.084, P < 0.05 and d = 0.133, P < 0.05, respectively) and the lowest Procrustes distances between detritivores and sifters (d = 0.016, P < 0.05 and d = 0.066, P < 0.05, respectively). A better separation of the putative feeding ecomorphs was obtained in the analysis of head shape (Fig. 4) and all p values were significant (Table 3). The CVA analysis, only allowed a clear separation of the piscivorous shape. Finally, the shapes identified by means of the CVA analysis of the lower pharyngeal jaw amongst sympatric distributed species (H. labridens - H. bartoni and H. pame - H. steindachneri - H. tamasopoensis) (Digital Appendix 3), were congruent with low levels of overlap detected via stomach content analysis and Pi.
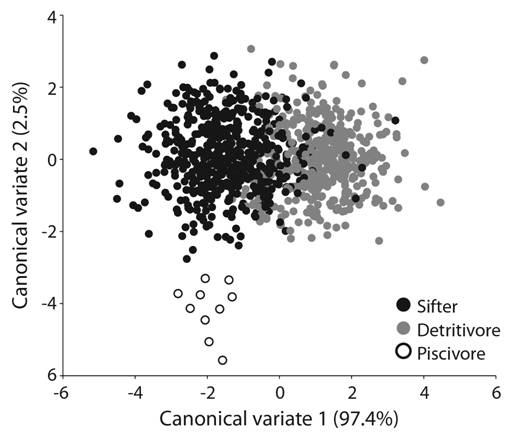
Fig. 4 Canonical variate analysis of the shape of the head for the three different foraging modes present in the genus Herichthys.
TABLE 3 Procrustes distances between groups for the shapes of the lower pharyngeal jaw (LPJ) and the head for the three foraging modes present in the genus Herichthys
| Lower Pharingeal Jaw | Head | ||||
| Detritivorous | Piscivorous | Piscivorous | Detritivorous | ||
| Piscivorous | 0.0845* | Detritivorous | 0.1336* | ||
| Sifter | 0.0162* | 0.072* | Sifter | 0.0924* | 0.0667* |
P < 0.05 values are indicated with an asterisk (*).
The results of the forward stepwise multiple regression showed that the relative warps (RW) 2 and 3 for the lower pharyngeal jaws (LPJ) showed a relationship with arthropods, invertebrates, fishes, vascular plants, algae and detritus, meanwhile, the three relative warps of the head shape (HD) showed a relationship with any of the food items (Digital Appendix 4). On the other hand, the scatter plots graphs of the RW showed that the sympatric species H. bartoni and H. labridens are clearly different in shapes of both LPJ and HD, meanwhile, for the sympatric species H. steindachneri - H. pame and H. tamasopoensis, the first one is clearly different in both shapes and the differences in the latter two are evident only in the HD (Digital Appendix 4).
Discussion
In summary, our stomach content analysis evidenced that in all but one of the micro-endemic sympatric species, one food type predominated (61.1 %, 76.5 %, 76.9 % and 93.7 % single type predominance); whereas, among the non-sympatrically distributed species only one species had strong dominance of just one food type (Table 1).
The results found in this study are consistent with previous studies of other species found in the same genus. For example, Swanson et al. (2003) reported that in general H. minckleyi feeds of detritus, algae, soft body invertebrates, snails and fishes (regardless of the morph), Hulsey et al. (2006) found that detritus represents the major component in the diet of H. minckleyi (mineral and vegetal detritus > 66 %), Magalhaes, Ornelas-Garcıa, Leal-Cardin, Ramírez and Barluenga (2015) reported the occurrence of gastropods, insects, zooplankton, plants, algae and detritus in H. minckleyi. Also, Hulsey et al. (2006) and Magalhaes et al. (2015) found that mollusks represented up to (90 %) of the stomach contents in the molariform ecomorph, while detritus was the major component in the papilliform ecomorph (55 %). The great variation found in the stomach content of H. minckleyi is likely the result of the lower pharyngeal jaw polymorphism consisting on two pharyngeal morphotypes: one with molariform and one with papilliform teeth (Trapani, 2003a; Trapani, 2003b). Thus, the papilliform ecomorph preferentially feeds soft diet items, while the molariform ecomorph consumes both soft items and large quantities of snails and bivalves (Swanson et al., 2003; Hulsey et al., 2006; Říčan et al., 2016; Hernández, Espinosa-Pérez, & Souza, 2017). The low proportion of snails recovered in this study (1.5 %) is potentially explained by the fact that the majority of the specimens analyzed correspond to the papilliform morph (80 %), also the most frequently found ecomorph in the wild (Swanson et al., 2003) which preferentially feeds on plant material (Hulsey, 2006; Hulsey et al., 2006).
For the remaining species most of the putative preferred diet has been inferred from head and jaw teeth morphologies, such as H. steindachneri, characterized as piscivorous (Miller, Minckley, & Norris, 2005; De La Maza-Benignos et al., 2015; Říčan et al., 2016) and confirmed as such in this study (76.9 % of fishes). Říčan et al. (2016) classified the rest of the species of the genus Herichthys as detritivorous, based on morphological comparisons with other Middle American cichlid genera. Our study revealed that they include species with a broad diet that includes plants, algae, small arthropods, fine particles and mollusks. All above mentioned authors classified H. labridens as a molluskivorous species based on its stout LPJ and its medial teeth, although the current analysis showed that the most frequent items preyed by this species were arthropods (57.6 %) followed by snails and shells (40.6 %).
Dietary comparisons between the sympatric species H. labridens and H. bartoni shows a clear trophic segregation (arthropods and snails vs. algae) with low overlapping and strong differences in the lower pharyngeal jaw shape (Digital Appendix 3). Similar clear and strong separation was found in the sympatric species H. tamasopoensis, H. steindachneri, and H. pame (Digital Appendix 3).
H. labridens and H. bartoni are lacustrine, surviving now in much reduced spring lakes in an area of a former paleo-lake in the region of Río Verde basin (Planer-Friedrich, Armienta, & Merkel, 2001). The two species probably arose in sympatry (Říčan et al., 2016; Pérez-Miranda et al., 2018) as a product of an assumed niche segregation in a lacustrine setting as has been documented in the genus Amphilophus in the crater lakes of Nicaragua (Barluenga et al., 2006).
H. steindachneri and H. pame clearly evolved in sympatry in a riverine setting (Říčan et al., 2016; Pérez-Miranda et al., 2018). In this case the speciation process probably has a parallel in the Paraná and Uruguay Crenicichla complexes in South America (Piálek, Říčan, Casciotta, Almirón, & Zrzavý, 2012; Burres et al., 2013; Burres et al., 2018). The third sympatric species H. tamasopoensis is unrelated to the species pair and has colonized the area secondarily (Říčan et al., 2016; Pérez-Miranda et al., 2018), this species probably arose through parapatric speciation from an ancestral population of H. carpintis that shared the same predominantly detritivorous diet.
The two widespread and sympatric H. pantostictus and H. carpintis showed contrasting results. While the diet of H. pantostictus was the most variable within the genus, ranging from arthropods to algae to detritus, H. carpintis consumed almost exclusively detritus through its complete geographic distribution (Fig. 2). One possible explanation is that in a widely distributed species diet can vary between populations as has been reported in the stomach content and lower pharyngeal jaw shape in Astatotilapia burtoni populations (Theis, Ronco, Indermaur, Salzburger, & Egger, 2014). Both the high variation in food items found in H. pantosticus and the low variation found in H. carpintis have been found in the upper Zambezi river, for example, the carnivorous Serranochromis species showed high variation in the consumed food items, meanwhile, the detritivorous species of the genus Oreochromis and Tilapia showed little variation in their diet (Winemiller, 1991; Winemiller & Kelso-Winemiller, 2003).
The morphologically most distinctive species in head shape and LPJ shape is H. steidanchneri as demonstrated by clear separation in CVA analysis. This is consistent with the results of Burress (2016) who found that strict piscivorous (and also molluskivorous) species have a markedly different lower pharyngeal jaw morphology in comparison with omnivorous, herbivorous and invertivorous species. Nevertheless, Burress (2016) wrongly attributed H. cyanoguttatus as a molluskivorous species, probably because of its confusion with H. minckleyi (Hulsey et al., 2006), as well as Hulsey (2006) who classified H. cyanoguttatus as a molluskivorous species even when the proportion of mollusks in its diet represented less than 5 %, as in the present study.
The head shape allowed for better morphologically-based separation of feeding groups than the shape of the lower pharyngeal jaw in the genus Herichthys (Říčan et al., 2016). Similar to the present study, López-Fernández et al. (2013) and Říčan et al. (2016) found that the head shapes in H. steindachneri and H. bartoni are more differentiated than the head shapes in H. cyanoguttatus and H. carpintis, a pattern similar to that recovered in other Middle American cichlids such as Amphilophus, Vieja, Nandopsis and Amatitlania where the head morphologies are more variable compared to a more conserved body shapes (e.g. Fig. 7 in Říčan et al., 2016). Finally, further studies would be necessary in the future to clearly understand the role of diet in the diversification process due that it can vary both with ontogeny and seasonally.

