











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
Altitudinal gradients allow to study evolution and selective forces in changing environments, as traits associated with survival and fitness may show phenotypic plasticity and local adaptation (Byars, Papst, & Hoffmann, 2007; Laine, 2008; Hughes, 2012). Environmental conditions along different elevations change rapidly at a small geographic scale, allowing to naturally control for potential latitudinal effects such as spatial changes of light intensity and photoperiod (Hodkinson, 2005). In organisms with broad elevational distributions, populations at different elevations will experience contrasting environmental conditions, including abiotic (e.g. temperature, precipitation, radiation and partial pressure of atmospheric gases) and biotic factors (e.g. food abundance/quality, predatory and parasitic pressures) (Hodkinson, 2005) that shape their morphology, physiology and performance (Dahlhoff & Rank, 2007).
Temperature is one of the most important abiotic factors shaping animal morphology and physiology. Insect populations living at high elevations (and associated cold temperatures) might respond to the environment by increasing their body size (Mousseau & Roff, 1989; Blanckenhorn & Fairbairn, 1995) and it has been suggested that the survival of insects at colder sites is favored by having larger body sizes (Fischer & Fiedler, 2000), as food intake is higher and food transformation into body mass occurs more efficiently at colder temperatures (Karl & Fischer, 2008). However, this is not a general pattern, as for some species there is no evident relationship between altitude and body size (Sanabria-Urbán et al., 2015). Despite contrasting evidence in the response of body size to different environmental temperatures, there is no doubt that insects and other ectotherms are particularly sensitive to temperature changes, as they depend on environmental conditions to regulate their own temperature (Stevenson, 1985). Because body size affects survival and reproductive success of insects (Blanckenhorn, 2000), further studies need to evaluate the relationship between body size and temperature of insect populations at different elevations (Stevenson, 1985). Reaching larger body sizes implies higher metabolic requirements for growth and development, which may favor higher energetic storage at higher altitudes (Lease & Wolf, 2010).
In addition to temperature, biotic factors such as parasite pressure can vary with altitude, shaping energetic condition and host immune responses (Zamora-Vilchis, Williams, & Johnson, 2012). For example, low temperatures associated with high altitudes prevent the development of some insect parasites, such as malaria in Anopheles mosquitoes (Bødker et al., 2003). As altitude governs the distribution of parasites (van Rooyen, Lalubin, Glaizot, & Christe, 2013), living at different altitudes also requires physiological mechanisms of immune regulation that allow to deal efficiently against different rates of parasitism. Evidence in insects show that high temperatures are associated with high immune responses (Catalán, Wozniak, Niemeyer, Kalergis, & Bozinovic, 2012). However, the extent to which temperature affects insect immune response depends on other factors (Murdock, Paaijmans, Cox-Foster, Read, & Thomas, 2012). For example, in the broadly distributed butterfly Lycaena tityrus, local adaptation drives the intensity of the immune response, as populations at high elevation show stronger melanization immune response at colder temperatures, whereas populations at low elevations show stronger melanization response at warmer temperatures (Karl, Hoffmann, & Fischer, 2010). In other cases, the immune response is maximum at extreme temperatures, whether or not they are preferred by insects (Murdock et al., 2012). Because maintaining a strong immune response involves a high energetic investment, food availability and energetic condition (González-Tokman, Córdoba-Aguilar, & Forbes, 2012) may also vary with altitude and define the strength of immune responses; as a consequence, immune systems rapidly become locally adapted at different elevations (Schmid-Hempel, 2005; Sternberg & Thomas, 2014).
In this study, we measured the body size, energetic condition and immune response in two populations of two species of rolled-leaf beetles (Chrysomelidae) present at low (i.e., rainforest) and high (i.e., montane forest) elevations, in Costa Rica (García-Robledo, Kuprewicz, Staines, Erwin, & Kress, 2016). We studied two species that inhabit the same localities, but that differ in body size and feeding regime. Cephaloleia belti is a small polyphagous beetle, while Chelobasis bicolor is a larger species with a more restricted feeding regime. We predicted that body size and energetic condition would be higher at high elevation. On the other hand, as parasitic pressure decreases with altitude (Hodkinson, 2005), we predicted that the immune response would be lower at high elevations in both species, reflecting local adaptation or phenotypic plasticity to deal with local conditions.
Materials and methods
The present study was carried out in June 2016 in Costa Rica at two localities along the Barva Volcano elevational gradient. One location was in La Selva Biological Station, a tropical rain forest at 50 m elevation (10°25’58’’ N - 88°00’13’’ W; temperature range: 20-32 °C). The other location was around 40 km south, in Braulio Carrillo National Park, at 2 000 m elevation (10°10’47’’ N - 84°06’46’’ W; temperature range 10-20 °C). The area receives around 4 000 mm of rain annually and reaches 4 500 mm in higher lands (Timm, Wilson, Clauson, LaVal, & Vaughan, 1989). We selected two species of rolled-leaf beetles, the first species, Cephaloleia belti, is polyphagous, feeding on 18 host plants in the order Zingiberales. The second species, Chelobasis bicolor, feeds only on four host plants in the family Heliconiaceae (García-Robledo, Erickson, Staines, Erwin, & Kress, 2013; Schmitt & Frank, 2013). Individuals of C. belti and Ch. bicolor were collected in the tropical rain forest at 50 m elevation on Heliconia latispatha and H. irrasa (Heliconiaceae). Individuals in the montane forest at 2 000 m elevation were collected on Heliconia lankesteri (Heliconiaceae). Sample sizes are given in Table 1.
TABLE 1 Body length and physiological traits of rolled-leaf beetles in a tropical rain forest (50 m) and a tropical montane forest (2 000 m)
| Chelobasis bicolor | Cephaloleia belti | ||||||||
| 2 000 m | 50 m | 2 000 m | 50 m | ||||||
| Females (N = 10) | Males (N = 11) | Females (N = 19) | Males (N = 23) | Females (N = 16) | Males (N = 22) | Females (N = 26) | Males (N = 45) | ||
| Body length (mm) | 12.87 ± 0.46 | 11.51 ± 0.64 | 9.81 ± 1.03 | 9.27 ± 0.44 | 6.87 ± 0.30 | 6.60 ± 0.29 | 6.41 ± 0.32 (N = 27) | 5.98 ± 0.27 | |
| Dry body mass (mg) | 37.9 ± 4.0 | 28.5 ± 3.1 | 18.8 ± 4.7 | 16.0 ± 2.52 | 4.23 ± 0.65 | 3.59 ± 0.70 | 3.72 ± 1.01 | 3.38 ± 0.78 (N = 44) € | |
| Lipid mass (mg) | 3.35 ± 1.16 | 2.35 ± 1.08 | 1.35 ± 1.21 | 1.04 ± 0.98 | 0.41 ± 0.28 | 0.51 ± 0.39 | 0.19 ± 0.29 | 0.37 ± 0.35 (N = 44) € | |
| Muscle mass (mg) | 23.7 ± 4.4 | 17.4 ± 2.6 | 12.0 ± 2.6 | 9.25 ± 1.75 | 3.38 ± 1.03 | 2.74 ± 0.80 | 2.58 ± 0.84 | 2.35 ± 0.83 (N = 44) € | |
| Lipid/Muscle (proportion) | 0.15 ± 0.07 | 0.14 ± 0.06 | 0.11 ± 0.11 | 0.11 ± 0.09 | 0.11 ± 0.08 (N = 15)* | 0.22 ± 0.20 | 0.10 ± 0.20 | 0.19 ± 0.25 (N = 44) € | |
| Melanization (relative) # | 0.59 ± 0.13 (N = 7) | 0.57 ± 0.15 (N = 7) | 0.40 ± 0.14 (N = 10) | 0.29 ± 0.16 (N = 18) | 0.44 ± 0.10 (N = 15) | 0.37 ± 0.14 (N = 19) | 0.31 ± 0.14 (N = 23) | 0.29 ± 0.13 | |
| * One outlier was removed. # Some implants could not be recovered for melanization measurements, reducing sample size. | € After removing some implants, the body was damaged and was not used for other physiological measurements. | Data are shown as mean ± s. d. Sample sizes are shown in parentheses. | |||||||
Beetles were transported to the laboratory located at La Selva Biological Station, where they were kept at 20 °C with natural light: darkness cycle, and fed ad libitum with the rolled young leaves of the host plant species where they were collected (H. latispatha or H. irrasa for the low elevation populations and H. lankesteri for the high elevation populations). Twenty four hours after collection, a nylon implant (2 mm long x 0.2 mm width, previously sanded with fine sandpaper in order to remove the wax cover) was inserted between the thorax and the elytra, close to the scutellum. Half of the implant was left out of the beetle. Twenty-four hours later the beetles were sacrificed in the freezer (-20 °C) and the implant was removed with forceps for subsequent measurement of melanization immune response (see methods below). The insect body was used for measuring body size, body mass, lipid mass and muscle mass (see methods below).
Melanization is a main component of insect immune response that is a good indicator of general physiological and immune condition and is considered a good measure of insect response against natural parasites such as mites and nematodes (Pomfret & Knell, 2006). The removed nylon implants were placed under a stereoscopic microscope (Zeiss Stemi SV 11) and photographed at 1.2X with an adapted digital camera MU1400 using the software AmScope (www.amscope.com). The implant was photographed from two different positions to have a better estimator of the total surface area of the thread covered by melanin. The images were transformed to gray scale and we calculated implant darkness with respect to the section of the implant that remained out of the insect’s body (i. e. not melanized), using Adobe Photoshop 7.0.1; see similar procedures in (González-Tokman, Córdoba-Aguilar, & Forbes, 2012). The average of both pictures was considered total implant darkness.
After removing the implant from each individual, we measured body length (from the clypeus to the tip of the pygidium) with a digital caliper (± 0.1 mm). Beetles were then dried in an oven at 60 °C for 24 h and weighed to the nearest 0.1 mg to obtain dry body mass. To quantify lipid content, dried individuals were placed in chloroform for 48 h to extract lipids. Beetles were re-dried for another 24 h at 60 °C and re-weighed, and the weight difference was considered the lipid content (Lee, Raubenheimer, & Simpson, 2004). After this, lipid-free individuals were submerged for 48 h in a potassium hydroxide solution (KOH, 0.8 M) to extract the muscle mass. The KOH solution was replaced, and the individuals were left in it for another 48 h. These individuals were re-dried for another 24 h at 60 °C and re-weighed, and the difference with the lipid-free weight was considered muscle content (Marden, 1989). We also analyzed the lipid/muscle mass ratio, as insect individual fitness depends on the optimal consumption of different macronutrients in the diet (Simpson et al., 2015).
Statistical analyses: Data were analyzed with generalized linear models (GLM) and general linear models (LM) when model assumptions were met. When variances were heterogeneous, data were analyzed with generalized least square models (GLS). Analyses considered separately each response variable (body length, body dry mass, lipid mass, muscle mass, lipid/muscle ratio and melanization response) and included the following predictive variables: Species identity, Altitude and Sex, plus the triple interaction Species/Altitude/Sex. In the analyses of dry mass, lipid mass, muscle mass, lipid/muscle ratio and melanization we controlled for body size by including the additive effect of Body Length as a covariate. Normal distribution of model residuals was inferred from normal Q-Q plots; variance heterogeneity was tested with Fligner-Killeen tests and the presence of outliers was evaluated based on Cook’s distances. All analyses were carried out in R software version 3.2.3 (R Development Core Team, 2015), according to (Crawley, 2007; Zuur, Ieno, Walker, Saveliev, & Smith, 2009).
Body length was analyzed with a GLS model that accounted for variance heterogeneity found by Species, Altitude and Sex (Fligner-Killeen tests, P < 0.05). The best variance structure included the triple interaction Species/Altitude/Sex. To explore differences among species we carried out linear models (LM) separately for each species, initially including the interaction effect of Altitude/Sex, and reducing the model based on AIC values.
Body dry mass was analyzed with a GLS model that accounted for variance heterogeneity found by Species and Sex (Fligner-Killeen tests, P < 0.05). The best variance structure included the interaction Species/Sex. The initial model included the interaction effect of Species/Altitude/Sex plus the additive effect of Body Length. The model was reduced based on AIC values. To explore differences within species we carried out linear models (LM) separately for each species, initially including the interaction effect of Altitude/Sex, and reducing the model based on AIC values.
Lipid mass was analyzed with a GLS model that accounted for variance heterogeneity found by Species, Altitude and Sex (Fligner-Killeen tests, P < 0.05). The best variance structure included the effect of Species. To explore differences within species we carried out linear models (LM) separately for each species, initially including the interaction effect of Altitude/Sex and the additive effect of Body Length, and reducing the model based on AIC values.
Muscle mass was analyzed with a GLS model that accounted for variance heterogeneity found by Species, Altitude and Sex (Fligner-Killeen tests, P<0.05). The best variance structure included the effect of Species/Altitude. To explore differences within species we carried out a GLS to account for heterogeneous variances found by Altitude (Fligner-Killeen tests, P < 0.05) for Ch. bicolor, initially including the interaction effect of Altitude/Sex and the additive effect of Body Length, and removing terms that did not improve model fit based on AIC values. For C. belti we carried out a linear model (LM), initially including the interaction effect of Altitude/Sex and the additive effect of Body length, and reducing the model based on AIC values. The ratio Lipid/Muscle mass was analyzed with a GLS model that accounted for variance heterogeneity found by Species (Fligner-Killeen tests, P < 0.05).
Melanization of nylon implants was analyzed with a LM that included implant darkness as response variable, and Species, Altitude, Sex, the interaction Species/Altitude/Sex and the additive effect of Body Length. The model was reduced based on AIC values.
Results
Body length of rolled-leaf beetles differed by Species, Altitude and Sex (Table 2, Fig. 1). For Ch. bicolor, both males and females showed longer bodies at high altitudes, although the effect of altitude was stronger for females (Table 2, Fig. 1A). For C. belti, individuals were larger at higher altitudes and females were larger than males (Table 2, Fig. 1B).
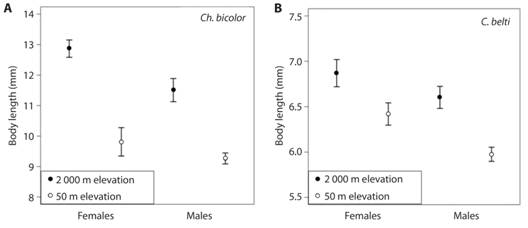
Fig. 1 Body length of male and female rolled-leaf beetles in a tropical rain forest (50 m elevation) and a tropical montane forest (2 000 m elevation). A) Chelobasis bicolor and (B) Cephaloleia belti. Bars represent means ± 95 % C. I.
Dry body mass of rolled-leaf beetles differed by Species, Altitude, Sex and Body Length (Table 2; Fig. 2). For Ch. bicolor, both males and females showed heavier bodies at high altitudes, although the effect was stronger for females; females were heavier than males (Table 2; Fig. 2A). For C. belti, both males and females showed heavier bodies at high altitudes, females were heavier than males and larger individuals were heavier (Table 2, Fig. 2B).
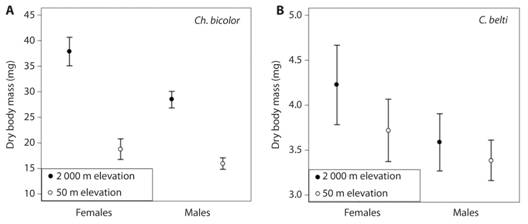
Fig. 2 Dry body mass of male and female rolled-leaf beetles in a tropical rain forest (50 m elevation) and a tropical montane forest (2 000 m elevation). A) Chelobasis bicolor and (B) Cephaloleia belti. Bars represent means ± 95% C. I.
TABLE 2 Effect of altitude on body size and physiological condition of two species of rolled-leaf beetles in a tropical rain forest and a tropical montane forest
| Body length | Dry body mass | Lipid mass | Muscle mass | Lipid/Muscle mass | Melanization | |
| Both species | d.f. = 1.165 | d.f. = 1.162 | d.f. = 1.165 | d.f. = 1.163 | d.f. = 1.167 | d.f.=1.141 |
| Species | F = 3 098.92 P < 0.001 | F = 2 675.78 P < 0.001 | F = 159.34 P < 0.001 | F = 1 870.61 P < 0.001 | F = 3.25 P = 0.073 | F = 10.91 P = 0.001 |
| Altitude | F = 310.46 P < 0.001 | F = 40.92 P < 0.001 | F = 41.05 P < 0.001 | F = 230.10 P < 0.001 | NS | F = 35.92 P < 0.001 |
| Sex | F = 101.26 P < 0.001 | F = 16.04 P < 0.001 | F = 0.94 P = 0.334 | F = 43.52 P < 0.001 | NS | F = 5.47 P = 0.021 |
| Species X Altitude | F = 208.16 P < 0.001 | F = 159.20 P < 0.001 | NS | F = 50.51 P < 0.001 | NS | F = 8.04 P = 0.005 |
| Species X Sex | F = 10.30 P = 0.002 | F = 25.41 P < 0.001 | F = 4.06 P = 0.046 | F = 27.77 P < 0.001 | NS | NS |
| Altitude X Sex | F = 0.34 P = 0.558 | F = 5.83 P = 0.017 | NS | F = 5.71 P = 0.018 | NS | NS |
| Species X Altitude X Sex | F = 7.19 P = 0.008 | F = 8.41 P = 0.004 | NS | NS | NS | NS |
| Body length | NA | F = 262.48 P < 0.001 | F = 63.86 P < 0.001 | F = 309.00 P < 0.001 | F = 6.17 P = 0.014 | NS |
| Ch. bicolor | d.f. = 1.59 | d.f. = 1.58 | d.f. = 1.61 | d.f. = 1.59 | d.f. = 1.38 | |
| Altitude | F = 197.69 P < 0.001 | F = 628.01 P < 0.001 | NS | F = 235.68 P < 0.001 | NA | F = 29.47 P < 0.001 |
| Sex | F = 21.11 P < 0.001 | F = 71.86 P < 0.001 | NS | F = 40.80 P < 0.001 | NA | F = 3.27 P = 0.078 |
| Altitude X Sex | F = 4.70 P = 0.034 | F = 6.59 P = 0.013 | NS | NS | NA | NS |
| Body length | NA | F = 103.53 P < 0.001 | F = 51.18 P < 0.001 | F = 22.56 P < 0.001 | NA | F = 3.47 P = 0.070 |
| C. belti | d.f. = 1.107 | d.f. = 1.104 | d.f. = 1.105 | d.f. = 1.105 | d.f. = 1.102 | |
| Altitude | F = 96.18 P < 0.001 | F = 4.93 P = 0.029 | F = 6.28 P = 0.014 | F = 10.87 P = 0.001 | NA | NS |
| Sex | F = 44.83 P < 0.001 | F = 8.46 P = 0.004 | F = 5.20 P = 0.025 | F = 5.10 P = 0.026 | NA | NS |
| Altitude X Sex | NS | NS | NS | NS | NA | NS |
| Body length | NA | F = 12.04 P < 0.001 | NS | NS | NA | F = 17.75 P < 0.001 |
Significant effects are shown in bold. D.f. = Degrees of freedom. NA = The variable was not included in the model. NS = The variable was not selected by the best supported model.
Lipid mass of rolled-leaf beetles differed by Species, Sex, Altitude and Body Length (Table 2, Fig. 3A). For Ch. bicolor, lipid mass was only explained by the effect of Body Length, as larger individuals had more lipids. Altitude and Sex did not explain differences in lipid mass for this species. For C. belti, males and females showed more lipids at high altitudes and males had more lipids than females (Table 2, Fig. 3A, Fig. 3B).
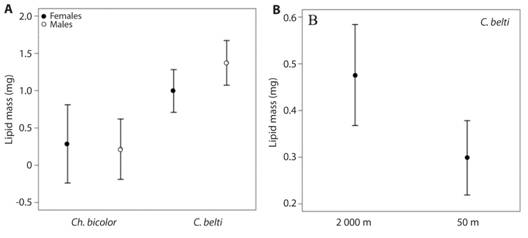
Fig. 3 Lipid content in (A) males and females of two species of rolled-leaf beetles in (B) a tropical rain forest (50 m elevation) and a tropical montane forest (2 000 m elevation). Bars represent means ± 95 % C. I.
Muscle mass of rolled-leaf beetles differed by Species, Altitude, Sex and Body Length (Table 2, Fig. 4). For Ch. bicolor, beetles at high altitudes had more muscle than at low altitudes (Fig. 4A), females had more muscle than males (Fig. 4B), and larger individuals had more muscle. For C. belti, beetles at high altitudes had more muscle than at low altitudes and females had more muscle than males (Fig. 4C, Fig. 4D).
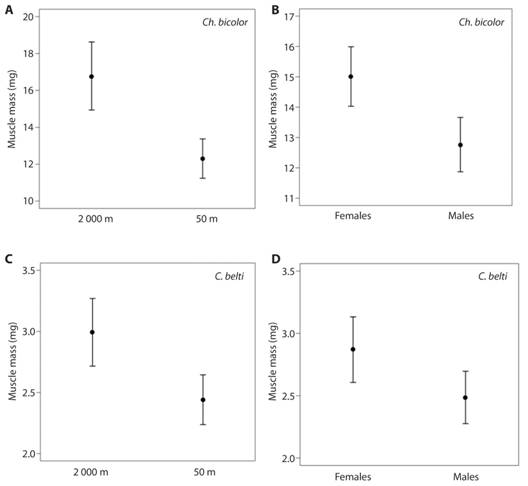
Fig. 4 Muscle content in male and female rolled-leaf beetles in a tropical rain forest (50 m elevation) and a tropical montane forest (2 000 m elevation). (A., B.) Chelobasis bicolor and (C., D.) Cephaloleia belti. Bars represent means ± 95 % C. I.
The lipid/muscle mass ratio of rolled-leaf beetles differed by Species and Body Length, and was not affected by Altitude or Sex (Table 2). C. belti presented marginally higher lipid/muscle values than Ch. bicolor and larger individuals showed higher lipid/muscle values.
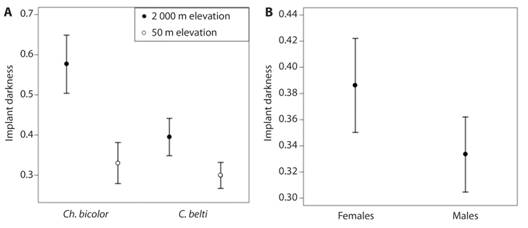
Melanization of nylon implants of rolled-leaf beetles differed by Species, Altitude and Sex (Table 2; Fig. 5). In both species, melanization tended to be higher at high altitudes although the effect was only significant for Ch. bicolor (Fig. 5A). Females had higher melanization than males (Fig. 5B). Body length was positively related to melanization only in C. belti.
Discussion
In this study we found support to our prediction, as both a food specialist, Ch. bicolor, and a food generalist beetle, C. belti, were bigger, heavier and had more muscles (even after controlling for body size) at high altitude compared to low altitude. Regarding energetic reserves, our prediction was only met for C. belti beetles, as individuals were fatter at high altitude. However, contrary to our prediction, both species of beetles also presented stronger melanization immune response at high altitude than at low altitude. These results, as previous evidence (Mousseau & Roff, 1989; Dahlhoff & Rank, 2007), indicate that altitude is fundamental for shaping physiological and morphological insect traits that strongly depend on food supply and parasitic pressure. However, further studies in the same or different insects should be done in different altitudinal gradients and including reciprocal transplants to determine whether the observed patterns of response to altitude are observed in other systems and if they result from local adaptation to altitude or arise from phenotypic plasticity. For example, phenotypic plasticity could mediate evolutionary changes via genetic accommodation (Crispo, 2007), shaping physiological and morphological traits.
We also found that there are general sex differences in both species for all the studied traits. Such differences were maintained at different altitudes in all traits, except for body length and dry body mass. Sex differences in nutrient utilization are common in insects (Schmid-Hempel, 2005) and females usually store more energetic reserves than males due to the high lipid demands of egg production (Lease & Wolf, 2010). Given that males typically invest less resources than females into raising offspring, and invest more resources into competition for mates, males often compromise other functions associated to survival, such as immune responses (Schmid-Hempel, 2005; Córdoba-Aguilar & Munguía-Steyer, 2013). Future studies should evaluate male and female investment in different life history traits (e.g. body size, mate searching, parental investment) related to survival and reproduction in our study system, and how different are energetic requirements at different altitudes in both sexes.
Despite there being diverse environmental factors associated to altitude that could explain variations in insect body size and physiological condition (e.g. temperature, plant growth, phenology, nutrient content and concentration of defensive compounds) (Hodkinson, 2005), it seems likely that food availability and quality could be modulating the expression of all the measured traits, which are dependent on individual nutritional condition. Further studies should evaluate the environmental factors associated to altitude that shape body size and physiological condition in our studied species.
The interaction of Species X Altitude found to explain most traits (except for lipid mass and lipid/muscle ratio) showed that the impact of altitude differed in magnitude among species, although it was always in the same direction. In our studied system, the impact of altitude was higher on Ch. bicolor (specialist species) than on C. belti (generalist species). Such differences have been observed in other system, such as weevils (Family Curculionidae), where species tend to increase body size with altitude, but such increase is not the same for all species (Chown & Klok, 2003). Further studies in chrysomelid beetles should evaluate to which extent food specialization, resource availability or temperature stress differ among species at different altitudes, shaping individual physiology response and morphology (Chown & Klok, 2003; Sørensen, Norry, Scannapieco, & Loeschcke, 2005).
Melanization immune response also differed between species and altitudes. At low altitudes, both species showed similar melanization immune response. However, at high altitudes it was Ch. bicolor, the more specialist species, which showed much higher melanization than the generalist, C. belti. According to Lampert (2012), generalist herbivores are expected to show lower immune responses compared to specialists due to the potential deleterious effects of secondary metabolites present in different plant hosts. Alternatively, the risk of parasitism may be higher for the specialist species, while at lower altitudes the risk may be similar for both species. Future studies should evaluate to which extent our findings result from local adaptation to parasitic pressure or from the deleterious effects of plant secondary metabolites affecting immune responses more strongly in generalist species (Schmid-Hempel, 2005; Lampert, 2012).
Despite melanization being a good indicator of parasitic resistance, it is also a costly response that depends on nutritional condition and the availability of the amino acid phenylalanine in diet (Freitak, Ots, Vanatoa, Hõrak, & Horak, 2003; Christensen, Li, Chen, & Nappi, 2005; Nappi & Christensen, 2005). Moreover, melanization response can be costly because its synthesis demands resources that can reduce fecundity or that result detrimental for the host survival (Schmid-Hempel, 2005). Therefore, a melanization response is likely to face trade-offs for its synthesis and use. Melanization can be different at high and low altitudes for three main reasons: 1) adaptations to different parasitic pressures, 2) nutrient availability to synthesize melanin or 3) abiotic conditions such as temperature regulating melanogenesis (Schmid-Hempel, 2005). Future studies need to evaluate which of these, and other environmental factors are driving insect responses to parasites at different altitudes, and if there are patterns of variation in immune responses in altitudinal gradients across insects and other taxa.
Even when lipid and muscle masses were higher at high altitudes compared to low altitudes, the ratio between lipid mass and muscle mass, which may indicate resource intake, did not differ between beetles of both species, both sexes and at both altitudes. Even though we had no clear predictions for the effect of altitude on the ratio lipid/muscle mass, these similarities indicate that there are internal mechanisms that regulate lipid storage and muscle building that do not differ between species. The quality of an animal’s diet depends on the optimal intake of protein:carbohydrate:lipid ratios that maximize individual survival and reproductive success (Simpson et al., 2015). Future studies in beetles from high and low-elevations could manipulate nutrients in the diet to evaluate whether optimal nutrient intake and storage are different at varying altitudes and how these traits vary among species with different feeding regimes.
Overall, our results indicate that altitudinal differences in tropical environments are important for shaping morphological and physiological traits in leaf-rolling beetles, including body size and immune response. If these findings are corroborated and generalized to other insects, they would become important in the face of global warming, since one potential scenario is that species from warm lowlands will migrate to higher elevations searching for cooler places (Sánchez-Guillén, Córdoba-Aguilar, Hansson, & Wellenreuther, 2016). Altitudinal migrations may imply new interactions with parasites and available food sources, requiring physiological, morphological and behavioral adjustments to deal with changing environmental conditions.

