











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
One of the greatest challenges for farmers and plant protection practitioners worldwide is the management of the whitefly Bemisia tabaci (Hemiptera: Aleyrodidae), which is not a single taxonomic entity, but a complex of many cryptic species (De Barro, Liu, Boykin, & Dinsdale, 2011) that diverge in a number of biological and genetic characteristics. These insects (named here in singular as B. tabaci, except when needed) became one of the most important pests in many tropical and subtropical regions, either as a direct pest or as a virus vector, since the early 1990s.
Intensive land cultivation, including irrigation in arid environments, has favored B. tabaci reproduction, as well as colonization of numerous cultivated and wild hosts, giving rise to their rapid spread and dominance in agricultural landscapes, causing its widespread and the maintenance of a continuous population. Among others, diseases caused by begomoviruses (genus Begomovirus, family Geminiviridae) (Zerbini et al., 2017) are certainly the most devastating. Members of the genus Begomovirus are the most damaging whitefly-borne viruses, posing a serious threat to agricultural crops throughout the World, such as those that infect cassava in Africa, cotton and okra in Asia, and tomato in the five continents (Ramesh, Sahu, Prasad, Praveen, & Pappu, 2017).
Among whiteflies, two species are considered the most predominant worldwide: B. tabaci and Trialeurodes vaporariorum. They are known as vectors of a large number of plant viruses, being the begomoviruses the main group of viruses associated with whiteflies (Navas-Castillo, Fiallo-Olivé, & Sanchez-Campos, 2011). Only species of the B. tabaci complex are able to transmit them, in a persistent manner. The process of transmission is summarized by Ghanim (2014). Most of the information on transmission is based upon the interaction of the begomovirus Tomato yellow leaf curl virus (TYLCV) and B. tabaci Middle East-Asia Minor 1 (MEAM1, formerly known as B biotype). It is believed that, in general, the interactions of distinct begomoviruses and different B. tabaci species follow a similar pattern (Rosen et al., 2015).
Begomoviruses are considered the most important viruses for many crops. In contrast, the study of transmission aspects of other viruses by B. tabaci and T. vaporariorum has received less attention. The topic of whitefly-transmitted viruses was reviewed by Navas-Castillo et al. (2011). Some aspects of viruses other than begomoviruses are summarized below.
Firstly, criniviruses (genus Crinivirus, family Closteroviridae) are transmitted in a semi-persistent manner by whiteflies. It is expected that the distribution and importance of a virus are directly related to their vector species, as exemplified in the following case. The crinivirus Tomato chlorosis virus (ToCV) is transmitted by B. tabaci and T. vaporariorum (and other Trialeurodes species), while its related species Tomato infectious chlorosis virus (TICV) is transmitted only by T. vaporariorum. ToCV is widespread and emerging worldwide, whereas TICV seems to have a limited distribution, possibly due to the extensive distribution and emergence of B. tabaci populations.
In the second place, two whitefly-transmitted carlaviruses (genus Carlavirus, family Betaflexiviridae) are transmitted in a semi-persistent manner by B. tabaci, Cowpea mild mottle virus (CPMMV) and Melon yellowing-associated virus (MYaV). CPMMV is widely spread across all continents, infecting fabaceous plants (Brunt & Kenten, 1973; Costa, Gaspar, & Vega, 1983; Antignus & Cohen, 1987), whereas MYaV has been reported only in Brazil, where it causes serious damage to melon crops (Nagata et al., 2003, 2005). Unlike other carlaviruses transmitted by aphids or for which their vector is unknown, MYaV and CPMMV are readily vectored by B. tabaci.
Thirdly, ipomoviruses (genus Ipomovirus, family Potyviridae) are also semi-persistently transmitted by B. tabaci, and have recently gained more importance because of the progressive spread of diseases caused by them (Dombrovsky, Reingold, & Antignus, 2014). These viruses share a typical genome organization of potyviruses, and because of their different vector, they were classified in a distinct genus from those transmitted by aphids, mites, or fungi.
Finally, torradoviruses (genus Torradovirus, family Secoviridae), are vectored by B. tabaci, except for Carrot torradovirus 1 (CaTV1), which is transmitted by aphids, and Lettuce necrotic leaf curl virus (LNLCV) which does not have a known vector. Members of this genus were found to be transmitted in a semi-persistent and stylet-borne manner by at least three whiteflies species (Verbeek, van Bekkum, Dullemans, & van der Vlugt, 2014).
The objective of the present paper is to provide an overview of how the interactions between different whitefly vector and virus species have been expressed so far in Costa Rica, with an emphasis on vegetables. Although specific management recommendations are not considered in this review, hopefully, the information provided will allow farmers, extension agents, and researchers involved in vegetable production, to develop more sound practical responses to current and future problems, based upon epidemiological grounds.
Emergence of begomoviruses in the world
General characteristics of begomoviruses (geminiviruses): Geminiviruses (family Geminiviridae) are small, non-enveloped plantinfecting viruses with circular single-stranded DNA (ssDNA) genomes of 2.5-5.2 kb, and a unique particle morphology, consisting of twinned (geminate) incomplete icosahedra (T=1, 22 nm x 38 nm) with a single coat protein (Zerbini et al., 2017). There are nine different genera in the family Geminiviridae, named Becurtovirus, Begomovirus, Capulovirus, Curtovirus, Eragrovirus, Grablovirus, Mastrevirus, Topucovirus, and Turncurtovirus. The delineating criteria for genus differentiation include features such as the vector, host range, or the genome organization. Geminiviruses are transmitted in nature by various types of insects. Thus, members of the genus Begomovirus are disseminated by whiteflies, while those in the genera Becurtovirus, Curtovirus, Grablovirus, Mastrevirus, and Turncurtovirus are transmitted by specific leafhoppers, the single member of the genus Topocuvirus by a treehopper, and the single member of the genus Capulavirus by an aphid. About 440 geminivirus species have been reported, being begomoviruses by far the most abundant, with more than 380 species (http://ictvonline.org/).
The genome of begomoviruses comprises either two (bipartite) DNA components (named DNA-A and DNA-B) or a single (monopartite) DNA component, each of 2.5-2.8 kb. The genome components have a typical non-coding intergenic region (IR) that contains a conserved nonanucleotide “TAATATTAC” within a stem-loop structure that includes the replication origin, essential for the initiation of rolling circle replication (Jeske, 2009). The DNA-A and a DNA-B of bipartite begomoviruses share approximately 200 bases of sequence named common region (CR) located in the IR. The nature of the genomes and their gene function has been addressed in multiple reviews (Zerbini et al., 2017). Essentially, the genome comprises six to eight proteins coded to perform all the vital functions. In bipartite begomoviruses, the DNA-A component encodes proteins associated with viral DNA replication, encapsidation, vector transmission and viral suppressors of RNA silencing (VSRs), whereas DNA-B encodes proteins involved in intercellular and intracellular movement. Monopartite begomoviruses have a more compacted genome, with a single DNA component homologous to the DNA-A of bipartite viruses, in which the movement functions are provided by the coat protein (V1) gene or the protein coded by the V2 open reading frame.
Begomoviruses do not encode a DNA polymerase but they rely on host factors recruited during the early stages of replication (Gutiérrez, 1999). Upon infection, a double stranded replicative form is generated inside the plant nucleus. Coding regions in both strands diverge from the IR, and transcription is bi-directional, using multiple overlapping transcripts for gene expression. The genome of begomoviruses evolves rapidly, and genetic exchange by recombination seems to be the major force driving their evolution (Lefeuvre & Moriones, 2015; Lima et al., 2017).
Satellites: Begomoviruses can be found associated with additional circular ssDNA components, referred to as alphasatellites or betasatellites (Briddon et al., 2012). The genome of alphasatellites (about 1.3 kb) encodes a single Rep protein and thus self-replicate, but they are dependent on the helper virus for movement, encapsidation, and vector transmission. Alphasatellites share features of origin of replication with members of the family Nanoviridae. Unlike alphasatellites, betasatellites (about 1.3-1.4 kb) cannot self-replicate but encode an important pathogenicity and symptom determinant βC1 protein, and depend completely on their helper virus for replication, encapsidation, and vector transmission functions. The families Alphasatellitidae and Tolecusatellitidae have been established to include these ssDNA satellites (Adams et al., 2017). Recently, it was also shown the association with begomoviruses of small noncoding DNA satellites that are a quarter of the size of the begomovirus genome; the name deltasatellites has been proposed for these satellites (Lozano et al., 2016).
The presence of circular ssDNA satellites co-infecting with begomoviruses in disease complexes can result in symptom modulation, and it is suggested that these interactions expand the host range of the virus. Begomoviruses effectively utilize mixed infections with DNA satellites as a mean to adapt and explore new hosts (Fiallo-Olivé, Martínez-Zubiaur, Moriones & Navas-Castillo, 2012).
Emergence of begomoviruses around the world: As indicated above, begomoviruses are transmitted in a persistent (circulative) manner by members of the B. tabaci cryptic species complex. They severely affect production of food, feed and fiber crops, threatening nutritional security worldwide, such as the significant yield losses, for instance, in cassava in Africa (Ramesh et al., 2017). Worldwide, begomoviruses are emerging as serious constraint for the production of important vegetable crops (Moriones & Navas-Castillo, 2000; Morales & Anderson 2001; Moriones, Navas-Castillo, & Díaz-Pendón, 2011). Factors underlying the success of their emergence have been reviewed in several studies (Seal, vandenBosch, & Jeger, 2006; Jones, 2009; Navas-Castillo et al., 2011; Varma, Mandal, & Singh, 2011; Lefeuvre & Moriones, 2015).
Recently, the seed-borne nature of begomoviruses has been suggested (Kil et al., 2016; Kothandaraman, Devadason, & Ganesan 2016), as well as their fruit-borne property (Delatte et al., 2003; Just, Arif, Luik, & Kvarnheden, 2017), which increases the chances for long-distance spread. Nevertheless, what has greatly contributed to the global widespread of begomoviruses as important pathogens is the introduction and emergence of its whitefly vector in tropical and subtropical areas and in protected crops of temperate regions (De Barro, 2011).
There are several examples of the successful spread of begomoviruses over long-distances in recent times, such us the global emergence of TYLCV causing damage in tomato, the rapid spread of cucurbit-infecting begomoviruses (Squash leaf curl virus, SLCuV; Watermelon chlorotic stunt virus, WmCSV; Tomato leaf curl New Delhi virus, ToLCNDV) in the Middle East and/or Mediterranean countries, or the spread of the cassava mosaic disease-associated begomoviruses in continental and insular Africa.
The tomato yellow leaf curl disease (TYLCD) severely damage tomato crops worldwide and was associated with infections of isolates of several begomovirus species and strains among them TYLCV (Moriones et al., 2011). Two of the most important begomoviruses causing TYLCD are the strains Israel (IL or type strain) and Mild (Mld) of TYLCV (TYLCV-IL and TYLCV-Mld, respectively), both with nucleotide differences that may propitiate their divergence into distinct begomovirus species in the future (Lefeuvre et al., 2010). In the particular case of TYLCV, its epidemics and its spread worldwide have been studied in detail. TYLCV is a well-documented case of the importance of international trade in the long-distance spread of begomoviruses. It is the first reported Old World (OW) monopartite begomovirus introduced into the New World (NW), and the introduction seems to have occurred through infected tomato plantlets imported from the OW in the late 1990s (Polston, Bois, Serra, & Concepción, 1994). Since then, a rapid dissemination has occurred in tomato growing regions of the Caribbean, Central, South, and North America.
Lefeuvre et al. (2010) were able to trace the routes of the ongoing global spread of the TYLCV, thanks to use of molecular epidemiology tools and the availability of a significant number of complete genome sequences of this virus, obtained from the areas most affected worldwide. They concluded that TYLCV probably originated in the Middle East during the first half of the 20th century and that global spread occurred in the 1980s, with the Mediterranean basin being the source of global TYLCV epidemics. The Western Mediterranean basin seems to have been a hub for different TYLCV-like types, with recombination among them driving their diversification (García-Andrés, Accotto, Navas-Castillo, & Moriones, 2007).
The spread of TYLCD-associated viruses to other regions of the World has occurred with multiple introductions, and afterwards the secondary spreads were documented, such as in the NW (Duffy & Holmes, 2007; Kon, Melgarejo, Almanzar, & Gilbertson, 2014; Romay, Chirinos, Geraud-Pouey, & Gillis, 2014), as well as in China and East Asia (Ueda, Kimura, Onuki, Hanada, & Iwanami, 2004; Pan et al., 2012a). Interestingly, the spread to China, Southeastern Indian Ocean and Southwestern Pacific regions seemed to have different sources (Mabvakure et al., 2016). Thus, whereas epidemics in Australia and China likely originated through multiple independent viral introductions from the East-Asian region around Japan and Korea, those in the south-western Pacific islands were caused by variants from the Western Mediterranean region.
In the Americas, TYLCD was reported primarily in the United States (USA) during 1996-1997, in Florida; it later spread to other parts of the country (Polston, McGovern, & Brown, 1999). Furthermore, there are reports of its presence in the Bahamas, Cuba, Puerto Rico, and Mexico (Sinisterra, Patte, Siewnath, & Polston, 2000; Bird, Idris, Rogan, & Brown, 2001; Salati et al., 2002; Martínez-Zubiaur, Fiallo-Olivé, Carrillo-Tripp, & Rivera-Bustamante, 2008). However, in recent years, countries such as Guatemala (Salati et al., 2010), Costa Rica (Barboza, Blanco-Meneses, Hallwass, Moriones, & Inoue-Nagata, 2014) and Venezuela (Geraud-Pouey et al., 2015) have shown evidence of the presence of TYLCD-associated virus in their tomato productions. While the strains reported for Guatemala and Costa Rica are TYLCV-IL, in Venezuela the presence of the TYLCV-Mld is confirmed.
There are no reliable estimates of losses caused nationwide by TYLCD; but, in Costa Rica for example, a report from 2014 informed it has become quite a relevant disease problem (Barboza, Blanco-Meneses, Esker, Moriones, & Inoue-Nagata, 2018). TYLCV has been found to severely affect tomato in both open-field and commercial greenhouse production systems. The symptoms observed consist of a reduction in the internodes, distortion and upward curling of leaflet margins, flower abortion, and reduction of the size of the fruits (Fig. 1).

Fig. 1 Tomato plants infected with the monopartite begomovirus Tomato yellow leaf curl virus (TYLCV) showing symptoms of curling and distortion in leaves (A) and of chlorosis and stunting of the plant (B).
Begomoviruses affecting cucurbits are another example of issues that cause losses in the OW and the NW. From the early 2000s, two bipartite begomovirus species (SLCuV and WmCSV) have emerged and are spreading in the eastern Mediterranean basin. The former virus was first reported in California in the late 1970s (Flock & Mayhew, 1981) and was the first NW bipartite begomovirus successfully invading the OW, whereas the latter is an OW begomovirus originally detected in Yemen (Jones, Sattar, & Al Kaff, 1998) and in the Arabian Peninsula (Lapidot et al., 2014). These two begomoviruses are spreading rapidly throughout Middle Eastern countries, causing severe diseases in cucurbits, in synergistic interaction in co-infected plants (Abudy et al., 2010; Sufrin-Ringwald & Lapidot, 2011). Molecular epidemiology studies conducted with SLCuV and WmCSV suggested similar emergence patterns of these two viruses in the Middle East, with no particular hurdles for the NW begomovirus when emerging in the OW (Lapidot et al., 2014).
After the long-distance spread of SLCuV from the NW to the OW, WmCSV movement between countries of the Middle East is likely occurring through movement of viruliferous whiteflies and infected plant materials. Similar to these two viruses, recent long-distance movement has occurred for the OW bipartite begomovirus ToLCNDV from the Indian subcontinent to the Western Mediterranean basin, with subsequent emergence in neighboring countries (Moriones, Praveen, & Chakraborty, 2017). Data suggest that a different strain is developing in the Mediterranean basin, which seems to be better adapted to infect cucurbit crops, causing severe yield losses (Fortes et al., 2016; Moriones et al., 2017).
Finally, another significant example of begomoviruses is that of cassava mosaic disease (CMD)-associated viruses in the African continent and nearby islands. This is a relevant case because of the importance of cassava as a staple food crop in this continent. Cassava in Africa is an example of how the introduction of an exotic plant species into foreign ecosystems provides opportunities for novel interactions, and it resulted in enormous socio-economic consequences. After the introduction of cassava from the Americas onto the African continent in the 16th to 18th centuries, this crop spread throughout tropical Africa, where today it is a major source of dietary carbohydrates for over 500 million people.
This crop is currently heavily affected by CMD, with the involvement of at least nine bipartite native African begomoviruses, collectively named cassava mosaic geminiviruses (CMGs) (Sserubombwe et al., 2008; De Bruyn et al., 2016). Such a high degree of CMGs diversity suggests that multiple native virus introduction events have occurred in cassava. Madagascar might depict a typical case of multiple introduction events and successive spread and interaction of CMGs within a relatively limited geographical area (De Bruyn et al., 2016). The rapid geographical expansion of the CMD pandemic, caused by CMG, has devastated cassava plantations in 12 countries of mainland Africa and the South West Indian Ocean islands since the late 1980s, with a continuous pattern of annual spread westward and southward along a contiguous front, for which a pivotal role is played by the vector in the pandemic (Legg et al., 2011; De Bruyn et al., 2016).
The CMG species known so far have broadly overlapping geographical ranges and frequently co-occur within mixed infections, with recombination and genome reassortment, apparently resulting into infections with increased severity (De Bruyn et al., 2016). The widespread practice of vegetative propagation of cassava seems to underlie the rapid dispersal of CMGs in tropical Africa through movement of infected plant material with subsequent spread by the vector B. tabaci.
Wild hosts (weeds) as virus reservoirs: Although it is known that wild hosts are important reservoirs of plant viruses (Duffus, 1971; Sacristan, Fraile, & García-Arenal, 2004), our knowledge about the occurrence of viruses is mostly fragmentary. The epidemiology of a virus in a given area is a complex phenomenon involving crops, virus vectors and, more importantly, wild hosts that act as vectors or virus reservoirs. The most relevant sources of plant viruses are infected plants, and an extensive host range provides viruses with greater opportunities to survive between cropping seasons (Duffus, 1971).
It is important to note that most known viruses studied in crops induce an acute infection in crop plants, whereas in wild plants viruses cause infections that are often unapparent and seem to persist with their plant hosts indefinitely (Prendeville, Ye, Morris, & Pilson, 2012; Roossinck, 2013). Moreover, recent efforts have expanded the knowledge of the true diversity of plant viruses, by studying infections in wild, undomesticated plants, showing that viruses adapt and move across the border between native wild systems and artificial, domesticated environments, contributing to disease emergence (Roossinck & García-Arenal, 2015; Stobbe & Roossinck, 2016).
In the specific case of begomoviruses, Rodelo-Urrego et al. (2013) have shown that in Mexico landscape heterogeneity is a major determinant of the infection pattern and population structure of two begomoviruses that affect Capsicum species, demonstrating the importance of wild hosts on epidemics. Wild hosts, such as Solanum nigrum or Merremia sp., have been shown to be sources of begomovirus genetic diversity for epidemics (García-Andrés, Monci, Navas-Castillo, & Moriones 2006; Idris, Al-Saleh, Zakri, & Brown, 2018). Silva et al. (2012) demonstrated the wide diversity of begomoviruses infecting leguminous weeds in Brazil, suggesting they could play a significant role in begomovirus epidemics, both as inoculum sources and as sources of emerging novel viruses. Therefore, the role of wild hosts as reservoirs and sources of infection, as well as in the evolution of virus populations, should be taken into account for a better understanding of begomovirus epidemics.
In recent times there has been an increasing concern about weeds as sources of viruses that affect crops. It is most likely that weeds commonly found within or around a crop field play a role in the epidemiology of a viral disease. Thus Barreto, Hallwass, Anquino, and Inoue-Nagata (2013) indicated that Nicandra physaloides, Euphorbia heterophylla, Sida spp. and Crotalaria spp. are potential important reservoirs of Tomato severe rugose virus (ToSRV), which affect tomato in Brazil. Furthermore, common beans (Macedo et al., 2017a) and soybeans (Macedo et al., 2017b) are susceptible to ToSRV. This indicates that we have to be particularly aware about the host range, the local cropping system, and also the agricultural landscape to understand the survival and spread of viruses in the field.
The agroecological context of Costa Rica
Biogeographical aspects: Costa Rica, a country with only 51 900 km2, is located between 8° and 11°15' N of the Equator, so it exhibits climatic characteristics typical of tropical areas, meaning that temperature, rainfall, and air humidity are very high. However, temperature is fairly stable throughout the year, which means rainfall is the driving factor for seasonality. Moreover, since its territory is part of the Mesoamerican isthmus, its narrow shape makes possible that both oceans have a moderating influence on climate, in accordance to the principle of continentality. These favorable climatic conditions strongly impinge on the biology and ecology of organisms, including those associated with agricultural settings.
In addition, from a biogeographical perspective, when in ancient times this isthmus arose and connected the two large North and South American land masses, it acted as an ecological bridge, allowing migration of organisms in both directions and favoring endemism, as well. This phenomenon helps explain why the Mesoamerican region ranks among one of the most species-rich in the World. This is an asset considered from a biological viewpoint, but in the case of both herbivorous insects and plant viruses it may become a burden because this situation implies specialists often face undescribed organisms, lacking basic information on their biology, ecology, and suitable management approaches.
Agriculture is a human activity that dates from about 10 000 years, so both insects and viruses evolved in the wilderness along with their host plants, and much later colonized agricultural fields and settled there. In the case of plants, out of some 12 000 species of vascular plants present in Costa Rica, about 1 200 are endemic ones, according to unpublished data by the National Institute of Biodiversity (INBio) (Randall García pers. comm.). Some of them may be potential host plants for either insects or viruses, or both.
Begomoviruses in vegetable cropping systems: In regard to begomoviruses, their insect vector B. tabaci has been collected from sea level to 1 700 m.a.s.l. in Costa Rica (Hilje, 2007; Hernández, Guevara, Vargas, & Ramírez, 2014), where it feeds on many wild hosts and some 15 annual crops.
In terms of their socio-economic importance, planted areas and damage levels, the major annual crops are melon, watermelon, tomato, sweet pepper, hot pepper, and common bean (Hilje, 2007); other affected vegetables are cucumber, eggplant, green bean, lettuce, pumpkin, squash, and cole crops. These crops are produced either for local consumption (tomato, sweet pepper, hot pepper, squash, lettuce, cucumber, eggplant, and cole crops) or as cash crops for export (melon and watermelon). Moreover, they can be grown along a rather wide altitudinal range, although they are mainly concentrated below 1 200 m.a.s.l., with melon and watermelon planted especially in the flatlands of Guanacaste and Puntarenas, in the Pacific basin.
From a spatial viewpoint, these vegetables can be found in a wide range of cropping systems, extending from home backyards to large monocultures, and including many small and medium-size farmers. For instance, in the year 2014 tomato (1 250 ha), peppers (1 085 ha), squash (1 098 ha) and lettuce (817 ha) were planted in 1 759, 2 365, 1 359 and 2 012 farms, respectively, yielding the following average plot sizes: 0.7, 0.46, 0.8 and 0.4 ha, respectively (INEC, 2015), which means that land tenure is very fragmented; common beans (19 470 ha in 14 707 farms) reached a value slightly higher (1.32 ha). On the contrary, there were 5 912 and 2 357 ha of melon and watermelon planted in 121 and 428 farms, giving rise to averages of 49 and 5.5 ha plot sizes, respectively. Nevertheless, data from that survey do not mention that a single melon farm owned by a foreign company reaches some 1 200 ha.
Moreover, from a temporal viewpoint, it is quite common that within a given landscape small farmers grow patches of tomato, sweet pepper, cucumber, snap bean, and common bean, interspersed among fields with perennial or semi-perennial crops, like coffee and sugarcane, thus giving rise to an agricultural "mosaic". However, vegetables represent a short-lived resource; so, once the current crop reaches physiological maturity and is not attractive anymore B. tabaci adults migrate to a new vegetable plot of any of these crops, so that such "mosaics" act as functional monocultures on a regional scale.
In the case of tomato, varieties for paste (industrial) are not planted in Costa Rica, but only table or salad tomato cultivars are used. Currently, their seedlings are sold at seedbed commercial facilities. Formerly, until the late 1990s, they used to be planted by direct seeding, but high crop losses due to begomoviruses forced farmers not to use this practice anymore.
Tomato is grown in both the Pacific and Caribbean watersheds, which are very different, as in the first one there is a sharp contrast between the dry and the rainy season, extending from December-April and May-November, respectively. In the Pacific, main rainfall peaks take place between May-June and September-October, whereas in the Caribbean, the normal pattern is one of almost permanent rainfall all year round, with short and rather unpredictable dry periods.
The Central Valley, which is a kind of plateau in the Pacific watershed, is the most important region for tomato production, where it is planted intercropped with coffee and not as a monoculture. In fact, in Grecia, Sarchí and elsewhere it is a common tradition that when coffee fields need to be renewed or pruned -so that shrubs are small-, farmers temporarily lend or rent their land to tomato producers, while benefiting from some agronomic practices (weeding, fertilization, etc.) carried out by the latter in their property.
Concerning export crops, like melon and watermelon, since their commercialization depends on temporal "windows" in the international markets, during the dry season they are planted in irrigated fields, in a staggered sequence, every two weeks. Therefore, due to food constancy, B. tabaci can reach high populations and remain active throughout the dry season, and once the last harvest takes place, adults move to alternative crops or wild plants in the surroundings, where they spend the rainy season.
As it can be readily seen, begomoviruses associated with B. tabaci are favored by such movements in both spatial and temporal terms, and even if crops are not present, wild plants ("weeds") may act as virus reservoirs. Despite the fact that some 50 wild plant species have been recorded as B. tabaci hosts in Central America, the majority being present in Costa Rica (Hilje, 1995), the Tomato yellow mottle virus (ToYMoV), the main native tomato begomovirus has not yet been detected in wild plants even though other begomoviruses like Sida golden mosaic Costa Rica virus (SiGMCRV) have been found (Rivas, Ramírez, Cubillo, & Hilje, 1995a; Höfer, Engel, Jeske, & Frischmuth, 1997; Jovel, Ramírez, Valverde, & Hilje, 1999).
Vegetable agroecosystems: From a plant protection perspective, in the real world no farm is isolated from its surroundings, and that is why the concept of agroecosystem sensuHart (1985) includes not only a given crop, but groups of crops within a certain farm or even a region. In addition, agroecosystems are composed of several interconnected subsystems: soil, crops themselves, wild plants ("weeds"), herbivorous insects, and plant pathogens.
For instance, in Central American vegetable agroecosystems include at least 182 insect pest species (Saunders, Coto, & King, 1998), which vary in importance depending on each country and particular cropping systems. In that sense, B. tabaci and begomoviruses represent only two elements of the hundreds of species belonging to the herbivorous insects and the plant pathogens subsystems. However, what farmers do to control them may seriously disturb other components of the whole vegetable agroecosystem, not to mention effects on beneficial insects (natural enemies and pollinators), as well as on aquatic ecosystems and wildlife outside the limits of such agroecosystem.
Looking at the concept of agroecosystem from another standpoint, in vegetable agroecosystems the four subsystems involved (main crops, wild plants, herbivorous insects and plant pathogens) can be conceptually linked to better analyze and understand a specific plant disease problem. This gives rise to a particular pathosystem. Therefore, whitefly-borne begomoviruses affecting a certain crop, like tomato, constitute the core of a pathosystem, along with other components, such as neighboring vegetables and other host crops spatially or temporarily connected to the main crop, as well as feral vegetation where either B. tabaci or begomoviruses can strive during food shortages due to a lack of any suitable host crop.
This approach will be used as a conceptual framework towards the end of this paper, in order to interpret and understand how the interactions of B. tabaci and its associated begomoviruses have evolved in the last 25 years in the Central Valley, the main tomato-producing area in Costa Rica.
Emergence of B. tabaci as a virus vector worldwide, and distribution in Costa Rica
Genetic diversity of B. tabaci worldwide: B. tabaci is characterized by its high genetic and biological plasticity, reproductive potential, and transmission of more than 380 plant viruses (Hilje & Morales, 2008; Navas-Castillo et al., 2011).
It is noteworthy that B. tabaci is a complex of "cryptic species", until recently known as "biotypes", which means its members are very similar morphologically, but exhibit a broad range of genetic variations and are considered sibling species. The external morphology is complex and is believed to have remained quite static since ancient times (Gill, 1990; Rosell et al., 1997; Martin, 2003; Dinsdale, Cook, Riginos, Buckley, & De Barro, 2010). With advances in molecular biology, the genetic differences observed between populations of B. tabaci have allowed to separate at least 47 B. tabaci sister clades (species) (Firdaus et al., 2013; Alemandri et al., 2015; Qin, Pan & Liu, 2016; Tay et al., 2017; Bertin et al., 2018) for which the status of "species" is proposed (De Barro et al., 2011).
Invasive species of whiteflies: B. tabaci species, MEAM1 and Mediterranean (MED, formerly known as Q biotype), are two of the most invasive species and have recently spread to a considerable number of countries in the world. They are found on a broad range of hosts, from cotton to ornamental plants, and particularly on vegetable crops (Hu et al., 2011; De Barro, 2012).
MEAM1 and MED species seem to come originally from the Mediterranean/Asia Minor/North African region (Boykin et al., 2007; Simón, Cenis, & De la Rúa, 2007; Dinsdale et al., 2010). MEAM1 has spread worldwide since the late 1980s (Bellows, Perring, Gill, & Headrick, 1994). Recently, MED has emerged in many parts of the World, displacing or co-existing with native species (Dalton, 2006; Martínez-Carrillo & Brown, 2007; Bethke, Byrne, Hodges, McKenzie, & Shatters, 2009; Grille, Gauthier, Buenahora, Basso, & Bonato, 2011; Guevara-Coto, Barboza-Vargas, Hernández-Jiménez, Hammond & Ramírez-Fonseca, 2011; da Fonseca et al., 2015). Reproductive barriers seem to exist between MEAM1 and MED, and gene flow between them is unlikely (Perring & Symmes, 2006; Elbaz, Lahav, & Morin, 2010; Sun, Xu, & Liu, 2011).
Damage caused by members of the B. tabaci complex has been reported in all continents, though in cold climates low temperatures prevent high infestations, except in protected crops (Dinsdale et al., 2010). Both MEAM1 and MED cause considerable damage to a large number of crops either through direct feeding, the transmission of plant viruses, or both (Chu et al., 2006; Dinsdale et al., 2010; De Barro et al., 2011; Hu et al., 2011; Navas-Castillo et al., 2011; Gilbertson, Batuman, Webster, & Adkins, 2015).
In America, B. tabaci New World (NW) and New World 2 (NW2) have been reported in different countries (Brown & Idris, 2005; Morales et al., 2005; Marubayashi et al., 2013; Alemandri et al., 2015). In Costa Rica, whiteflies and whitefly-borne viruses cause serious damage to many crops (Hilje, Cubillo & Segura, 1993a; Morales et al., 2005; Vargas-Ascencio et al., 2013). Previous studies showed that NW was the first whitefly species reported in Costa Rica, alone or in association with MEAM1 (Morales et al., 2005). In 2011, MED species was reported for the first time in the agricultural highlands of the Alfaro Ruiz region, in Alajuela, Costa Rica (Guevara-Coto et al., 2011).
In addition to genetic traits, the principal characteristics to differentiate the types of B. tabaci include specialization to particular hosts, host range, dispersion behavior, reproductive compatibility, resistance to different classes of insecticides, transmission of viruses to plants, and secondary endosymbionts composition (Chu et al., 2011; Pan et al., 2012b).
Biological attributes of B. tabaci as a virus vector
Among the many intrinsic biological aspects that characterize a living organism, there are five key attributes that explain why B. tabaci has been successful as a direct pest, as well as a virus vector: mouthparts and food preferences, life cycle and reproductive potential, genetic plasticity and adaptability, short and long-range movements, and host selection.
Mouthparts and food preferences: B. tabaci is a gregarious insect, and all its stages (eggs, nymphs, and adults) concentrate on the underside of leaves; nymph size ranges from 0.03-0.07 mm, whereas adults measure about 1 mm (Byrne & Bellows, 1991). Crawlers emerging from eggs barely move from the point of hatching, after which they settle for the remainder of their lives, undergoing several molts and giving rise to three additional nymphal instars, after which the adult emerges.
Both nymphs and adults have piercing-sucking mouthparts, which allow them to extract large amounts of sap from the host plant. In addition to direct damage resulting from weakening the plant as a whole, excreted honeydew drops fall on the leaves below, providing a suitable substrate for fungi (sooty molds) which then grow on these leaves, blocking sunlight and decreasing photosynthetic activity (Schuster, Stansly, & Polston, 1996). In the case of vegetables, molds also reduce the quality of produce, as it often occurs with sweet pepper in Costa Rica (Hilje & Stansly, 2018).
When feeding on its host plant, both nymphs and adults can acquire viruses if the plant is already infected, then becoming viruliferous. Even though nymphs can carry viruses inside their bodies, they are unable to disseminate them within important distances, so virus dispersal depends entirely on the adults, which are the mobile stage and also the responsible for selection and colonization of the host plant (Byrne & Bellows, 1991).
Concerning adults' mouthparts, they have seven pairs of sensillae at the tip of the labium, whose ultrastructure suggests they can act either as chemoreceptors or mechanochemoreceptors (Walker & Gordh, 1989); interestingly, tarsal sensillae are not chemoreceptors, but proprioreceptors (Walker, Perring, & Freeman, 2010). After landing on a plant, adults repeatedly rub the tip of the labium on the leaf surface, and once they decide to feed on it, insert the stylets, which are slender sucking tubes, to penetrate into the leaf tissue and reach the phloem vessels. Even though stylets are flexible enough as to move through interstitial spaces between cells, failures can often occur, as witnessed by the presence of ramified and blind salivary sheaths (Walker & Perring, 1994). The time elapsed from the beginning of the rub to when the phloem is reached can amount to 16-42 min (Walker & Perring, 1994); therefore, this is a key period to prevent virus inoculation.
Moreover, once the phloem vessels are reached, adults release saliva to dilute sap, which is rich in sugars and aminoacids, and then facilitate its ingestion. However, if they dislike any substance present inside the plant tissue or in the phloem, they are able to detect it by means of one or more of a series of nine sensillae (pegs) aligned in the cibarial chamber (Hunter, Hiebert, Webb, Polston, & Tsai, 1996), thus discriminating between suitable and unsuitable hosts.
B. tabaci adults are very polyphagous, and have been recorded feeding on at least 500 plant species in 74 families worldwide (Greathead, 1986), mainly belonging to Asteraceae, Cucurbitaceae, Euphorbiaceae, Fabaceae, Malvaceae and Solanaceae; the same trend holds for Mesoamerica (Hilje, 1995). However, its full host range remains unknown and will certainly continue increasing, due to the ability with which eventual novel species adapt to new host plants, as it occurred with MEAM1 and cole crops in different countries, in recent years (Hilje & Stansly, 2018).
This means they are endowed with a metabolic machinery (enzymes) that allows them to detoxify a vast gamut of secondary metabolites that act as chemical defenses. Interestingly enough, in Costa Rica, deterrent principles against B. tabaci have been detected in members of unusual families (Alliaceae, Chenopodiaceae, Meliaceae, Simaroubaceae and Winteraceae), as expected (Hilje, 2007). However, deterrent principles are also present in members of the most preferred families, like Asteraceae (Tithonia diversifolia), Cucurbitaceae (Momordica charantia and Sechium pittieri) and Fabaceae (Tephrosia vogelii) suggesting that the latter species contain peculiar principles that are absent in related species.
It is important to remark that B. tabaci had been observed feeding, but not always breeding, on several wild hosts and annual crops. For instance, in Costa Rica, in the recent past the predominant NW species multiplied heavily on sweet pepper but not on tomato, whilst the exact opposite occurred with the MEAM1 (Hilje & Stansly, 2018); nevertheless, when exposed to several host crops, both strongly preferred eggplant over tomato and sweet pepper.
Life cycle and reproductive potential: To be successful as a pest of vegetable crops -which represent an ephemeral resource, as their typical cropping season ranges from only 3-5 months, an insect species must be endowed with some favorable reproductive and development traits, such as a high fecundity and a short life-span. However, these traits strongly depend on two key physical factors (temperature and relative humidity), as well as on the quality and the age of the host plant.
For instance, under controlled conditions (26.5 ºC and 68 % RH) in Colombia, and reared on bean, average fecundity and generation time for B. tabaci corresponded to 75 eggs and 37-39 days (Eichelkraut & Cardona, 1989); sex ratio was 1:1. However, in Venezuela (at 25 ºC, 65 % RH, and reared on tomato), the respective figures were 194 eggs and 42 days, with a sex ratio of 2.73 females: male (Salas & Mendoza, 1995). Although differences may be due to host type, it is quite possible that the species involved were different, with the NW2 in Colombia and MEAM1 in Venezuela. This is so because by 1989 the MEAM1 species had not invaded the American continent; in addition, it is now accepted that MEAM1 normally doubles NW in fecundity (Bethke, Paine, & Nuessly, 1991). Furthermore, in the case of MED species collected in France and reared on tomato at 25 °C and 60 % RH, the respective values corresponded to 94 eggs and 22 days, with sex ratio being 1:1 (Bonato, Lurette, Vidal, & Fargues, 2007).
Concerning fecundity, its highest values (309 ± 115 eggs) have been recorded in populations exposed to sublethal quantities of insecticides, probably due to hormoligosis (Dittrich, Uk, & Ernst, 1990). This phenomenon occurs because such doses are unable to kill an insect population, but instead cause a type of stress that stimulates females to lay more eggs, while giving rise to female-biased offspring. In consequence, ironically, sublethal doses of certain insecticides may rather induce whitefly outbreaks, which in turn can favor begomovirus epidemics.
In regard to adults, they feed soon after emerging, and females mate a few hours later, normally laying less than 10 eggs a day (Eichelkraut & Cardona, 1989). Furthermore, under the conditions already stated, females and males can live for 14-19, 11-19 and 20-24 days, respectively (Eichelkraut & Cardona, 1989; Salas & Mendoza, 1995; Bonato et al., 2007). Even though laboratory conditions and constant temperatures are not representative of the real world, these figures provide an approximation of the periods during which they can act as begomovirus vectors. Females can mate repeatedly during their lifetime (Eichelkraut & Cardona, 1989), but if virgin ones do not find males, they can give rise to offspring by the phenomenon of arrhenotokic parthenogenesis (Byrne & Bellows, 1991), meaning her progeny is entirely composed by males.
Now, since B. tabaci is poikilothermic, even though the lower thermal threshold for B. tabaci corresponds to 12 ºC, optimal ambient temperatures for development range between 20-30 ºC (Gerling, Horowitz & Baumgaertner, 1986); at these values, fecundity increases and generation time shortens, leading to very rapid population growth, so that completion of its life cycle (from egg to adult) takes 27 days at 15-25 ºC, whereas at 25-35 ºC it lasts 21 days. Extrapolating these data to real world situations for vegetable cropping systems in the Pacific watershed of Costa Rica, such temperature values are easily reached during the dry season, when thermal accumulation (physiological time) is high, so that B. tabaci can fully express its reproductive potential.
This is particularly true in irrigated arid areas devoted to export crops such as melon and watermelon, like Guanacaste and Puntarenas, where furrow or drip irrigation provide enough moisture for nymphs to complete their development; on the contrary, sprinkler irrigation, which mimics rainfall, immediately decimates populations, due to dislodgement of adults from plants, as well as negative effects of relative humidity as high as 80-90 % on immature stages (Gerling et al., 1986; Hilje et al., 1993a, Castle, Henneberry & Toscano, 1996). Along the same line, commercial greenhouses there and elsewhere are ideal, since internal temperatures are quite high, rainfall is absent, and drip irrigation gives rise to satisfactory relative humidity levels for B. tabaci to accomplish development.
In summary, at the altitudinal range where vegetables are planted in Costa Rica, either in the field or in commercial greenhouses, temperature is rather high and very stable, so that B. tabaci can breed throughout the year, giving rise to very high population densities and overlapping generations (Fig. 2A and Fig. 2B), which in turn often cause devastating virus epidemics.

Fig. 2 An infestation of Bemisia tabaci adults on eggplant (A), as well as a close-up of some individuals inserting their stylets into tomato leaf (B).
Now, in addition to adverse physical factors and food scarcity, the reproductive potential of B. tabaci may be restrained by its natural enemies, of which 12 species of parasitic wasps in the genus Encarsia and two species of Eretmocerus (Aphelinidae) have been reported for Mesoamerica (Cave, 1996), as well as at least three predatory species (Chrysoperla externa, Coleomegilla maculata and Delphastus mexicanus). However, in ephemeral vegetable cropping systems, due to their slow action, parasitic are not effective to opportunely suppress whitefly populations.
In the case of B. tabaci as a vector, parasitoids have a nil effect on adults, as they are specific to nymphs, whereas predators -which normally attack both adults and nymphs- have a very narrow window to prevent plant infection, as it takes only 16-42 min for a virus to be inoculated (Walker & Perring, 1994). In other words, natural enemies cannot help preclude virus epidemics, since even if they substantially decimate nymphal stages in advance, coexisting adults can readily inoculate plants.
A particular situation where no viruses are involved takes place in some ornamental plant greenhouses in Costa Rica. Releases of two imported parasitoids (Eretmocerus mundus and E. eremicus) and the entomopathogen Beauveria bassiana, along with other management practices, allow them to complement each other if applied on the right sequence, thus substantially reducing whitefly population numbers and attaining satisfactory control (Irene Mata, pers. comm.). In protected crops, an interesting control can be achieved by means of natural enemies reducing B. tabaci population numbers, and consequently begomovirus spread.
Genetic plasticity and adaptability: B. tabaci exhibits a remarkable genetic plasticity, which is revealed by the following three biological aspects.
In the first place, immature stages can adopt different morphologies, depending on certain physical characteristics of its host plant. Thus, when developing on a plant with pubescent leaves, 4th instar nymphs show setae that confer them a hairy appearance, whilst those developing on glabrous ones lack setae and look nude (Mound, 1963). Obviously, this phenomenon is troublesome for species identification, even to expert taxonomists, but it could also affect the efficiency of its natural enemies, as the parasitoid Encarsia formosa faces difficulties in displacing through leaves with high trichome densities (Hoddle, Van Driesche, & Sanderson, 1998).
In the second place, B. tabaci can rapidly develop resistance to insecticides, including novel ones with unusual modes of action, like neonicotinoids and insect growth regulators (Palumbo, Horowitz, & Prabhaker, 2001). In the tropics, factors contributing to that include its short life cycle and continuous reproduction, favored by food availability all year long, as well as its capability to detoxify a quite diverse array of plant secondary metabolites, which provides some kind of pre-adaptation to deal with toxicants present in insecticides. This situation gets exacerbated in commercial greenhouses, where high temperatures shorten the life cycle even more, thus increasing populations, which forces growers to apply insecticides at higher doses and more frequently, in turn intensifying selection pressure.
Moreover, an important explanation for this phenomenon is rooted upon the B. tabaci breeding system, based upon haplodiploidy (Denholm, Cahill, Dennehy, & Horowitz, 1998). This means that, in addition to normal eggs laid by mated females -which are diploid-, virgin females have the ability to lay eggs too, but obviously they are haploid; the latter give rise to only males, by arrhenotoky, meaning that the offspring is composed exclusively by males. According to these authors, if a resistance gene arises by mutation in a population, regardless of its intrinsic dominance or recessiveness, it can be exposed to selection from the outset in haploid males, as they are hemizygous, then accelerating resistance development.
Finally, even though B. tabaci had been restricted to tropical and subtropical regions (Bink-Moenen & Mound, 1990), in the last three decades it has expanded its geographical range, thus reaching locations at higher latitudes and altitudes (Brown, 1993). Initially, this was explained under the assumption that B. tabaci has given rise to a series or complex of races (now species) which vary in their host-plant relationships, climatic adaptability, and so on (Brown et al., 1995). For instance, in contrast to NW species, MEAM1 is more cold-tolerant and can feed or breed on cole crops and lettuce, at the same time causing particular syndromes or physiological disorders on tomato, squash, cole crops and lettuce, which are induced by the nymphs' saliva.
Concerning the situation in Costa Rica, historically B. tabaci had been recorded only below 100 m.a.s.l. (Hilje et al., 1993b), but in recent years it has been collected at altitudes as high as 1 500 and 1 700 m.a.s.l. in Cervantes (Cartago) and Zarcero (Alajuela), respectively (Hilje, 2007; Hernández et al., 2014). It was determined they corresponded to the more cold-tolerant MEAM1 and MED species, respectively.
Short and long-range movements: B. tabaci adults are not good fliers, and tend to fly less than 2 m, especially below 50 cm from the ground (Gerling & Horowitz, 1984). Moreover, even in in-flight chamber experiments some of them are able to sustain flight for over 2 h; this is rather unusual, the average being about 15 min (Blackmer & Byrne, 1993; Byrne & Blackmer, 1996).
Therefore, for long-range movements they rely on wind currents at great heights, which can displace them to at least 7 km from their point of take-off, although they normally land less than 5 km away from such a point (Byrne & Blackmer, 1996). This occurs mainly early in the morning, in response to cues associated with skylight, so that adults are able to ascend by taking advantage of small turbulences caused by cold air drainages at ground level. Ultraviolet wavelengths emanating from the sky are the ones inducing this migratory behavior (Mound, 1962).
Interestingly enough, both in laboratory colonies and field situations, it has been shown that populations consist of migratory and trivial flying morphs (Byrne & Blackmer, 1996). In the real world, this means that individuals that belong to the trivial morph remain active all day long in a particular vegetable plot, making possible begomovirus dissemination within that plot. Despite their continuous activity, there is a flight peak in the morning, at about 10 h, as it has been recorded in Costa Rica (Jovel, Kleinn, Cartín, Valverde, & Hilje, 2000a). On the contrary, those adults belonging to the migratory morph are the ones that colonize new vegetable plots, carrying primary inoculum with them. Under controlled laboratory conditions, emigration increases as the crop reaches senescence (Blackmer & Byrne, 1993), as it has been attested in field experiments in Costa Rica, where adults readily move from old tomato plots to plots with succulent plants in their immediate vicinity (Hilje & Stansly, 2008). By means of yellow traps placed around tomato fields, in Costa Rica it has been shown that even though immigration is continuous during daylight hours, there is a clear-cut peak close to dawn, at about 18 h (Jovel et al., 2000a).
Finally, since B. tabaci adults are so dependent on wind direction, it is quite common to find them on those sides of a given crop field at which air currents are predominant. In consequence, once females lay eggs and nymphs emerge, a clear-cut aggregated or contagious within-plot distribution pattern arises. Knowing this information is essential for any sampling program. In addition, if incoming adults are viruliferous, it would be expected that virus spread will start near those sides.
Host selection: Once adults depart from an old vegetable field, carried by wind currents, there is a great deal of uncertainty about their survival. It is very likely that so many of them soon die, while others will be drifted away from agricultural fields, especially in regions with very fragmented landscapes, with no large monocultures. This is so because, even if B. tabaci adults use color as the main cue to select a host from a distance (van Lenteren & Noldus, 1990) their visual acuity is rather poor (Walker et al., 2010) and they are quite short-sighted, meaning they are unable to detect and select a given crop while flying (Byrne & Blackmer, 1996).
Migratory behavior associated with departure ought to be interrupted, as adults need to land for feeding, so there is a shift to vegetative behavior (Mound, 1962), for which other types of visual cues come into place. Then, they are attracted to greenish patches, associated with the likely presence of suitable crops.
Interestingly enough, they are not attracted to green or dark green, but mainly to deep yellow and yellow/green (Mound, 1962; van Lenteren & Noldus 1990). Such findings were experimentally corroborated in a greenhouse in Costa Rica, by exposing adults for 5 h to a gradient of seven colored sticky traps, ranging from dark green (Pantone code 341) to deep yellow (Pantone code 116); average extreme numbers were 13 and 170, respectively (Hilje, unpubl.). Moreover, in an experiment with six hues ranging from green-yellowish to deep yellow (Pantone codes 490, 526, 598, 597, 634 and 705), by exposing adults for 24 h, they attracted 111, 89, 87, 93, 124 and 93 per trap, on average, with no statistical differences between them. The biological meaning of these results is that such hues are typical of shoots and young foliage, which are rich in sugars and nitrogen, so that females prefer them to oviposit (van Lenteren & Noldus, 1990).
In addition to color, contrast between plant rows and bare soil could serve to guide adults to the crop, as has been shown in Costa Rica by planting living ground covers, including perennial peanuts (Arachis pintoi, Fabaceae), "cinquillo" (Drymaria cordata, Caryophyllaceae) and coriander (Coriandrum sativum, Apiaceae) in the furrows between tomato rows, as well as on the rows themselves (Hilje & Stansly, 2008). The ground cover could serve to efface this contrast, thus masking tomato plants. Then, without bare ground as a guide, the likelihood of visually searching adults descending on the companion ground would be a function of the area occupied by the respective cover, which is by far larger than that of the crop itself. In addition, once they land, after repeatedly tapping on the inappropriate host sensuFinch and Collier (2000), adults eventually abandon the search and exit from the tomato plot. At the end, it was shown that living ground covers significantly reduced the spread of ToYMoV, thus decreasing disease severity and increasing tomato yield (Hilje & Stansly, 2008).
Now, in order to choose a particular plant, in addition to its physical characteristics (leaf toughness, pubescence, etc.), chemical cues are quite important. In fact, ultrastructure of the antennae shows five types of sensilla, two of them (basiconic and coeloconic) very likely associated with olfactory and chemosensory functions, respectively (Walker et al., 2010).
In the search for deterrents and repellents to B. tabaci adults, a great number of substances have been tested under greenhouse conditions in Costa Rica (Hilje, 2007). For instance, tomato plants sprayed with mineral oils are significantly avoided by adults once in contact with them, suggesting that labial sensillae (Walker & Gordh, 1989), which were already discussed, act as such on earlier stages of plant colonization to discard by a tactile mechanism what is perceived as an inappropriate host.
However, in the case of hydroalcoholic plant extracts causing feeding deterrence, a different mechanism may be involved. Effective extracts, such as bitterwood (Quassia amara, Simaroubaceae), wild "tacaco" (Sechium pittieri, Cucurbitaceae), mother of cocoa (Gliricidia sepium, Fabaceae), wild sunflower (Tithonia diversifolia, Asteraceae), balsam pear (Momordica charantia, Cucurbitaceae) and fish bean (Tephrosia vogelii, Fabaceae), contain a large array of substances which seem to work in a synergistic way (Hilje, 2007). Therefore, some active principles in these extracts may be initially detected by antennal coeloconic sensillae, or later on by the sensillae (pegs) present in the cibarial chamber.
In opposition to these gustatory reactions, olfactory ones explain why volatile pure substances of botanical origin, formulated in controlled-release dispensers, repel B. tabaci adults, as it occurs with cinnamaldehyde, perialdehyde, 1-hexanal and Z-3-hexen-1-al (Hilje, 2007). In this case, thanks to antennal basiconic sensillae, adults react negatively to some olfactory cues, rejecting tomato plants protected with such substances.
Begomoviruses in mesoamerica
Main diseases in Mesoamerica: The adaptation and emergence of begomoviruses depend on a large array of factors, including their ability for genetic and biological variation through the appearance of mutants and recombinants, transport of infected plant material, introduction of species or genotypes of susceptible crops into a region with an endemic virus, vector population changes, local and regional cropping systems, and climate (Seal et al., 2006; Nawaz-ul-Rehman & Fauquet, 2009).
Studies over the past 40 years in Mesoamerica, which for the purpose and scope of this paper extends from Mexico to Costa Rica, have reported the emergence of begomoviruses in different crops. The common bean was one of the first reported crops in the region, which was affected by whitefly and begomovirus. Bean golden mosaic disease of common bean was characterized in dry bean production crops in Central, South America, and the Caribbean Basin (Morales & Anderson, 2001). It is caused by two different begomovirus Bean golden mosaic virus (BGMV) (known as BGMV type I) reported in South America (Brazil) and Bean golden yellow mosaic virus (BGYMV) (known as BGYMV type II) and reported in Central America, the Caribbean Basin and Florida (Gilbertson, Faria, Ahlquist, & Maxwell 1993; Faria et al., 1994). In the 70´s, yield losses were around 40 % to 100 %, especially in the Central American region (Gálvez & Morales, 1989). To date, bean varieties that have begomovirus resistance genes are used in the region as one of the management approaches to deal with this disease. Due to the diversity of crops planted in the region, Table 1 depicts a list of the begomoviruses reported for Mesoamerica by crop.
TABLE 1 Begomovirus species reported in Mesoamerica (from Mexico to Costa Rica) according their host crops, accepted by the International Committee on Taxonomy of Viruses (ICTV)
| Crop | Species | Acronym |
|---|---|---|
| Abelmoschus esculentus (Okra) | Okra yellow mosaic Mexico virus | OYMV |
| Abutilon permolle (Abutilon) | Abutilon golden mosaic virus | AbGMV |
| Capraria biflora | Capraria yellow spot virus | CarYSV |
| Capsicum spp. (Pepper) | Euphorbia mosaic virus | EuMV |
| Pepper golden mosaic virus | PepGMV | |
| Pepper huasteco yellow vein virus | PHYVV | |
| Carica papaya (Papaya) | Melon chlorotic leaf curl virus | MCLCuV |
| Citrullus lanatus (Watermelon) | Cucurbit leaf crumple virus | CuLCrV |
| Melon chlorotic leaf curl virus | MCLCuV | |
| Corchorus siliquosus | Corchorus yellow spot virus | CoYSV |
| Cucumis melo (Melon) | Cucurbit leaf crumple virus | CuLCrV |
| Melon chlorotic leaf curl virus | MCLCuV | |
| Cucurbita moschata (Squash) | Melon chlorotic leaf curl viru | MCLCuV |
| Desmodium spp. | Desmodium leaf distortion virus | DesLDV |
| Euphorbia spp. | Euphorbia mosaic virus | EuMV |
| Gossypium hirsutum (Cotton) | Cotton leaf crumple virus | CLCrV |
| Jacquemontia pentantha | Jacquemontia mosaic Yucatan virus | JacMYuV |
| Malva parviflora | Bean calico mosaic virus | BChV |
| Nicotiana benthamiana | Bean calico mosaic virus | BChV |
| Euphorbia mosaic virus | EuMV | |
| Pepper golden mosaic virus | PepGMV | |
| Nicotiana tabacum (Tobacco) | Bean calico mosaic virus | BChV |
| Pepper golden mosaic virus | PepGMV | |
| Pepper huasteco yellow vein virus | PHYVV | |
| Phaseolus vulgaris (Common bean) | Bean calico mosaic virus | BChV |
| Bean golden yellow mosaic virus | BGYMV | |
| Cucurbit leaf crumple virus | CuLCrV | |
| Sida golden mosaic Costa Rica virus | SiGMCRV | |
| Rhynchosia spp. | Rhynchosia golden mosaic virus | RhGMV |
| Sida sp. | Sida golden mosaic Costa Rica virus | SiGMCRV |
| Sida mosaic Sinaloa virus | SiMSiV | |
| Sida yellow mosaic Yucatan virus | SiYMYuV | |
| Sida yellow vein virus | SiYVV | |
| Solanum lycopersicum (Tomato) | Chino del tomate virus | CdTV |
| Pepper golden mosaic virus | PepGMV | |
| Pepper huasteco yellow vein virus | PHYVV | |
| Potato yellow mosaic Panama virus | PYMPV | |
| Tomato chino La Paz virus | ToChLPV | |
| Tomato golden mottle virus | ToGMoV | |
| Tomato leaf curl Sinaloa virus | ToLCSiV | |
| Tomato mosaic Havana virus | ToMHaV | |
| Tomato mottle virus-(Flo) | ToMoV-(Flo) | |
| Tomato severe leaf curl virus | ToSLCV | |
| Tomato yellow mottle virus | ToYMoV | |
| Tomato yellow leaf curl virus | TYLCV | |
| Vigna elegans | Vigna yellow mosaic virus | ViYMV |
With the exception of Tomato yellow leaf curl virus (TYLCV), the rest of the species have a bipartite genome.
Below, we prepared a review on the main aspects on begomoviruses found in cucurbits, tomato, and peppers in the region.
Begomoviruses infecting cucurbits: The major cucurbitaceous crops cultivated in Costa Rica are squash, pumpkin, melon, and watermelon. Initially, the cucurbit-infecting bipartite begomoviruses SLCuV and Squash mild leaf curl virus (SMLCuV) were reported in Southwestern USA (Brown, Idris, Alteri, & Stenger, 2002; Fauquet, Mayo, Maniloff, Desselberger, & Ball, 2005). In Arizona, Texas, North-central Mexico (Brown et al., 2000, 2002) and California (Guzmán et al., 2000), another virus, Cucurbit leaf crumple virus (CuLCrV), also known as cucurbit leaf curl virus, has been reported.
In Central America, in 2000, Melon chlorotic leaf curl virus (MCLCuV) was reported in Guatemala, being highly virulent in both melon and watermelon (Brown, Idris, Rogan, Hussein, & Palmieri, 2001; Idris, Mills-Lujan, Martin, & Brown, 2008). A putative isolate of this virus (MCLCuV-CR) was found infecting papaya (Carica papaya) and squash (C. moschata) in Costa Rica (formerly known as squash yellow mottle virus) (Karkashian, Maxwell, & Ramírez, 2002). Maliaño (2014) demonstrated that isolates of MCLCuV-CR from squash plants of Costa Rica (GenBank accession KC153490 and KC153491) infected pumpkin, squash, watermelon, and common beans. In melons, the Honeydew variety resulted resistant, whereas the Cantaloupe variety was asymptomatic; but, the virus was detected in 39 % of the samples, using PCR. Idris et al. (2008) suggested that Mesoamerica is probably the center of diversity for cucurbit begomoviruses.
Begomoviruses infecting tomato and peppers: Several bipartite begomoviruses are present in Mesoamerica, affecting tomato crops. Chino del tomate virus (CdTV) has been reported since the mid-1990s in tomato samples from Mexico (Torres-Pacheco, Garzón-Tiznado, Brown, Becerra-Flora, & Rivera-Bustamante, 1996), as well as Pepper huasteco yellow vein virus (PHYVV) and Pepper golden mosaic virus (PepGMV), infecting solanaceous crops (García-Neria & Rivera-Bustamante, 2011). PHYVV was also reported in Guatemala and El Salvador (Nakhla et al., 2005).
Tomato severe leaf curl virus (ToSLCV) and Tomato mosaic Havana virus (ToMHaV) were reported in Honduras, whereas ToSLCV, Tomato golden mottle virus (ToGMoV), ToMHaV, PepGMV, and Tomato leaf curl Sinaloa virus (ToLCSiV) were reported in Guatemala. In Belize, McLaughlin, McLaughlin, Maxwell and Roye (2008) found PepGMV and Tomato mottle virus-(Flo) (ToMoV-(Flo)) associated with tomato and sweet/hot pepper. Moreover, ToSLCV, ToLCSiV and PepGMV were observed infecting both tomato and pepper in Nicaragua (Ala-Pokeila et al., 2005). Likewise, ToYMoV and ToLCSiV are present in tomato in Costa Rica; and PepGMV in sweet pepper (Idris, Rivas-Platero, Torres-Jerez, & Brown, 1999; Nakhla et al., 2005, Barboza et al., 2018). Finally, Potato yellow mosaic Panama virus (PYMPV), ToLCSiV, and ToYMoV were reported in Panama (Engel, Fernández, Jeske, & Frischmuth, 1998; Valderrama, Velásquez, & Fernández, 2002; Herrera-Vásquez et al., 2015, 2016).
The situation aggravated after the introduction of the monopartite begomovirus TYLCV in Mesoamerica (Mexico, Guatemala and Costa Rica), USA and the Caribbean. In Costa Rica, this virus was found for the first time in 2012 at low incidence levels in tomato, in the Central Valley (Barboza et al., 2014). Since then, an increase in the incidence of plants with symptoms associated with this disease has been observed in tomato crops of the main tomato-producing areas of the country (unpublished data).
Epidemics of B. tabaci and begomoviruses in Costa Rica
A historical overview: In Costa Rica, B. tabaci had not been reported either as a direct pest or as a virus vector in vegetables before 1988. In coincidence with other Central American countries, the 1960-1970s injury records were restricted to cotton (Kraemer, 1966) and common bean (Gámez, 1971) in Guanacaste, below 100 m (Hilje et al., 1993b), as a direct pest in the first case, and as a vector of the BGYMV in the latter.
The first record of B. tabaci damage in tomato corresponds to 1988, at a rather high altitude (840 m), in Alajuela, in the Central Valley (Rosset, Meneses, Lastra, & González, 1990). Similar observations were reported in neighboring countries (Hilje, 2003) such as Nicaragua (1986), Guatemala (1987), Dominican Republic (1988), Belize, Honduras, El Salvador, Cuba and Puerto Rico (1989), Haiti (1990) and Mexico, Panama and Jamaica (1991). Even though MEAM1 was blamed for this emergence, the first report of this whitefly species in America comes from the USA territory in the early 1990s, spreading and causing severe losses in crops like cotton, melon, tomato, and ornamental plants in California, Arizona, Texas and Florida (Brown & Bird, 1992; Brown, 1994). As no whitefly analysis was available from Central America prior to 1990, this is an aspect that still needs to be clarified. Perhaps the analysis of individuals preserved in some insect collections can shed some light into this aspect, in the future.
In a survey conducted between 1990 and 1992 in Central American countries (except El Salvador and Panama) (Brown, 1993), where samples were analyzed by means of gel electrophoresis, the NW species was not detected, whereas MEAM1 was found in Guatemala (tomato and pumpkin), Belize (sweet pepper) and Nicaragua (cotton); at that time, it was suspected that MEAM1 might be present in Honduras and Costa Rica. Moreover, three unknown B. tabaci species (biotypes) were detected: C in Costa Rica (tomato and pumpkin), D in Nicaragua (tomato and pumpkin), and F in Guatemala (hosts not mentioned).
In the case of species present in Costa Rica, in 1988, it was noticed that B. tabaci was unable to reproduce on tomato plants, and even if field adult populations were quite low, they were capable of rapidly disseminating the virus which caused the mosaico amarillo del tomate (Rosset et al., 1990), today known as the ToYMoV. The same pattern was observed in successive years, with typical numbers being 0-2 nymphs and 1-6 adults per leaf, in a sharp contrast with sweet pepper, where numbers reached 50 nymphs and 80 adults per leaf (Hilje, unpublished).
Later on, in a very detailed and comprehensive survey conducted in 1998, the C species (biotype) was not detected anymore, and the overwhelmingly predominant species in Costa Rica was NW (Morales et al., 2005), present in 81 % of 43 plant samples (locations) from all provinces, including tomato, sweet pepper, hot pepper, pumpkin, cucumber, sweet potato, melon, watermelon, snap bean, and common bean. In addition, MEAM1 was detected in only eight locations, within three provinces, namely Cartago (tomato and hot pepper), Puntarenas (melon and watermelon) and Alajuela (cucumber and watermelon). The latter province was represented by Orotina, which is out of the Central Valley and very close to Puntarenas; on the contrary, MEAM1 was absent in all locations of Alajuela (Central Alajuela, Grecia, Naranjo, San Ramón and Atenas).
Interestingly enough, a few unnamed whitefly species can coexist on the same crop plant and even on the same plant, as it was noticed in Guanacaste, where individuals of the NW and MEAM1 species were collected along with other undescribed biotype (now species) on the same jalapeño pepper plants (Hilje, 2007).
So far, NW, NW2, MEAM1 and MED are the only species that have received some degree of attention in terms of understanding their life cycles, physiology, behavior, and host-plant relationships. Such studies are crucial, not just because of their academic value, but also due to their practical consequences.
Current distribution of whitefly species in Costa Rica. Both B. tabaci and T. vaporariorum have a rather widespread distribution in Costa Rica, which has been expanding over the years (Fig. 3A, Fig. 3B and Fig. 3C).
For instance, as a part of the project Sustainable Integrated Management of Whiteflies as Pests and Vectors of Plant Viruses in the Tropics, coordinated by researchers at the International Center for Tropical Agriculture (CIAT) in Colombia, a very intensive survey was carried out in 1998. It focused on agricultural regions where whiteflies were being troublesome, in order to determine which biotypes were involved. Both whitefly nymphs and adults were collected and identified, and global positioning system (GPS) coordinates were recorded, to identify "hot spots". Even though crops other than vegetables were sampled, maps herewith presented include only tomato, sweet pepper and hot pepper, which are the focus of this review. Obviously, the Central Valley is overrepresented, as it is the main area where these crops are planted.
At that time, three whitefly species were collected (Fig. 3A): the native B. tabaci NW, the aggressive and highly competitive B. tabaci MEAM1, and T. vaporariorum. Some other non-pest species were found (not shown in the map). Noteworthy, NW was the most common species in both, lowlands (Guanacaste and Limón) and mid-highlands, in the Central Valley, where T. vaporariorum is also rather common. Finally, MEAM1 was found in quite contrasting situations, like Guanacaste, at sea level, as well as in the Caribbean watershed, in Cervantes and Turrialba, at 1 500 m.a.s.l. and 600 m.a.s.l., respectively.
Thirteen years later, and for two consecutive years (2011 and 2012), Barboza (2017) sampled the main tomato-producing areas in the Central Valley, including Alajuela and Cartago locations (Fig. 3B). The species composition shifted dramatically, with the presence of the MED species, which had been detected and reported in 2009 in Zarcero, Alajuela (Guevara-Coto et al., 2011). By the time the survey was carried out, the formerly predominant B. tabaci NW had been displaced by B. tabaci MED, which had become the prevalent species, alone or coexisting with either MEAM1 or T. vaporariorum. Interestingly enough, B. tabaci MEAM1 was found in only and atypical site, coexisting with T. vaporariorum. This location corresponds to Cervantes, in Cartago, and was close to a biological reserve; this suggests that B. tabaci MED displaced B. tabaci MEAM1 from crop fields, forcing it to take refuge in wild hosts.
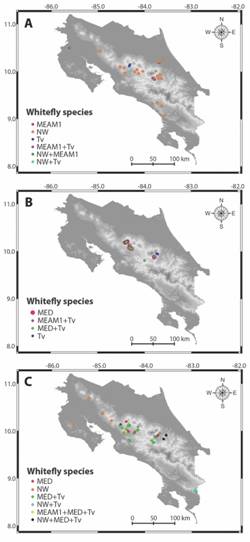
Fig. 3 Locations of Costa Rica where Bemisia tabaci and Trialeurodes vaporariorum (Tv) have been collected and analyzed for species determination over time, in tomato, sweet and hot pepper crops, in open field or greenhouse facilities: 1998 (A), 2011-2012 (B) and 2015-2016 (C). Surveys in A and C included the whole national territory, whereas the one in B was restricted to the Central Valley. Acronyms for B. tabaci species are New World (NW), Middle East-Asia Minor 1 (MEAM1), and Mediterranean (MED) and for T. vaporariorum (Tv).
A third survey was performed in 2015-2016, similar to the first one. More than 80 locations were sampled within the main tomato-producing areas across the country, and more than 600 whitefly adults were collected and analyzed. This survey revealed that MED was overwhelmingly predominant, either alone or coexisting with NW, T. vaporariorum or both (Fig. 3C). Moreover, MEAM1 was collected in only one location, along with MED and T. vaporariorum; this site corresponds to Atenas, in 2016, at 700 m.a.s.l. It is important to highlight that since the sampled area was expanded, in this opportunity it was possible to detect the presence of NW, but away from the east and west borders of the Central Valley.
In summary, these surveys show how dynamic the situation of whitefly species composition has been in Costa Rica, to the point that the initial prevalence of NW faded away, whereas MED has taken over, while the highly competitive MEAM1 has not been able to successfully colonize tomato and peppers throughout the country. In the case of T. vaporariorum, it has remained at altitudes over 1 000 m.a.s.l., mainly in the Central Valley, as well as in lowlands near the Panama border in Agua Buena, over 1 100 m.a.s.l. Of course, this represents a partial overview and a somewhat a biased picture, as the presence and economic importance of these species in other crops may be critical in particular situations.
At any rate, the causes of the observed patterns remain to be clarified. In addition to intrinsic biological traits of each whitefly species, agroecological aspects, including changes in some agronomic practices favoring one or another species, need to be studied. For example, by the time the last two surveys were carried out, vegetable production in greenhouses, fostered by government agencies, had gained considerable acceptance by farmers.
Vegetable pathosystems: Coming back to the concept of agroecosystem (Hart, 1985) and its usefulness to understand how specific pathosystems function and evolve, due to their intrinsic nature, the latter are quite dynamic and ever-changing. New B. tabaci species and new virus species or strains (including viruses in other genera, like Carlavirus, Crinivirus, Ipomovirus, and Torradovirus) may appear and establish in a certain region. Complexity can be added if new tomato and other crop varieties, as well as changes in some cultural practices (irrigation, ground covers, weed suppression, etc.), impinge upon some important characteristics of both crop and wild hosts subsystems.
Now, for an in-depth analysis and understanding of pathosystems centered on begomoviruses infecting tomato as the key crop, some factors ought to be considered. In the first place, due to the high levels of biodiversity inherent to the neotropics, there are an unknown number of endemic whitefly-borne viruses yet to be discovered and named; furthermore, sometimes they appear in mixed infections, giving rise to complex synergistic or antagonistic interactions. For instance, 19 begomovirus species had been detected in tomato until 1997 in America (Polston & Anderson, 1997), all of them native and with a rather restricted geographical range. At that time, ToYMoV was the only one found in Costa Rica.
Nevertheless, by 1998 the ToLCSiV, endemic to Mexico, was already established in Costa Rica (Idris et al., 1999), and more recently, in 2012, TYLCV, was detected in Grecia, Alajuela (Barboza et al., 2014). In addition to such tomato viruses, these authors detected PepGMV in sweet and hot pepper in Cartago, which may also be present in Alajuela. Therefore, since PepGMV can also infect tomato, as it has been shown in Nicaragua (Ala-Poikela et al., 2005), four begomoviruses could potentially coexist in the Central Valley vegetable pathosystems and elsewhere in Costa Rica, either in individual or mixed infections (TYLCV, PepGMV, TLCSiV, and ToYMoV). Or perhaps some of these viruses could eventually be displaced by the more competitive TYLCV, as has already occurred in several countries (Martínez-Zubiaur, Chang-Sidorchuk, & González-Álvarez, 2017).
In summary, the original Central Valley vegetable pathosystems driven by the B. tabaci NW species and ToYMoV have faded away over the years, becoming much more complex, as nowadays there are three species (NW, MEAM1 and MED) and four begomovirus species involved. Moreover, since the Central Valley is not an even plateau, but exhibits a rather rugged structure, with altitudes ranging from about 800 to 2 000 m.a.s.l., as well as many microclimates, on theoretical grounds it would be expected to find a great deal of ecological heterogeneity, thus favoring peculiar species-species associations between viruses and vectors in specific microhabitats where vegetables are grown. However, another scenario may occur over time, if exotic and invasive species, like TYLCV and B. tabaci MED are able to completely displace their counterparts, giving rise to very simplified vegetable pathosystems.
Secondly, each one of the aforementioned viruses does not necessarily have the same mode of interaction with its vector species, so there is the need to count upon a set of standard criteria to analyze any particular virus-vector relationship and compare it with others. Such criteria correspond to a number of critical parameters, like acquisition/inoculation periods, extrinsic and intrinsic incubation periods, and retention times for the specific virus, as well as the proportion of viruliferous whitefly adults at a particular time (Anderson, 1993). Other parameters include the rates for vector feeding, immigration, reproduction and mortality, as well as crop plant density, and the interval during which the crop is most susceptible to begomovirus injury (critical period). These parameters can be incorporated into a mathematical model, which can then reveal which are the more important variables to be intervened, as to improve decision-making procedures by farmers (Anderson, 1993).
Unfortunately, except for their diagnosis and their main host crops, epidemiological aspects of native begomoviruses are poorly known not only in Costa Rica, but in other Latin American countries. Conversely, the exotic and widely spread TYLCV has been subjected to detailed studies, which have shed light into its relationships with some of its vector species. For instance, an exhaustive review by Czosnek, Hariton-Shalev, Sobol, Gorovits, and Ghanim (2017) describes the information known to date about the above-mentioned parameters, showing that only 90 min after the whiteflies start feeding, virus particles are detected in their haemolymph (Czosnek, Ghanim, & Ghanim, 2002).
At any rate, some findings deserve to be mentioned, concerning the relationships between ToYMoV and its whitefly vector (either NW species or C biotype) in Costa Rica, which can be important for improving managing begomovirus epidemics. In regard to critical parameters, the average acquisition/inoculation period corresponds to 4 h (Bonilla, 1993), whereas the incubation (latency) period extends from 4 to 20 h (Lastra, 1993). Once in the salivary glands, virions are available to infect a new plant, and the vector can remain viruliferous for several days. However, they can acquire viruses over and over, so they can be viruliferous for their lifetime, as adult longevity typically ranges from 11-24 days (Eichelkraut & Cardona, 1989; Salas & Mendoza, 1995; Bonato et al., 2007).
Concerning the response of tomato plants to begomoviruses, when assessed by its impact on yield, tomato plants are most susceptible to ToYMoV during the first nine weeks after emergence, being this interval the critical period of infection (Acuña, 1993). Moreover, in a greenhouse experiment in Turrialba, in 15 day-old plantlets inoculated by means of viruliferous B. tabaci adults kept inside clip cages hold on leaves, it took 24 h for ToYMoV virions to get translocated all over the plant; yet, symptoms did not express until 39 days later (Rivas, Ramírez, Cubillo, & Hilje, 1995b).
These facts help explain what occurs in commercial tomato fields in Costa Rica, where it is quite common to reach 100 % of infected plants. Moreover, the course of an epidemic always follows a sigmoid pattern. The initial phase corresponds to the establishment of the epidemics, for which begomovirus inoculum should be present. However, under normal conditions a lag would be expected, as it takes some time for adults to locate and colonize a new tomato field when arriving either from neighboring crops or from wild hosts. As ToYMoV has not been found in any wild host so far (Rivas et al., 1995a; Jovel et al., 1999), primary inoculum depends entirely on other crops, probably senescent, from which B. tabaci adults fly away in search of a suitable crop. On theoretical grounds, it would be expected that all of them were viruliferous, and ready to inoculate ToYMoV virions to new tomato plants.
Based upon a set of data from researches for whitefly management purposes, Hilje (2007) summarized the following trends. In the old-fashioned system of direct seeding used in the Central Valley until the late 1990s, where tomato seedlings were exposed to the vector immediately after emerging, ToYMoV symptoms became evident 45-50 days after seeding (das), taking 25-27 additional days to infect 100 % of the plants; in contrast to findings by Rivas et al. (1995b), such a delay may be due to slow colonization by B. tabaci adults. In the case of seedlings produced in protected seedbeds for about 30 days, symptoms showed up at 15-19 days after transplant (dat), equivalent to 45-50 das, with 100 % infection being attained 30-32 days later. In other words, symptom appearance was almost constant, regardless of the planting system, suggesting that crop phenology (critical period) was the main factor that drove the epidemics, as exposure to viruliferous vectors was very different between both systems.
Manipulation of whitefly populations to appraise the impact of different proportions of viruliferous adults on tomato yields is very difficult to carry out under open-field conditions. However, in one occasion an experimental plot was seriously affected by a small senescent commercial plot in the vicinity of the experiment station, in Grecia. B. tabaci readily colonized the new plot, where the typical sigmoid pattern of the epidemics shifted to one described by a rectangular hyperbola (Fig. 4A). Moreover, the T50 value (50 % of infected plants) was reached in only 19 days, in contrast to the expected value (about 60 days) for a tomato field located far from a senescent one; also, of course, ToYMoV severity was much higher and yield much lower. Therefore, the closer old tomato fields are, the higher the rate of colonization by the vector, so that the establishment phase of the epidemics shortens and the peak of infection is attained much faster. Of course, in addition to such a high proportion of viruliferous adults (100 %), injury levels were intensified by the fact that plants (about 34 days old) were in their critical period of susceptibility.
In this particular plot, as in several others in Grecia and Turrialba, compiled and summarized by Hilje (2007), very often whitefly populations were quite low, with numbers not surpassing 10 adults when sampled in the uppermost fully-expanded leaf. This is an expression of the inability of B. tabaci NW or C biotypes to breed on tomato plants; in fact, egg and nymph numbers rarely exceed two and six per leaf, respectively (Hilje, unpublished). More astonishing is the fact that even with an average density as low as 0.3 adults/plant, 100 % of plants become infected with ToYMoV in tomato fields (Hilje, 2007) (Fig. 4B). This finding explains why, even during the rainy season, when B. tabaci population levels are very low, it is not uncommon to observe commercial tomato plots 100 % affected by ToYMoV (Hilje, 2007).
Fig. 4 Contrast between epidemics of Tomato yellow mottle virus (ToYMoV) in new plots planted close to (a) or far away from (b) old tomato fields (A), as well as numbers of Bemisia tabaci adults sampled on an upper leaf of a tomato plant (a) and incidence of ToYMoV (b), in Costa Rica (B) (After Hilje, 2007).
A possible explanation of why a density of less than one B. tabaci adult per plant can cause such epidemics is that adults keep moving all day long inside tomato fields (Jovel et al., 2000a). Moreover, by means of an original mathematical model to analyze the epidemics in Turrialba (Kleinn, Jovel, & Hilje, 1999; Jovel, Kleinn, Hilje, & Ramírez, 2000b), it was shown that the pattern of spread for ToYMoV was dependent on distance from already infected plants, and that it spread from virus foci within the tomato plots. In addition, the newly infected plants tended to appear particularly within each crop row in both directions, probably due to the fact that interplant distance was much smaller within rows (40 cm) than between rows (1.2 m), which makes foliage of neighboring plants overlap and thus protect the vector from strong wind currents.
Finally, it is important to examine the relationship between adult density, ToYMoV severity and tomato yield. In a field experiment at the Centro Agronómico Tropical de Investigación y Enseñanza (CATIE), in Turrialba, in an area with no vegetables around and surrounded by sugarcane and coffee plantations, 42-day plants were inoculated with increasing numbers (2, 5, 10, 15, 20 and 30/plant) of viruliferous adults taken from a greenhouse colony and kept inside clip cages on leaves (Salazar, Ramírez, Cubillo, Rivas, & Hilje, 1998). Severity increased as vector density did, whereas the yield was stabilized from 25 adults/plant and on, meaning that vector density affects tomato production but only up to a certain point. In other words, if vector densities exceed that figure the yield remains about the same. However, that figure may be much lower if tomato plants get inoculated at an early stage, for instance when plantlets are transplanted, at about 22-25 days. In practical terms, this means that once the vector reaches that sort of threshold figure, any investments made to control the viral disease will not improve yield altogether.
To conclude this section, it has to be stressed that in Costa Rica the preceding bioecological and epidemiological information has been quite useful for proposing integrated pest management schemes and tactics to deal with B. tabaci as a begomovirus vector in tomato, both for open-field and commercial greenhouses. Such schemes are based upon a preventative strategy centered on the critical period of tomato susceptibility to begomovirus and implemented by manipulating some components of the tomato-driven pathosystem (including crop-free periods, removal of crop residues, weed management, deployment of living barriers, trap crops, and living ground covers), plus oils as feeding deterrents. Obviously, discussion of these issues goes well beyond the scope of this paper, but it is found elsewhere (Hilje, 2001; Hilje, 2007 ; Hilje, 2014).
Concluding remarks
In Costa Rica, like in almost any tropical and subtropical country, problems with the B. tabaci-begomovirus complex were expressed more or less at the same time during the early 1990s, causing a real agricultural crisis. Almost 30 years later, now it is well known that, metaphorically-speaking, such a complex represents a Pandora's Box, given that the biotypes originally involved are actually different species, whereas new species of begomoviruses and other types of viruses have been described. At the end, these biological entities, their particular interactions and the agroecological settings where they are expressed have given rise to unique and ever-changing pathosystems which evolve continuously.
Nevertheless, Costa Rica has been fortunate in trying to face these plant protection issues, at least in comparison to the majority of Latin American countries. In the first place, the Centro de Investigación en Biología Celular y Molecular (CIBCM) existed well before the crisis started. Therefore, infrastructure and expertise in molecular methods for diagnosing insect-borne viruses and performing studies of economic importance were in place, as well as high scientific level profiting of contacts with the United States and European research groups still underway nowadays.
In the second place, in congruence with its continental mission, CATIE promptly responded to the crisis by creating the Action Plan for Whitefly and Geminivirus Management in Ibero America and the Caribbean, with the involvement of 21 Latin American countries, plus Spain and Portugal. The main objective of this Plan, which lasted for 12 years, was to validate and transfer promissory or effective technologies to farmers, in order to manage the B. tabaci-begomovirus complex. Diagnosis, research, and training activities were carried out in an interactive way.
Moreover, CATIE became involved in international networks, which allowed the Plan's participants to get continuous inputs from many institutions around the world. Such networks were the Silverleaf Whitefly: National Research, Action, and Technology Transfer, in the U.S.; the European Whitefly Studies Network (EWSN); and the project Sustainable Integrated Management of Whiteflies as Pest and Vectors of Plant Viruses in the Tropics, coordinated by CIAT (Colombia). These liaisons made possible the visit of renowned entomologists and virologists to Costa Rica, either to collect biological material or to share their expertise. In addition, some of them developed collaborative projects with CATIE and CIBCM, which had already established strong scientific liaisons, mainly through mentoring and guidance for graduate students' M.Sc. theses.
Even though such networks faded away due to several natural circumstances, long-lasting links between researchers have remained. In fact, this revision is a clear-cut evidence of this fact. Then, thanks to their particular contributions, it has been possible to count upon a comprehensive overview, with solid scientific grounds, of how the B. tabaci-begomovirus complex has expressed in Costa Rica over the years.
As a result of the intrinsic characteristics of both whiteflies and begomoviruses, as well as their unpredictable interactions, it is expected that novel plant protection problems and crises will appear sooner or later. In dealing with that, hopefully, the information herewith presented could allow researchers and extension agents to appraise the types of practical recommendations to be implemented in the field. Needless to say, such suggestions should consider the complexity of tropical agroecosystems, critical environmental issues, and the well-being of the rural inhabitants of Costa Rica and Mesoamerica.

