











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
Geographic isolation may promote differences in shape, color, and behavior between insular and mainland populations (Meiri, Dayan, & Simberloff, 2005; McClain, Durst, Boyer, & Clinton, 2013) due to particular selective pressures of the islands (Libois, Fons, & Bordenave, 1993) and/or to genetic differences (Salgueiro, Ruedi, Coelho, & Palmeirim, 2007). Therefore, the use of external features to correctly identify species on islands may be inadequate. However, traditional and geometric morphometric analyses of mammalian cranium are useful for taxonomic studies since they allow differentiating between highly similar species, particularly in bats (Sztencel-Jabłonka, Jones & Bogdanowicz, 2009; Marchán-Rivadeneira, Phillips, Strauss, Guerrero, Mancina & Baker, 2010; Jansky, 2013). On the other hand, the structure of echolocation calls is a useful tool for classifying bats since is strongly related to bat feeding and flying (Neuwieler, 1983; Russo & Jones, 2002; Hintze, Barbier, & Bernard, 2016). In particular, the high-flying habits of insectivorous bats results in low capture rates with mist nets, which usually underestimate their abundance and diversity (Hintze et al., 2016; Cormier, 2014). Therefore, echolocation calls are fundamental for species identification and are commonly used for survey and monitoring insectivorous bats (Hintze et al., 2016; Cormier, 2014). Thus, insectivorous bats from different families differ in the structure of echolocation calls, whereas frequency parameters are useful to differentiate among related species (O’Farrel & Miller, 1999). Therefore, the use of echolocation calls, additionally to morphometric analyses, may help to correctly identify populations of insectivorous bats on islands.
In Gorgona National Natural Park (NNP), a continental island, the whole assemblage of bats is listed as a conservation target; but there has been controversy about the taxonomic identity of species (i.e. Murillo-García, 2014). Thus, the taxonomic identity of the population of sac-winged bats is uncertain; it was reported as S. leptura (Alberico, 1986; Murillo, Bedoya, Velandia-Perilla, & Yusti-Muñoz, 2014) and as S. bilineata (Cadena, Gómez-Laverde, Andrade, & Peñuela, 1990; Acevedo, Beltrán, & Caicedo, 2004). These two species in mainland can be sympatric and can share shelters occasionally (Bradbury & Emmons, 1974; Bradbury & Vehrencamp, 1976; Yancey, Goetze, & Jones, 1998a; Yancey, Goetze, & Jones, 1998b). Additionally, they can feed at the same time and place without attitudinal segregation (Bradbury & Vehrencamp, 1976), which suggests potential differentiation in the preys they consume. As in other vertebrates, bats can reduce the effects of interspecific competition by decreasing niche overlapping through morphological variations; which may lead to populations with intermediate morphologies between species (Case & Sidell, 1983; Stevens & Willig, 1999). Individuals of Gorgona population share morphological traits with both species: presence of yellowish dormant lines as in S. leptura, however, those lines are strongly marked and have a dorsally darker fur with a lighter belly as in S. bilineata. Furthermore, external measures used to differentiate between them (i.e. forearm length) are intermediate between the two species. On the other hand, this population of Saccopteryx from Gorgona differs from their mainland congeners because they have diurnal activity (Cadena et al., 1990), an extremely rare behavior in Yangochiroptera (Bradbury & Emmons, 1974; Moore, 1975; Russo, Cistrone, Garonna, & Jones, 2009; Russo, Maglio, Rainho, Meyer, & Palmeirim, 2010). Due to their conservation value, identifying correctly bat species is crucial to optimize monitoring and research efforts in this protected area.
The aim of this work was to determine the taxonomic identity of the population of Saccopteryx residing in Gorgona NNP. Consequently, we evaluated the degree of morphological and acoustic similarity between Saccopteryx population from Gorgona, and S. bilineata and S. leptura from mainland. In addition, since both species of Saccopteryx differ in their mating system, with S. bilineata being polygamous and S. leptura being monogamous (reference); we assessed roosting association of Gorgona population as a surrogate for mating system. Even though individuals from Gorgona looks externally similar but slightly bigger than expected for S. leptura, we predicted that population from Gorgona NNP would be S. leptura since populations of small mammals on islands tend to be larger than in mainland (Libois et al., 1993; Lomolino, 2005).
Material and methods
Morphometric analyses: We measured a total of 62 specimens deposited in the mammal’s collection of the Universidad del Valle (UV) (Appendix I). We corroborated the identifications based on diagnostic traits proposed by Gardner (2007) and Diaz et al. (2016). For traditional morphometrics, we measured 23 skull traits (18 of cranium and 5 of the jaw, Appendix II) for S. bilineata (N = 9 individuals), S. leptura (N = 22) and Gorgona population (N = 21) using a digital caliper with 0.01 mm precision (Appendix III). For geometric morphometrics, ventral views of cranium and lateral views of jaw were digitalized for S. bilineata (7 craniums and 7 jaws), S. leptura (16 craniums and 16 jaws), and population from Gorgona (23 craniums and 19 jaws). Then, we digitized 13 landmarks for cranium and 15 for jaw (Appendix II) that were selected based on previous studies on insectivorous bats (Van Cakenberghe, Herrel & Aguirre, 2002; Sztencel-Jablonka et al., 2009; Jansky, 2013) with TpsDig v2.12 (Rohlf, 2008a) and TpsUtil v1.44 (Rohlf, 2009).
We performed Procrustes analyses to obtain shape (weighted matrix) and size variables (centroid size) of cranium and jaws with TpsRelw v1.1 (Rohlf, 2008b). Consensus configurations diagrams for craniums and jaws were plotted in CorelDRAW® X7. We also performed an analysis of repeatability to verify the consistency in locating landmarks. Then, we digitalized landmarks again for a random sample of 12 craniums and 19 jaws, and obtained repeatability values by using packages TETwin, VARwin, and MOGwin; included in CLIC program (Dujardin, 2015).
Acoustic analyses: We recorded echolocation calls of Gorgona population, and continental populations of S. leptura and S. bilineata (Appendix I) using an Echo meter touch acoustic detector (Wildlife Acoustics, 2016). For acoustic analysis, a total of 30 vocalizations, with at least 30 pulses each, were randomly selected for each population. Identify and differentiate among S. leptura and S. bilineata calls was relatively simple, since they strongly differ from each other (mainly in frequency) and from other insectivorous species and families of bats (Yancey et al., 1998a; Yancey et al., 1998b; Jung, Kalko & Von Helversen, 2007; Jakobsen, Olsen & Surlykke et al. 2015; López-Baucells, Rocha, Bobrowiec, Bernard, Palmeirim & Meyer, 2016). For each vocalization we measured minimum signal frequency (Fpmin), power-weighted mean frequency (Fpmean), peak frequency (Fppeak), average duration of echolocation pulses (Dur), average maximum frequency, (Fmax), average minimum frequency (Fmin), time weighted average frequency (Fmean), lowest absolute slope of the end of the call body (Fc), average time offset from the beginning of the call to Fc (Tc), beginning of the call body (Fk), and the average time offset from the beginning of the call to Fk (Tk). Variables were measured with software Kaleidoscope v4.1.0 (Wildlife Acoustics, 2017, Appendix IV).
We compared morphology and echolocation calls among populations by using discriminant analyses (DA). To describe cranial shape, we used the weighted matrix from geometric morphometric analyses and cranial measurements from traditional morphometric. Additionally, to describe echolocation calls we used spectral and temporal call parameters. We performed separated DAs for traditional morphometric, geometric morphometric and echolocation calls using three groups: Gorgona population, S. leptura and S. bilineata. Then, we used values of the first two discriminant functions to calculate differences among groups, based on Mahalanobis distances; calculated from centroids of Gorgona population to all individuals. Afterwards, to compare the Mahalanobis distances between groups (from Gorgona centroid to S. leptura and S. bilineata) and within groups (from Gorgona centroid to all individuals of Gorgona population), we calculated the log-ratio (Hu, Greenwood & Beyene, 2009); which is an effect size measure of differences between groups and is independent of the sample size of the groups being compared. Finally, we compared 95 % confidence intervals to estimate morphological and acoustic differentiation between groups as described in Murillo-García (2014).
Allometry assessment: To assess differences in cranium and jaw size among populations, we conducted Kruskall-Wallis tests on values of the centroid size since they were not normally distributed (KS = 1.00, P < 0.01). Additionally, we evaluated the strength and significance of allometric effects on shape using linear regressions. We used centroid size to represent cranium size and the first dimension from DA on the weighted matrix to represent cranial shape. Finally, we used coefficient of determination R2 (the square of the correlation coefficient between observed and predicted values) as a measure of the proportion of the observed shape variation that is explained by allometry. For analyses, we used the program R 3.5.0 (R Core Team, 2016), and the package MASS (Venables & Ripley, 2002) and rmeta (Lumley, 2018).
Roosting association: In order to identify the most likely mating system for Gorgona population, we identify the roosting association. Therefore, we identified and monitored roosting sites across Gorgona to establish the amount, gender and age of occupants.
Results
Traditional morphometrics: Cross-validation analyses for DA revealed a high agreement between the a priori and a posteriori classification (100 % for S. bilineata, 86 % for S. leptura, and 86 % for Saccopteryx from Gorgona). Confidence intervals for log-ratio indicated that all populations are morphologically different, but Gorgona population is more similar to S. leptura than to S. bilineata (Fig. 1A). The variable that most contributed to differentiation between S. bilineata and S. leptura was the height of the zygomatic arch (Factor load = 4.87) on the first dimension (Fig. 1B). On the other hand, maxillary canine-molar distance (Factor load = 4.47) was the variable that most contribute to differentiation between Gorgona population and both species along the second dimension (Fig. 1B).
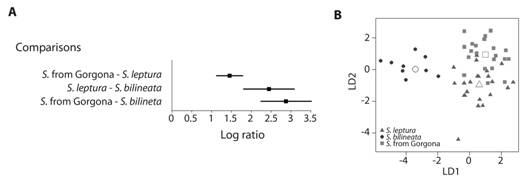
Fig. 1 Comparison of the population of Saccopteryx from Gorgona with S. leptura and S. bilineata by traditional morphometrics. A. 95 % confidence intervals for the Log ratio of between- and within-group Mahalanobis distances; and B. Scatter plot of cranial variation between Saccopteryx from Gorgona, S. leptura and S. bilineata (empty symbols represent centroids of the groups, LD1: first linear discriminant axis, LD2: second linear discriminant axis).
Geometric morphometrics: Repeatability analysis showed a 95 % of precision locating the landmarks on the cranium, and 89 % on the jaw. Cross-validation analyses for DA revealed a high agreement between the a priori and a posteriori classifications (100 % for cranium in all three populations, and, 100 % for jaw of S. bilineata, 95 % for S. leptura, and 95 % for Gorgona population). Log-ratio indicated differences in cranium and jaw morphology between populations (Fig. 2A, Fig. 2B), but Saccopteryx from Gorgona is more similar to S. leptura than to S. bilineata (Fig. 2C, Fig. 2D). This differentiation among populations for cranium and jaw was supported by consensus configurations resulting from Procrustes analysis (Fig. 2E, Fig. 2F). S. leptura has a narrower maxilla and a lower coronoid process compared with S. bilineata and Gorgona population (Fig. 2E). On the other hand, S. bilineata presents a larger maxilla, with a larger tooth row, and the back of the brain case wider than the other populations (Fig. 2F). Finally, the jaw from Gorgona population has a less pronounced curvature of the inferior border of mandibular corpus than their mainland congeners, with a different positioning of the teeth.
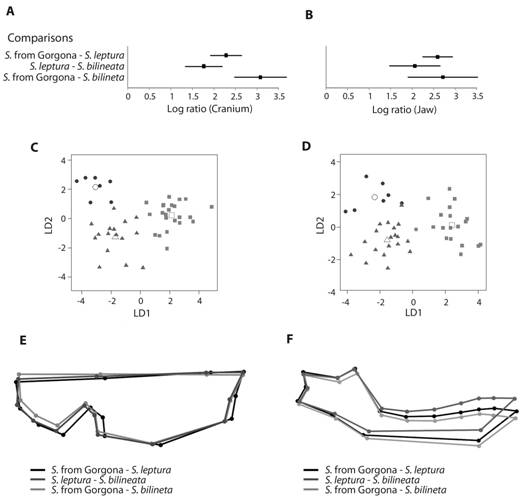
Fig. 2 Comparison of the population of Saccopteryx from Gorgona with S. leptura and S. bilineata for geometric morphometrics. 95 % confidence intervals for the Log ratio of between- and within -group Mahalanobis distances of cranium A. and jaw B.; scatter plot of cranial variation between Saccopteryx from Gorgona, S. leptura and S. bilineata for cranium C. and jaw D. (empty symbols represent centroids of the groups, LD1: first linear discriminant axis, LD2: second linear discriminant axis); and consensus configuration of landmarks for S. leptura, S. bilineata, and Saccopteryx from Gorgona NNP for cranium E. and jaw F.
Acoustic analysis: Cross-validation analyses for DA revealed a high agreement between the a priori and a posteriori classifications (100 % for S. bilineata, 86 % for S. leptura, and 83 % for Gorgona population). Log-ratio confidence intervals indicated more affinities between Gorgona population and S. leptura than with S. bilineata (Fig. 3A); which is consistent with morphological evidence. Weighted average frequency (Fmean = -2.21) was the variable that most contributed to differences between S. bilineata and S. leptura on the first dimension, and the beginning of the call body (Fk = 3.21) to differentiate between S. leptura and Gorgona population on the second dimension (Fig. 3B). Finally, echolocation calls of Saccopteryx from Gorgona population have a slightly higher frequency compared with their mainland counterparts (Fig. 4).
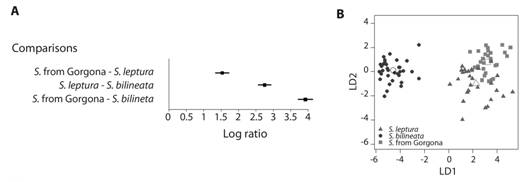
Fig. 3 A. Comparison of spectral and temporal parameters of echolocation calls of the population of Saccopteryx from Gorgona with S. leptura and S. bilineata for geometric morphometrics. A. 95 % confidence intervals for the Log ratio of between- and within-group Mahalanobis distances, and B. scatter plot of variation in echolocation calls between Saccopteryx from Gorgona, S. leptura and S. bilineata (empty symbols represent centroids of the groups, LD1: first linear discriminant axis, LD2: second linear discriminant axis).
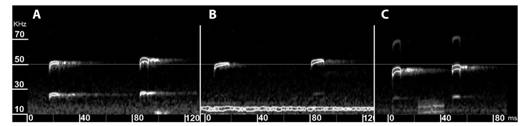
Fig. 4 Comparison of spectrograms for echolocation calls of A. Saccopteryx from Gorgona, B. S. leptura, and C. S. bilineata.
Allometry: Kruskal-Wallis test revealed significant differences in cranium (K-W = 16.29, d.f. = 2, P < 0.05) and jaw size (K-W = 12.59, d.f. = 2, P < 0.05) between the three populations. Additionally, a significant allometric relation was found for cranium (F = 0.13, R2 = 0.22, d.f. = 44, P < 0.05) and jaw (F = 0.56, R2 = 0.01, d.f. = 43, P < 0.05), indicating that 22 % of differences in conformation can be explained by variation in cranium size, and only 1 % for variations in jaw size.
Roosting association: Based in 23 shelters, we determined that the mean conformation of Saccopteryx groups was 2.47 ± 1.06 individuals where 41 % were couples, 23 % groups of two couples, 18 % a couple with a young, and 18 % solitary individuals. These results suggest that the most likely mating system for this population is monogamy.
Discusion
Gorgona population was more similar to S. leptura than to S. bilineata morphologically, vocally and reproductively. In addition, Gorgona population was bigger in size than continental populations, with a small proportion of shape differences in skull explained by size. Consequently, we conclude that Saccopteryx population from Gorgona corresponds to S. leptura. Since Gorgona Island has been isolated from mainland, at least from the last glaciation, differences with continental population of the species may be due to adaptations to environmental conditions of island, a founder effect and/or genetic drift.
Although all populations of sac-winged bats were morphologically different among themselves, population from Gorgona was more similar to S. leptura than to S. bilineata; which indicates that this population may correspond to S. leptura. Insular populations, for example, can often present considerable differences in size and/or shape with mainland populations (Libois et al., 1993, Welch, 2009; Murillo-García, 2014). Saccopteryx population of Gorgona was bigger than continental individuals, but allometric analyses showed that 22 % of variation in cranium can be explained by variations in size, whereas for the jaw just the 1 %. Gorgona is a continental island that has been connected to the continent in the past as a result of the sea level oscillation during Pleistocene glaciations, but isolated during interglacial periods (Alberico, 1986). Hence, it is considered that terrestrial populations of Gorgona have been separated from continental ones since the last glaciation, at least, 21 000 years ago (Seltzer et al., 2002). Consequently, the small founder population that colonized Gorgona has been isolated for many generations, and then genetic drift could have led to the random loss of additional alleles and fixation of others; reducing genetic diversity and increasing difference with mainland populations. Additional differentiation could have occurred through adaptations to the local environment of the island. Thus, adaptations to particular selective pressures of the island related with local environmental (i.e. temperature, humidity, luminosity) and biotic conditions (i.e. poor competitor and predator species richness) may explain differences between Gorgona and mainland populations.
Cranial variations observed might be due to differences in food resources derived from the diurnal activity of Saccopteryx on the island. Allometric changes in craniomandibular structures tend to occur as a dietary adjustment (Ravosa, Scott, McAbee, Veit, & Fling, 2015). Insectivorous bats that consume larger or hard-shelled preys tend to have big head, large gapes and tooth row, and a thicker jaw (Freeman, 1979; Fenton, 1988). Furthermore, prey texture may significantly influence craniomandibular structure (Ravosa et al., 2015; Böhmer & Böhmer, 2017). Individuals of Gorgona population have diurnal activity and they feed during the day (Vivas-Toro & Murillo-García, in prep.), which suggests a partially different diet from those in mainland since activity and density of potential insect preys changes between day and night. Compared to mainland, insular population has a higher coronoid process that can favor the closure of the jaw in lever (Van Cakenberghe et al., 2002), and a slightly larger maxilla that can be used to chew more sclerotized and/or larger preys. Besides, the increase in size of insectivorous bats can be advantageous on islands where resources are limited, because they can feed of insects of different sizes; but to take advantage of this feature, echolocation calls should modify the range of prey detection (Schnitzler & Kalko, 2001; Jung et al., 2007).
Echolocation calls also suggest that there is a greater similarity between Saccopteryx from Gorgona and S. leptura than with S. bilineata. However, calls from Gorgona population have a slightly higher frequency compared with their mainland counterparts, which suggest changes in prey consumption in this population. Echolocation frequencies usually have an inverse relation with the size of the bats, and higher frequencies are commonly used to detect small preys (Barclay & Brigham, 1994; Heller, 1995; Jung et al., 2007). This may indicate that Saccopteryx from Gorgona can potentially feed on smaller and harder insects with respect to mainland populations. Alternatively, echolocation structure can be very plastic among species, and can be modified according to habitat, flight height, and by many environmental and physiological factors (Schnitzler & Kalko, 2001; Russo & Jones, 2002). Hence, there may be significant changes in echolocation frequencies in populations of the same species that inhabits in different geographic areas (Heller & Helversen, 1989). However, echolocation calls of Emballonurids, specifically of genus Saccopteryx, tend to strongly preserve its structure and are clearly differentiated from the other species within the genus and family; making misidentifications unlikely despite where they live (Jung et al., 2007).
On the other hand, roosting association also supported the similarity between S. leptura and the population from Gorgona. The mean colony size for S. bilineata colonies is between 5.5 to 6.3 individuals /colony with majority of females, while for S. leptura the mean was between 2.6 to 4.6, usually grouped in couples; suggesting a polygynous mating system for S. bilinetata and a monogamous for S. leptura (Bradbury & Vehrencamp, 1976). Our mean value for roosts in Gorgona was 2.5, mainly conformed by couples, which suggests that individuals from Gorgona population are apparently monogamous such as S. leptura.
Identifying correctly bat species from Gorgona National Natural Park is crucial to optimize its monitoring and research since the whole assemblage is listed as a conservation target. In particular, the population of Saccopteryx is emblematic for the Park due to its singular diurnal activity. Even though population of sac-winged bats from Gorgona is different morphologically and vocally from both species, this population is more similar to S. leptura than to S. bilineata. Consequently, we conclude that Saccopteryx population from Gorgona corresponds to S. leptura based on morphology, echolocation calls and mating system. In addition, our results suggest that since Gorgona Island has been separated from continent for a long time (at least since the last glaciation), differences with continental population of the species may be due to genetic changes resulting from altered selection pressures and/or from stochastic effects of colonization on adaptive traits arising from founder effect and genetic drift.

