











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
The evaluation of the changes in patterns of bird abundance is important for our understanding of factors that could affect population dynamics (survival and mortality) and animal communities (changes in species composition) (Holt, 1993; Yodzis, 1993; Brown, 1995). In bird communities, species often adjust the timing of molting, migration and reproduction to fluctuations in food resourses, which in turn respond to predictable, seasonal climatic changes (Stiles, 1985; Loiselle & Blake, 1991; Worthington, 1996). Unpredictable climatic anomalies such as El Niño Southern Oscillation (hereafter El Niño) and presumably La Niña phenomenon, have a strong impact on survivorship and reproduction in many bird populations, and consequently affect the community structure (Grant & Grant, 1983; Stiles, 1992; Jaksic & Laso, 1999). El Niño occurs approximately every 4.1 years (Glynn, 1988), but oscillates between two and ten years (Philander, 1985). El Niño and La Niña represent extremes in sea-surface temperatures, with El Niño being above mean temperature and La Niña below it (Andreoli & Kayano, 2005).
The impact of El Niño and La Niña on bird populations often varies between taxonomic groups. Seabirds are presumably more strongly affected than land-bird species. For example, rates of survivorship and reproduction of many marine bird species (e.g., Phalacocrorax bougainvilli, Sula variegata, Pelecanus thagus, Nonnopternum harrisi, Diomedea irrorata, and Spenicus mendiculus) that nest in the area of maximum impact of El Niño are drastically reduced, particularly during strong El Niño events (Idyll, 1973; Boersma, 1978; Tovar, Guillen, & Cabrera, 1987; Rechten, 1985; Hays, 1986; Valle, Cruz, Cruz, Merlen, & Coulter, 1987; Glynn, 1988). During the strong El Niño events food supply for marine birds (e.g., the sardine Sardinops sagax and the anchovy Engraulis ringens) decrease strikingly, precluding birds to reproduce or greatly increasing mortality, particularly of juveniles (Ashmole, 1971; Hays, 1986; Anderson, 1989).
The effect of El Niño events on terrestrial bird populations in the Neotropics is difficult to evaluate for several reasons. First, occurrence, duration and intensity of El Niño are unpredictable (Cai et al., 2014). Second, the occurrence and intensity of La Niña is even less predictable (Grimm, Ferraz, & Gomes, 1998). Third, there is a lack of long-term monitoring data needed to evaluate the impact of these unpredictable climatic phenomena on the dynamics of bird populations in tropical forests (Wolfe & Ralph, 2009). During El Niño events, precipitation in general decreases in the Neotropical region, producing extensive droughts in areas where rains dominate year-round under normal conditions (Nobre & Shukla, 1996; Andreoli & Kayano, 2005; Paxton, Cohen, Paxton, Németh, & Moore, 2014), although precipitation at more localized scales could be higher in some areas during El Niño years (Villalobos & Retana, 2000). On the contrary, La Niña which sometimes occurs right after El Niño or not at all is often considered the anti-Niño because it has roughly the opposite effect on climatic conditions (Philander, 1985; Brenes & Bonilla, 2012). Our main objective here is then to test the effect of El Niño and La Niña events on the abundance of terrestrial bird populations in three different forest types in Costa Rica. We used data from Christmas Bird Counts from 1984 to 2011, the longest data set of bird abundance in Costa Rica. The effect of the El Niño is extremely variable over Central America (Ropelewski & Halpert, 1987; Enfield & Mayer, 1997; Sheffield & Wood, 2008). For instance, Waylen, & Caviedes (1996) and Giannini, Kushnir, & Cane (2000) show that El Niño events in Costa Rica produce contrasting climatic conditions between Pacific (decreases rainfall) and Caribbean slopes (increases rainfall). Thereby, we expect contrasting results on bird abundance among forest types since the effect of El Niño could affect differently each forest.
Materials and methods
Study sites: We selected three localities for which long term monitoring data (≥ 16 years) on bird abundance and species composition is available. This data set allows comparisons among multiple events of El Niño, La Niña, and neutral years. The three localities include three forest types: lowland rain forest (La Selva Biological Station, Caribbean slope), premontane forest (Grecia, Pacific slope), and montane forest (Monteverde, continental divide that includes portions of both Caribbean and Pacific slopes). Forest vegetation for La Selva and Monteverde were described by Hartshorn, (1983) and Haber, (2000) respectively. Vegetation in Grecia consists of a matrix of large sugar cane and coffee plantations, urban habitats and large fragments of riparian premontane forest.
Bird abundance data: We obtained bird species abundance from Christmas’ Bird Counts (CBC) conducted in each locality under Audubon protocol (National Audubon Society, 2010). CBCs use a standard protocol that consists of counting bird species and abundance inside a circle of 24 km in diameter (452.4 km2 area). In each locality, the circle includes the largest area of most forest types in that locality; the location of the circular sampling area is set prior to the first CBC and this location is maintained through time. Additionally, CBCs are conducted during a single 24 h period started at 00:00 h and finished at 24:00 h on the same routes year after year, which increases the comparative power of the data over time. The period of time analyzed varied across localities: lowland rain forest from 1985 to 2011, premontane forest from 1984 to 2003, and montane forest from 1994 to 2010. Data on CBC were obtained from The Organization for Tropical Studies (OTS, 2010) (http://www.ots.ac.cr/cbc_laselva) and National Audubon Society (http://www.christmasbirdcount.org).
For each locality, we selected resident large frugivorous (> 30 g), medium size frugivorous (20 - 30 g), small frugivorous (< 20 g), and nectarivorous bird species (Table S1). We selected this subset of species because they fulfilled the following criteria: (1) species are conspicuous reducing their false-positive identification, and (2) species main food resource (fruits and nectar) are seasonal and influenced by rainfall levels that change according to El Niño and La Niña occurrence. We only used species recorded in 50 % or more of the CBC in each locality.
Statistical analyses: We compared bird abundance (e.g. count frequency) of species included in each of the four-bird diet-categories for each of the forest types and three weather events (i.e., El Niño, La Niña, neutral years) using Generalized Linear Models with a Poisson distribution or quasipoisson distribution when residuals were over-dispersed (GLM, library MASS). We used Zero Inflated Models (library glmmADMB) with a negative binomial probability distribution when the number of ceros exceed those expected by a Poisson (or quasipoisson) distribution. With the negative binomial probability distribution, we could accomplish the assumptions of normality of residuals and homoscedasticity. El Niño, La Niña, or neutral years were defined with the Oceanic Niño Index (ONI), obtained from https://ggweather.com/enso/oni.htm. Events are defined by the National Oceanic and Atmospheric Administration (https://www.noaa.gov/) as five consecutive overlapping 3-month periods with values of the ONI index above the +0.5o anomaly for warm (El Niño) events, or below the -0.5º anomaly for cold (La Niña) events. For the analyses we included the median value (negative or positive) of the 3-month periods for each year defined as El Niño, La Niña, or neutral. Thus, the statistical models included bird-diet categories, weather events, the co-variable “ONI”, which account for the variation in climatic conditions during of El Niño, La Niña, and neutral years, and the total number of person-hours per bird count as another covariate in the analyses (i.e., time) to account for yearly-variation in effort and total distance covered, and species as random factors within the model. All analyses were done using the R statistical language, version 3.0.1 (R Core Team, 2013).
Results
The number of El Niño, La Niña, and Neutral years were similar for all three study sites. Lowland rain forest included seven years in each of the three weather categories; the premontane forest included six El Niño and La Niña years, and seven neutral years; and the montane forest included six El Niño years, seven La Niña years, and five neutral years.
Lowland rain forest: We tested the effect of weather events on the abundance of 17 large frugivorous (LF), 27 medium size frugivorous (MF), 36 small frugivorous (SF), and 12 nectarivorous (Nc) bird species in the lowland forest (Table S1). The abundance of large and medium size frugivores increased significantly during La Niña events in comparison to neutral years (Table 1, Fig. 1), and the abundance of Nc during the El Niño events relative to neutral years. Abundance of SF was not affected by La Niña or El Niño events; nor El Niño affected significantly the abundance in any of the three groups of frugivores (Table 1). Neither we found significant differences in the abundance of frugivores and nectarivores between La Niña and El Niño events (Table 1). The ONI did not have a significant effect on frugivorous and nectarivorous birds in the lowland rain forest (Table 1).
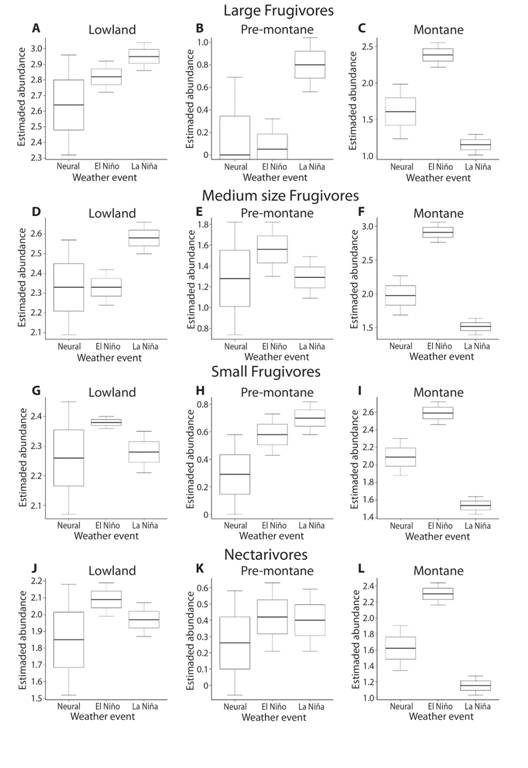
Fig. 1 Mean, standard error, and confidence intervals for three groups frugivorous birds (large, medium size, and small), and nectarivorous birds in three forest types (lowland, pre-montane, and montane). The mean was estimated from the intercept of the statistical models, which directly correlate with the mean values of the variables included in the models.
TABLE 1 Comparison between climatic events (El Niño, La Niña, and Neutral years) for Large frugivores, Medium size frugivores, Small frugivores, and Nectarivores in three forest types based on Generalized Linear Models with a Poisson probability distribution.
| Lowland rain forest | ||||
|---|---|---|---|---|
| Large frugivores | ||||
| Effect | Coefficient | SE | T | P |
| Neutral | 2.64 | 0.32 | 8.13 (446) | 0.0000 |
| Niño - Neutral | 0.18 | 0.10 | 1.81 (446) | 0.0708 |
| Niña - Neutral | 0.31 | 0.09 | 3.27(446) | 0.0011 |
| Niña - Niño | 01.2 | 0.16 | 0.79 (446) | 0.4274 |
| ONI | 0.05 | 0.05 | 0.92 (446) | 0.3532 |
| Counting hours | 0.00 | 0.00 | 8.79 (446) | 0.0000 |
| Medium frugivores | ||||
| Effect | Coefficient | SE | T | P |
| Neutral | 2.33 | 0.24 | 9.55 (696) | < 0.0001 |
| Niño - Neutral | 0.00 | 0.09 | 0.40 (696) | 0.9681 |
| Niña - Neutral | 0.25 | 0.08 | 2.99 (696) | 0.0029 |
| Niña - Niño | 0.25 | 0.14 | 1.75 (696) | 0.0806 |
| ONI | 0.09 | 0.05 | 1.90 (696) | 0.0581 |
| Counting hours | 0.00 | 0.00 | 8.25(696) | < 0.0001 |
| Small frugivores | ||||
| Effect | Coefficient | SE | T | P |
| Neutral | 2.26 | 0.19 | 11.57 (896) | < 0.0001 |
| Niño - Neutral | 0.12 | 0.02 | 1.60 (896) | 0.1089 |
| Niña - Neutral | 0.02 | 0.07 | 0.23 (896) | 0.7881 |
| Niña - Niño | -0.10 | 0.11 | -0.85 (896) | 0.3928 |
| ONI | -0.01 | 0.04 | -0.19 (896) | 0.8428 |
| Counting hours | 0.00 | 0.00 | 11.33 (896) | < 0.0001 |
| Nectarivores | ||||
| Effect | Coefficient | SE | T | P |
| Neutral | 1.85 | 0.33 | 5.11 (321) | < 0.0001 |
| Niño - Neutral | 0.24 | 0.10 | 2.33 (321) | 0.0203 |
| Niña - Neutral | 0.12 | 0.10 | 1.19 (321) | 0.2352 |
| Niña - Niño | -0.12 | 0.16 | -0.76 (321) | 0.4491 |
| ONI | 0.00 | 0.06 | 0.08 (321) | 0.9382 |
| Counting hours | 0.00 | 0.00 | 5.66 (321) | < 0.0001 |
| Pre-montane forest | ||||
| Large frugivores | ||||
| Effect | Coefficient | SE | T | P |
| Neutral | 0.00 | 0.69 | 0.00 (104) | 0.9968 |
| Niño - Neutral | 0.05 | 0.27 | 0.19 (104) | 0.8485 |
| Niña - Neutral | 0.80 | 0.24 | 3.27 (104) | 0.0015 |
| Niña - Niño | 0.74 | 0.44 | 1.70 (104) | 0.0913 |
| ONI | 0.22 | 0.14 | 1.62 (104) | 0.1069 |
| Counting hours | 0.03 | 0.00 | 6.00 (104) | < 0.0001 |
| Medium frugivores | ||||
| Effect | Coefficient | SE | Z | P |
| Neutral | 1.28 | 0.54 | 2.40 (228) | 0.017 |
| Niño - Neutral | 0.28 | 0.26 | 1.10 (228) | 0.272 |
| Niña - Neutral | 0.01 | 0.20 | 0.07 (228) | 0.942 |
| Niña - Niño | -0.27 | 0.40 | -0.67 (228) | 0.504 |
| ONI | -0.07 | 0.13 | -0.49 (228) | 0.623 |
| Counting hours | 0.02 | 0.00 | 4.92 (228) | < 0.001 |
| Small frugivores | ||||
| Effect | Coefficient | SE | T (339) | P |
| Neutral | 0.29 | 0.29 | 0.98 (338) | 0.3254 |
| Niño - Neutral | 0.29 | 0.15 | 1.99 (338) | 0.0469 |
| Niña - Neutral | 0.41 | 0.12 | 3.29 (338) | 0.0011 |
| Niña - Niño | -0.29 | 0.15 | -1.99 (338) | 0.5462 |
| ONI | 0.10 | 0.07 | 1.49 (338) | 0.1370 |
| Counting hours | 0.02 | 0.00 | 5.96 (338) | < 0.0001 |
| Nectarivores | ||||
| Effect | Coefficient | SE | T | P |
| Neutral | 0.26 | 0.32 | 0.82 (266) | 0.4147 |
| Niño - Neutral | 0.16 | 0.21 | 0.75 (266) | 0.4522 |
| Niña - Neutral | 0.14 | 0.19 | 0.72 (266) | 0.4680 |
| Niña - Niño | -0.02 | 0.35 | -0.05 (266) | 0.9611 |
| ONI | 0.13 | 0.11 | 1.21 (266) | 0.2244 |
| Counting hours | 0.02 | 0.00 | 4.99 (266) | < 0.0001 |
| Montane forest | ||||
| Large frugivores | ||||
| Effect | Coefficient | SE | T | P |
| Neutral | 1.61 | 0.37 | 4.35 (268) | < 0.0001 |
| Niño - Neutral | 0.77 | 0.17 | 4.42 (268) | < 0.0001 |
| Niña - Neutral | -0.45 | 0.14 | -3.20 (268) | 0.0016 |
| Niña - Niño | -1.23 | 0.27 | -4.65 (268) | < 0.0001 |
| ONI | -0.42 | 0.09 | -4.39 (268) | < 0.0001 |
| Counting hours | 0.00 | 0.00 | 4.22 (268) | < 0.0001 |
| Medium frugivores | ||||
| Effect | Coefficient | SE | T | P |
| Neutral | 1.98 | 0.29 | 6.73 (506) | < 0.0001 |
| Niño - Neutral | 0.93 | 0.15 | 5.98 (506) | < 0.0001 |
| Niña - Neutral | -0.46 | 0.12 | -3.81 (506) | 0.0002 |
| Niña - Niño | -1.39 | 0.23 | -6.14 (506) | < 0.0001 |
| ONI | -0.52 | 0.08 | -6.15 (506) | < 0.0001 |
| Counting hours | 0.00 | 0.00 | 2.78 (506) | 0.0055 |
| Small frugivores | ||||
| Effect | Coefficient | SE | T | P |
| Neutral | 2.09 | 0.21 | 10.06 (829) | < 0.0001 |
| Niño - Neutral | 0.50 | 0.13 | 3.77 (829) | < 0.0001 |
| Niña - Neutral | -0.55 | 0.10 | -5.24(829) | < 0.0001 |
| Niña - Niño | -1.05 | 0.13 | -3.77 (829) | < 0.0001 |
| ONI | -0.30 | 0.07 | -4.18 (829) | < 0.0001 |
| Counting hours | 0.00 | 0.00 | 4.69 (8.29) | < 0.0001 |
| Nectarivores | ||||
| Effect | Coefficient | SE | Z | P |
| Neutral | 1.62 | 0.28 | 5.87 (468) | < 0.0001 |
| Niño - Neutral | 0.68 | 0.14 | 4.66 (468) | < 0.0001 |
| Niña - Neutral | -0.47 | 0.12 | -4.14 (468) | < 0.0001 |
| Niña - Niño | -1.15 | 0.21 | -5.57 (468) | < 0.0001 |
| ONI | -0.41 | 0.08 | -5.35 (468) | < 0.0001 |
| Counting hours | 0.00 | 0.00 | 3.06 (468) | < 0.0001 |
The intensity of the climatic event (ONI Index) was included in the model.
The corresponding degrees of freedom are included in parentheses besides the T-values.
The Z-values correspond to the Zero Inflated Models.
Pre-montane forest: For the premontane forest, we analyzed the abundance of 6 LF, 12 MF, 19 SF, and 15 Nc bird species. In this forest, the abundance of LF and SF increased significantly during La Niña events when compared with neutral years (Table 1, Fig. 1), but it did not affect the abundance of MF nor Nc. The abundance of SF also increased during El Niño, but it did not affect the abundance of other groups. The ONI had no effect on abundance of any of the bird groups (Table 1).
Montane forest: For the montane forest, we analyzed the abundance of 17 LF, 30 MF, 49 SF, and 26 Nc bird species. In this forest the abundance of all three groups of frugivores and the nectarivores increased during El Niño events in comparison to neutral years, but decreased for all bird groups during La Niña events (Table 1, Fig. 1). The abundance of all four groups correlated negatively with ONI index values (Table 1). Thus, the abundance of birds in the montane forests has a strong increment during El Niño events, despite that bird abundance decreased as the index ONI increased.
The total number of person-hours per bird count correlated positively with bird abundance in all cases. However, the ONI index had only a significant effect on bird abundance in the Montane forest.
Discussion
The variation in the abundance of frugivorous and nectarivorous birds detected in this study could be the result of temporal and spatial changes in the phenology of flowering and fruiting species (food resources) caused by El Niño and La Niña events. The shifting in the pattern of precipitation, intensity of rainfall, and radiation caused by El Niño and La Niña directly affect the periodicity of phenological phases, the distribution, and mortality of plants (Wright, Carrasco, Calderón, & Paton, 1999; Villalobos & Retana, 2000; Holmgren, Scheffer, Ezcurra, Gutiérrez, & Mohren, 2001; Jaksic, 2001; McPhaden, Zebiak, & Glantz, 2006; Chen & Cane, 2008). For instance, flowering and fruiting of understory plant species correlate with increasing temperature and radiation (Kimura, Yumoto, Kikuzawa, & Kitayama, 2009). Increasing variation of food resources (Wright et al., 1999) could force birds to change their movement patterns or to move to areas where the effect of these climatic anomalies is milder. Such variation could also increase mortality of birds if food resources decrease drastically (Wolfe & Ralph, 2009; Boyle, Norris, & Guglielmo, 2010), and all these factors could affect the abundance of species locally (Jaksic & Laso, 1999).
The fluctuation in bird abundance across bird groups in this study is similar to that reported for other terrestrial ecosystems in the Neotropics. For instance, in Barro Colorado island (Panama) fruit production greatly increases during El Niño events but decreases drastically when a mild dry season (e.g., the Niña event) follows El Niño, thus increasing mortality of frugivorous mammals (Foster, 1982; Wright et al., 1999). In the Galapagos Islands, reproduction and recruitment of Geospiza fortis significantly increased as a result of high production of seeds during the strong rainy season of El Niño 1982-1983 (Grant, 1986). In Chile, the diversity and density of several groups of landbirds increased during El Niño 1997-1998 (Jaksic & Laso, 1999). These changes in diversity and density are presumably a direct consequence of increasing rainfall in the region during El Niño years (Jaksic & Laso, 1999).
The effect of El Niño on climate varies among events and geographical scales. In Chile, total rainfall increases during El Niño but in Barro Colorado Island, rainfall actually decreases during these events, although increases in food resources have in both cases been attributed to El Niño events (Wright et al., 1999; Jaksic, 2001). In Costa Rica, precipitation during the El Niño and La Niña events also varies between slopes and across events (Waylen, Quesada, & Caviedes, 1994; Waylen, & Caviedes, 1996; Waylen, Caviedes, Poveda, Mesa, & Quesada, 1998; Brenes & Bonilla, 2012). This variation affects differently the abundance of the bird groups in different forests. In the lowland forest abundance of frugivores (large and medium size) increased during La Niña years, but El Niño years did not have a significant effect on frugivores abundance, though abundance of nectarivores increased during these events (Table 1, Fig. 1). The montane forest showed an entirely opposite pattern for frugivores. In this forest, the abundance increased consistently for all frugivorous and nectarivorous birds during El Niño events, but their abundance decreased during La Niña. It is notable in the montane forest that El Niño has a strong positive effect on bird abundance, but this abundance tends to decrease if intensity of El Niño increases (Table 1). This suggests that there are at least two different processes affecting bird abundance in montane forests. One that likely makes birds to move uphill during El Niño, and another, perhaps less intense, affecting negatively their abundance at high elevation. In the premontane forest these climatic events had little effect on bird abundance. Only abundance of SF increased during La Niña events, but had no effect on LF, MF, and Nc (Table 1, Fig. 1). It is likely that differences in climatic conditions at small geographical scales affects differently plant phenology, and so avian food resources in each forest type, but until we have detailed information on the climatic conditions along the mountain slopes and their effect on bird food resources, the response to fluctuation in frugivores and nectarivores will remain elusive (Williams-Linera & Meave, 2002).
The changes in abundance during El Niño and La Niña events could be attributed to temporal and spatial fluctuation in food resources, but these climatic events may also affect bird reproduction and survivorship (Wolfe, Ralph, & Elizondo, 2015). Both climatic events may have drastic negative effects at the level of population and community structure. For instance, some populations of Darwin’s finches (Geospiza spp.) fail to reproduce during extremely dry years, which usually coincide with La Niña events, and mortality increases up to 85 % during these years and sex ratio skewed toward males (Boag & Grant, 1981; Boag & Grant, 1984; Grant & Grant, 1983; Grant, 1986). These factors delayed the recovery of populations (Boag & Grant, 1984). Similarly, survivorship of the Long-billed Hermit (Phaetornis longirostris) drastically decreased in a Costa Rican rain forest as a consequence of a flower shortage caused by a severe drought during El Niño event 1972-1973 (Stiles, 1992). Recovery of this population lasted about four years. Bird survival may also depend on the specific habitat features. For instance, in Costa Rica Survivorship of the White-collared Manakin (Manacus candei) was drastically affected by el Niño in disturbed forests but not in near mature forests (Wolfe et al., 2015).
Climate scenario models predict that El Niño events will increase in frequency as global, greenhouse warming increases (Cai et al., 2014). Considering that some populations of terrestrial birds require nearly four years to recover after the impact caused the by El Niño and that the mean period of El Niño occurrence is 4.1 yrs (Glynn, 1988), an increase in frequency of these events would increase the probability of some avian populations to become extinct if they are not capable to recover in a shorter time. We finally highlight the importance of long-term data on bird abundance and phenology of plants to evaluate more precisely the effects of the El Niño and La Niña climatic anomalies on population’s dynamics and community structure of tropical birds.
Ethical statement: authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.
Supporting Information
TABLE S1 Bird species used in the analysis of effect of El Niño and La Niña in three forests of Costa Rica according to the effects of El Niño and La Niña
| English name | Category | Size (cm) | Forest type | |||
| Lowland rainforest | Premontane forest | Monatane forest | ||||
| TINAMIFORMES | ||||||
| Tinamidae | ||||||
| Nothocercus bonapartei | Highland Tinamou | Large frugivouros | 38 | X | ||
| Tinamus major | Great Tinamou | Large frugivouros | 43 | X | X | |
| Crypturellus soui | Little Tinamou | Medium frugivouros | 23 | X | ||
| Crypturellus boucardi | Slaty-breasted Tinamou | Large frugivouros | 28 | X | ||
| GALLIFORMES | ||||||
| Cracidae | ||||||
| Ortalis cinereiceps | Gray-headed Chachalaca | Large frugivouros | 51 | X | X | |
| Penelope purpurascens | Crested Guan | Large frugivouros | 86 | X | X | |
| Chamaepetes unicolor | Black Guan | Large frugivouros | 64 | X | X | |
| Crax rubra | Great Curassow | Large frugivouros | 91 | X | ||
| Odontophoridae | ||||||
| Odontophorus leucolaemus | Black-breasted Wood-Quail | Medium frugivouros | 23 | X | ||
| COLUMBIFORMES | ||||||
| Columbidae | ||||||
| Patagioenas cayennensis | Pale-vented Pigeon | Large frugivouros | 30 | X | ||
| Patagioenas speciosa | Scaled Pigeon | Large frugivouros | 32 | X | ||
| Patagioenas flavirostris | Red-billed Pigeon | Large frugivouros | 30 | X | X | |
| Patagioenas fasciata | Band-tailed Pigeon | Large frugivouros | 35 | X | X | |
| Patagioenas subvinacea | Ruddy Pigeon | Medium frugivouros | 28 | X | X | |
| Patagioenas nigrirostris | Short-billed Pigeon | Medium frugivouros | 26.5 | X | X | |
| Columbina inca | Inca Dove | Medium frugivouros | 20 | X | ||
| Columbina passerina | Common Ground-Dove | Small frugivorous | 16 | X | X | |
| Columbina talpacoti | Ruddy Ground-Dove | Small frugivorous | 16.5 | X | ||
| Claravis pretiosa | Blue Ground-Dove | Medium frugivouros | 20 | X | ||
| Leptotila verreauxi | White-tipped Dove | Medium frugivouros | 26 | X | X | X |
| Leptotila cassinii | Gray-chested Dove | Medium frugivouros | 24 | X | ||
| Geotrygon veraguensis | Olive-backed Quail-Dove | Medium frugivouros | 21.5 | X | X | |
| Geotrygon chiriquensis | Chiriqui Quail-Dove | Medium frugivouros | 30 | X | ||
| Geotrygon lawrencii | Purplish-backed Quail-Dove | Medium frugivouros | 25 | X | ||
| Geotrygon costaricensis | Buff-fronted Quail-Dove | Medium frugivouros | 28 | X | ||
| Geotrygon montana | Ruddy Quail-Dove | Medium frugivouros | 23 | X | ||
| APODIFORMES | ||||||
| Trochilidae | ||||||
| Glaucis aeneus | Bronzy Hermit | Nectarivorous | X | |||
| Threnetes ruckeri | Band-tailed Barbthroat | Nectarivorous | X | |||
| Phaethornis guy | Green Hermit | Nectarivorous | X | |||
| Phaethornis longirostris | Long-billed Hermit | Nectarivorous | X | X | ||
| Phaethornis striigularis | Stripe-throated Hermit | Nectarivorous | X | X | ||
| Doryfera ludovicae | Green-fronted Lancebill | Nectarivorous | X | |||
| Colibri delphinae | Brown Violetear | Nectarivorous | X | |||
| Colibri cyanotus | Lesser Violetear | Nectarivorous | X | X | ||
| Heliothryx barroti | Purple-crowned Fairy | Nectarivorous | X | X | ||
| Anthracothorax prevostii | Green-breasted Mango | Nectarivorous | X | X | X | |
| Heliodoxa jacula | Green-crowned Brilliant | Nectarivorous | X | |||
| Panterpe insignis | Fiery-throated Hummingbird | Nectarivorous | X | X | ||
| Heliomaster constantii | Plain-capped Starthroat | Nectarivorous | X | X | ||
| Lampornis hemileucus | White-bellied Mountain-gem | Nectarivorous | X | |||
| Lampornis calolaemus | Purple-throated Mountain-gem | Nectarivorous | X | X | ||
| Calliphlox bryantae | Magenta-throated Woodstar | Nectarivorous | X | |||
| Selasphorus flammula | Volcano Hummingbird | Nectarivorous | X | |||
| Selasphorus scintilla | Scintillant Hummingbird | Nectarivorous | X | X | ||
| Chlorostilbon canivetii | Canivet’s Emerald | Nectarivorous | X | X | ||
| Klais guimeti | Violet-headed Hummingbird | Nectarivorous | X | X | ||
| Phaeochroa cuvierii | Scaly-breasted Hummingbird | Nectarivorous | X | |||
| Campylopterus hemileucurus | Violet Sabrewing | Nectarivorous | X | X | ||
| Eupherusa eximia | Stripe-tailed Hummingbird | Nectarivorous | X | X | ||
| Elvira cupreiceps | Coppery-headed Emerald | Nectarivorous | X | |||
| Chalyburaurochrysia | Bronze-tailed Plumeleteer | Nectarivorous | X | |||
| Thalurania colombica | Crowned Woodnymph | Nectarivorous | X | X | ||
| Amazilia amabilis | Blue-chested Hummingbird | Nectarivorous | X | |||
| Amazilia saucerrottei | Steely-vented Hummingbird | Nectarivorous | X | X | ||
| Amazilia tzacatl | Rufous-tailed Hummingbird | Nectarivorous | X | X | X | |
| Amazilia rutila | Cinnamon Hummingbird | Nectarivorous | X | X | ||
| Hylocharis eliciae | Blue-throated Goldentail | Nectarivorous | X | |||
| TROGONIFORMES | ||||||
| Trogonidae | ||||||
| Trogon clathratus | Lattice-tailed Trogon | Medium frugivouros | 30 | X | X | |
| Trogon massena | Slaty-tailed Trogon | Medium frugivouros | 30 | X | X | |
| Trogon caligatus | Gartered Trogon | Medium frugivouros | 23 | X | X | |
| Trogon rufus | Black-throated Trogon | Medium frugivouros | 23 | X | ||
| Trogon collaris | Collared Trogon | Medium frugivouros | 25 | X | ||
| Trogon aurantiiventris | Orange-bellied Trogon | Medium frugivouros | 25 | X | ||
| Pharomachrus mocinno | Resplendent Quetzal | Large frugivouros | 36 | X | X | |
| PICIFORMES | ||||||
| Capitonidae | ||||||
| Eubucco bourcierii | Red-headed Barbet | Small frugivorous | 15 | X | ||
| Semnornithidae | ||||||
| Semnornis frantzii | Prong-billed Barbet | Small frugivorous | 17 | X | ||
| Ramphastidae | ||||||
| Aulacorhynchus prasinus | Emerald Toucanet | Medium frugivouros | 29 | X | X | |
| Pteroglossus torquatus | Collared Aracari | Large frugivouros | 41 | X | X | |
| Selenidera spectabilis | Yellow-eared Toucanet | Large frugivouros | 36 | X | ||
| Ramphastos sulfuratus | Keel-billed Toucan | Large frugivouros | 47 | X | X | X |
| Ramphastos ambiguus | Yellow-throated Toucan | Large frugivouros | 56 | X | ||
| PSITTACIFORMES | ||||||
| Psittacidae | ||||||
| Eupsittula nana | Olive-throated Parakeet | Medium frugivouros | 23 | X | ||
| Eupsittula canicularis | Orange-fronted Parakeet | Medium frugivouros | 22.5 | X | ||
| Ara ambiguus | Great Green Macaw | Large frugivouros | 79 | X | ||
| Psittacara finschi | Crimson-fronted Parakeet | Medium frugivouros | 28 | X | X | X |
| Bolborhynchus lineola | Barred Parakeet | Small frugivorous | 16 | X | ||
| Brotogeris jugularis | Orange-chinned Parakeet | Small frugivorous | 18 | X | X | X |
| Touit costaricensis | Red-fronted Parrotlet | Small frugivorous | 17 | X | ||
| Pyrilia haematotis | Brown-hooded Parrot | Medium frugivouros | 21 | X | X | |
| Pionus senilis | White-crowned Parrot | Medium frugivouros | 24 | X | X | X |
| Amazona albifrons | White-fronted Parrot | Medium frugivouros | 25 | X | ||
| Amazona autumnalis | Red-lored Parrot | Large frugivouros | 34 | X | ||
| Amazona farinosa | Mealy Parrot | Large frugivouros | 38 | X | ||
| PASSERIFORMES | ||||||
| Tyrannidae | ||||||
| Elaenia flavogaster | Yellow-bellied Elaenia | Small frugivorous | 15 | X | X | X |
| Elaenia frantzii | Mountain Elaenia | Small frugivorous | 15 | X | X | |
| Mionectes olivaceus | Olive-striped Flycatcher | Small frugivorous | 13 | X | ||
| Mionectes oleagineus | Ochre-bellied Flycatcher | Small frugivorous | 12.5 | X | X | X |
| Zimmerius vilissimus | Paltry Tyrannulet | Small frugivorous | 9.5 | X | X | |
| Rhynchocyclus brevirostris | Eye-ringed Flatbill | Small frugivorous | 15 | X | ||
| Tityridae | ||||||
| Tityra semifasciata | Masked Tityra | Medium frugivouros | 21 | X | X | X |
| Tityra inquisitor | Black-crowned Tityra | Small frugivorous | 18.5 | X | ||
| Pachyramphus versicolor | Barred Becard | Small frugivorous | 12 | X | ||
| Pachyramphus cinnamomeus | Cinnamon Becard | Small frugivorous | 14 | X | X | |
| Pachyramphus polychopterus | White-winged Becard | Small frugivorous | 14.5 | X | ||
| Pachyramphus aglaiae | Rose-throated Becard | Small frugivorous | 16.5 | X | ||
| Cotingidae | ||||||
| Querula purpurata | Purple-throated Fruitcrow | Medium frugivouros | 28 | X | ||
| Cephalopterus glabricollis | Bare-necked Umbrellabird | Large frugivouros | 41 | X | X | |
| Lipaugus unirufus | Rufous Piha | Medium frugivouros | 23 | X | ||
| Procnias tricarunculatus | Three-wattled Bellbird | Medium frugivouros | 30 | X | ||
| Carpodectes nitidus | Snowy Cotinga | Medium frugivouros | 22.5 | X | ||
| Pipridae | ||||||
| Corapipo altera | White-ruffed Manakin | Small frugivorous | 10 | X | ||
| Chiroxiphia linearis | Long-tailed Manakin | Small frugivorous | 11.5 | X | X | |
| Dixiphia pipra | White-crowned Manakin | Small frugivorous | 10 | X | ||
| Ceratopipra mentalis | Red-capped Manakin | Small frugivorous | 10 | X | ||
| Manacus candei | White-collared Manakin | Small frugivorous | 11 | X | ||
| Oxyruncidae | ||||||
| Oxyruncus cristatus | Sharpbill | Small frugivorous | 16.5 | X | ||
| Vireonidae | ||||||
| Vireo leucophrys | Brown-capped Vireo | Small frugivorous | 12 | X | ||
| Vireo laniuspulchellus | Green Shrike-Vireo | Small frugivorous | 14 | X | ||
| Cyclarhis gujanensis | Rufous-browed Peppershrike | Small frugivorous | 14 | X | ||
| Corvidae | ||||||
| Cyanolyca cucullata | Azure-hooded Jay | Medium frugivouros | 29 | X | ||
| Calocitta formosa | White-throated Magpie-Jay | Large frugivouros | 46 | X | ||
| Psilorhinus morio | Brown Jay | Large frugivouros | 39 | X | X | X |
| Turdidae | ||||||
| Myadestes melanops | Black-faced Solitaire | Small frugivorous | 17 | X | X | |
| Catharus aurantiirostris | Orange-billed Nightingale-Thrush | Small frugivorous | 16 | X | X | |
| Catharus fuscater | Slaty-backed Nightingale-Thrush | Small frugivorous | 17 | X | ||
| Catharus frantzii | Ruddy-capped Nightingale-Thrush | Small frugivorous | 16 | X | ||
| Catharus mexicanus | Black-headed Nightingale-Thrush | Small frugivorous | 16 | X | X | |
| Turdus plebejus | Mountain Thrush | Medium frugivouros | 24 | X | X | |
| Turdus obsoletus | Pale-vented Thrush | Medium frugivouros | 23.5 | X | X | |
| Turdus grayi | Clay-colored Thrush | Medium frugivouros | 23.5 | X | X | X |
| Turdus assimilis | White-throated Thrush | Medium frugivouros | 22 | X | ||
| Ptiliogonatidae | ||||||
| Phainoptila melanoxantha | Black-and-yellow Silky-Flycatcher | Medium frugivouros | 21 | X | ||
| Ptiliogonys caudatus | Long-tailed Silky-Flycatcher | Medium frugivouros | 24 | X | ||
| Fringillidae | ||||||
| Euphonia luteicapilla | Yellow-crowned Euphonia | Small frugivorous | 9.5 | X | ||
| Euphonia hirundinacea | Yellow-throated Euphonia | Small frugivorous | 11 | X | X | |
| Euphonia elegantissima | Elegant Euphonia | Small frugivorous | 11 | X | ||
| Euphonia gouldi | Olive-backed Euphonia | Small frugivorous | 9.5 | X | ||
| Euphonia minuta | White-vented Euphonia | Small frugivorous | 9 | X | ||
| Euphonia anneae | Tawny-capped Euphonia | Small frugivorous | 11 | X | X | |
| Chlorophonia callophrys | Golden-browed Chlorophonia | small frugivorous | 13 | X | X | |
| Passerellidae | ||||||
| Chlorospingus flavopectus | Common Bush-Tanager | Small frugivorous | 13.5 | X | X | |
| Chlorospingus pileatus | Sooty-capped Bush-Tanager | Small frugivorous | 13.5 | X | X | |
| Chlorospingus canigularis | Ashy-throated Bush-Tanager | Small frugivorous | 13 | X | ||
| Icteridae | ||||||
| Amblycercus holosericeus | Yellow-billed Cacique | Medium frugivouros | 23 | X | X | X |
| Psarocolius wagleri | Chestnut-headed Oropendola | Large frugivouros | 35 | X | X | |
| Psarocolius montezuma | Montezuma Oropendola | Large frugivouros | 50 | X | X | X |
| Cacicus uropygialis | Scarlet-rumped Cacique | Medium frugivouros | 23 | X | X | |
| Icterus prosthemelas | Black-cowled Oriole | Small frugivorous | 19 | X | ||
| Molothrus oryzivorus | Giant Cowbird | Large frugivouros | 33 | X | ||
| Cardinalidae | ||||||
| Piranga flava | Hepatic Tanager | Small frugivorous | 18 | X | ||
| Piranga bidentata | Flame-colored Tanager | Small frugivorous | 18 | X | ||
| Habia rubica | Red-crowned Ant-Tanager | Small frugivorous | 18 | X | X | |
| Habia fuscicauda | Red-throated Ant-Tanager | Small frugivorous | 19 | X | ||
| Chlorothraupiscarmioli | Carmiol’s Tanager | Small frugivorous | 17 | X | X | |
| Caryothraustes poliogaster | Black-faced Grosbeak | Small frugivorous | 16.5 | X | ||
| Passerina caerulea | Blue Grosbeak | Small frugivorous | 16.5 | X | ||
| Thraupidae | ||||||
| Thraupis episcopus | Blue-gray Tanager | Small frugivorous | 15 | X | X | X |
| Thraupis palmarum | Palm Tanager | Small frugivorous | 16 | X | X | X |
| Tangara larvata | Golden-hooded Tanager | Small frugivorous | 13 | X | X | |
| Tangara guttata | Speckled Tanager | Small frugivorous | 13 | X | ||
| Tangara dowii | Spangle-cheeked Tanager | Small frugivorous | 13 | X | ||
| Tangara inornata | Plain-colored Tanager | Small frugivorous | 12 | X | ||
| Tangara gyrola | Bay-headed Tanager | Small frugivorous | 13.5 | X | ||
| Tangara florida | Emerald Tanager | Small frugivorous | 12 | X | ||
| Tangara icterocephala | Silver-throated Tanager | Small frugivorous | 13 | X | ||
| Diglossa plumbea | SlatyFlowerpiercer | Nectarivorous | X | X | ||
| Chrysothlypis chrysomelas | Black-and-yellow Tanager | Small frugivorous | 12 | X | X | |
| Chlorophanes spiza | Green Honeycreeper | Small frugivorous | 13 | X | X | |
| Tachyphonus luctuosus | White-shouldered Tanager | Small frugivorous | 14 | X | ||
| Tachyphonus delattrii | Tawny-crested Tanager | Small frugivorous | 14.5 | X | ||
| Tachyphonus rufus | White-lined Tanager | Small frugivorous | 17 | X | ||
| Lanio leucothorax | White-throated Shrike-Tanager | Medium frugivouros | 20 | X | X | |
| Ramphocelus sanguinolentus | Crimson-collared Tanager | Small frugivorous | 18.5 | X | X | |
| Ramphocelus passerinii | Passerini’s Tanager | Small frugivorous | 16 | X | X | |
| Cyanerpes lucidus | Shining Honeycreeper | Small frugivorous | 10 | X | ||
| Cyanerpes cyaneus | Red-legged Honeycreeper | Small frugivorous | 11.5 | X | X | X |
| Dacnis venusta | Scarlet-thighed Dacnis | Small frugivorous | 11.5 | X | ||
| Dacnis cayana | Blue Dacnis | Small frugivorous | 11.5 | X | ||
| Coereba flaveola | Bananaquit | Small frugivorous | 9 | X | X | |
| Saltator atriceps | Black-headed Saltator | Medium frugivouros | 24 | X | ||
| Saltator maximus | Buff-throated Saltator | Medium frugivouros | 20 | X | X | X |
| Saltator grossus | Slate-colored Grosbeak | Small frugivorous | 19 | X | ||
| Saltator coerulescens | Grayish Saltator | Medium frugivouros | 20 | X | X | X |
Category refers to the classification of each species according to guild and body size.
Size where obtained from Stiles and Skutch (1989).

