











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkThe widespread conversion of land uses have established patterns related to the modification of native vegetation to uses dominated by agricultural land and livestock production (Foley et al., 2005; Haslem & Bennett, 2008; Houghton, 2013). These changes, and consequently the effects of the fragmentation of native forests, are of interest for the conservation of biodiversity (Heikkinen, Luoto, Virkkala, & Rainio, 2004; Jeliazkov et al., 2016). The intensification of land use for the establishment of livestock or agricultural crops defines the characteristics of production at the local scale and the basic structure of plant or landscape typological formations (Foley et al., 2005; Velásquez-Valencia et al., 2005) The operation of these types of production systems as well as social and economic influences have created diverse landscapes, mosaics composed of varied elements of vegetation cover (forests, crops, pastures, barren fields) which are distributed across spaces in different ways.
These new structures of the landscape acts as environmental filters altering the distribution patterns of the components of the biological communities, and positively affect predation (Roldán & Simonetti, 2001; Renjifo, 2001; Olson, Dinerstein, Powell, & Wikramanayake, 2002), and parasitism (Laurance & Yensen, 1991; Murcia, 1995; Redford, 1992) and changes the spatial patterns of vegetation cover in landscapes (Lindenmayer & Fischer, 2007). In this sense, both habitat loss and fragmentation operate at different spatial, temporal, habitat and biological organization levels.
Some research has focused on analyzing the pattern of spatial configuration of remaining habitats and their influence on the ability of organisms to persist in these modified landscapes (Trzcinski, Fahrig, & Merriam, 1999; McGarigal & Cushman, 2002; Bennett, Radford, & Haslem, 2006) and the relationship to species richness and abundance, independently of the sensitivity of species to anthropogenic disturbance The size, shape and proximity of the patches and the capacity of the species to use the matrix (Antongiovanni & Metzger, 2005) have been identified as modelers of the distribution patterns of bird diversity (Fahrig, 2003).
The effects of habitat fragmentation on biodiversity (Laurance, 1994; Laurance et al., 2002; Laurance & Vasconcelos, 2009) and the physical degradation of soils caused by intensive farming (Chauvel, Grimaldi, & Tessier, 1991) have been described. Some alternatives have been proposed on how to manage deforestation (Laurance & Gascon, 1997), such as the use of collective practices of agroforestry, which causes less deterioration and loss of biodiversity and the values of agricultural landscapes for the conservation of nature (Haila, 2002; McGarigal & Cushman, 2002) and the increase in the permeability of the matrix (Antongiovanni & Metzger, 2005).
The relative effects of habitat composition and the configuration of avian biodiversity distribution may be dependent upon the context of the landscape being studied (Jokimäki & Huhta, 1996; Schmiegelow, Machtans, & Hannnon, 1997). Towards the interiors of these systems, abundance and richness of this group decrease in modified habitat gradients from secondary forest to pasture lands (Donald, Green, & Heath, 2001; Kaboli et al., 2001; Peterjohn & Sauer, 1999).
The Andean Amazon in Southeastern Colombia is one of the areas where the colonization front is the most active in the country, and is considered a hot spot of deforestation. In the last 50 years, more than 40 % of the forests of the Andean Amazon territory of the department of Caquetá have been lost to deforestation. This condition is the result of urban growth and the expansion of production systems that are not friendly to the environment, including illicit crop production, livestock production, the exploitation of timber resources and the extraction of mining and energy resources.
For this reason, it is important to determine the distribution patterns of native birds in the Andean Amazon region of Southeastern Colombia. As this region is subject to a deep and continuous transformation of the landscape, it is necessary to identify and explain the patterns that emerge in the bird community with the transformation of the landscape in mosaics with varied vegetation cover patches that are immersed in the matrix of agroforestry and silvopastoral systems. In this sense, the objective of this research was to evaluate the distribution of the richness, abundance and diversity of the bird community in the configuration and composition of the agroforestry and silvopastoral landscapes. To direct the analysis of the data, the following hypothesis is formulated there is a dependence of the richness and abundance of bird species to the mosaics with a greater number and diversity of patches type of closed vegetation cover in the production systems in the region of the Andean Amazon.
Materials and methods
Area of study: The study was carried out in the southeastern region of the Republic of Colombia, in the department of Caquetá (1°37′ N & 75°36′ W), in the municipalities of Florencia (1°36’ N & 75°36’ W), Morelia (1°29’ N & 75°43’ W), Belén de los Andaquíes (1°24’5 N & 75°52’ W) and San Vicente del Caguán (2°06’ N & 74°46’ W). The average annual temperature and humidity are 25.1 °C and 81 % respectively.
The nature of this region is denominated as dense, exuberant jungle of the interfluves of the Putumayo and Caquetá rivers (Hernández-Camacho, 1992). Nevertheless, this jungle region is affected by the regular deforestation and subsequent burning of its forests for purposes related to livestock, agricultural and illegal crop production. According to alerts from Colombia’s Instituto de Hidrología, Meteorología y Estudios Ambientales, IDEAM (2017), this region is home to two of Colombia’s early alert deforestation nuclei, and the deforestation dynamic in the region corresponds to 39.70 % of the nation’s total deforestation.
In the study area, three production systems have been typified: silvopastoral (SSP), agroforestry (SAF) and traditional or extensive livestock farming (SGT), which determine the economy of the region. In this sense, 25.70 % of the territory of the department is composed of grazing areas, with an annual production of 1 555 443 heads of cattle, and more than 1 500 hectares cultivated in Hevea rubber and Elaeis African palm.
The study was conducted on 39 mosaics that were established within the three production systems. Each mosaic consisted of a circular area with a radius of 0.5 km (78.5 ha.). The area of each mosaic allowed the inclusion of multiple types of vegetation cover in the production system landscapes as well as the replication of sampling (Haslem & Bennett, 2008). Given the situation of armed conflict in the study area, the decisions for the selection of mosaics for sampling were made with ease of access and security conditions of the territory taken into account. In total, nine mosaics were established in SSP, eleven in SAF and 19 in SGT.
The classification of the land coverings in the mosaics was made by describing the spatial configuration of the vegetation according to the intensity and heterogeneity of the mosaics by means of calculations with FRAG-STAT Version 3.3 and developed with ArcMap. Two categories of vegetation cover were established; the first corresponds to open vegetation cover, and the second category corresponds to closed vegetation cover.
The elaboration of the cartography and the interpretation of the elements of the landscape were carried out by means of satellite images, following the conceptual and methodological guidelines for the definition and limitation of ecosystems put forth by Etter (1998) and Armenteras, Gast, and Villareal (2003). The verification and adjustment of the map were carried out through fieldwork trips made during the period designated for the capture of primary information.
Table 1 shows the three landscape properties selected to quantify the mosaic variables: the spatial configuration (N = 5), the composition (N = 3) and the aggregation (N = 4) of the vegetation cover. The variables that describe the spatial configuration and the heterogeneity of the mosaics were calculated with FRAG-STAT Version 3.3. All spatial information was developed in ArcMap Version 10.
Table 1: Analysis of landscape properties and variables of the metrics of class and landscape mosaics
| Properties of the mosaic | Variable | Abbreviation | Description |
| Composition | Shannon diversity index | SHDI | Assesses landscape diversity, or heterogeneity, based on diversity of fragments |
| Richness of patch | PR | Bears the number of different types of land use/coverage that exist in the landscape | |
| Simpson diversity index | SIDI | Assesses, based on probabilities, landscape diversity or heterogeneity | |
| Number of patches | NP | Measures the fragmentation of landscape | |
| Aggregation | Division | DIVISION | Measures the probability that two cells chosen at random do not belong to the same region |
| Landscape form index | LSI | Calculates the relationship between area and perimeter for the combined landscape | |
| Distance of the nearest neighbor | ENN | Distance to the closest fragment of the same class | |
| Configuration | Mean perimeter area shape | PARA | Perimeter of the group of fragments |
| Mean shape index | SHAPE | Measures the complexity of the shape | |
| Area | AREA | Calculates the area that corresponds to each fragment | |
| Largest patch index | LPI | Quantifies the percentage of total area occupied by the largest region |
In total, twelve types of vegetation cover were determined in the mosaics, which were grouped into two categories. The first corresponds to the types of open vegetation cover, consisting of Lagoon (LGN), Pastures with Scattered Trees (PAD), Overgrown Pastures (PEN), Clean Pastures (PPL), Floodplain Pastures (PHO), and River (RIO). The second category corresponds to closed vegetation cover, consisting of Dense Forest (BDN), Fragmented Forest (BFG), Riparian Forest (BRP), Cultivation of Rubber Trees (CCH), Low Secondary Vegetation (RTT), and High Secondary Vegetation (RTV). The composition of these coverings in the mosaics, their configuration and their structural properties provide the habitat resources to birds present in the production systems of the Andean Amazon region.
In total, 65.4 % of the mosaic areas correspond to types of open vegetation coverage. The largest coverage area type was the Pastures with Scattered Trees, with 37.2 % of the total coverage area, which is double the surface size of Clean Pastures. These two types of coverage were found in 33 and 35 mosaics respectively. Dense forest and secondary vegetation presented the largest areas in closed vegetation types with 12.5 % and 8.5 % respectively of the total coverage area. Significant differences were found in the average coverage area (F = 17.28, g.l. = 252, P < 0.05) and the average of largest patch index (LPI) among the vegetation coverages in the mosaics (F = 18.35, g.l. = 252, P < 0.05).
Table 2 shows the analysis of the area of land cover in the production system landscapes. The coverage that presented the area of greatest occupation in the mosaics of silvopastoral production systems was PPL, as this type of coverage was present in all production systems. The CCH coverage was only present in SAF, and the RIO coverage present only in SGT. The average pasture area (PPL) showed significant differences between SSP and SGT (F = 3.62, g.l. = 44, P < 0.05). The average area of mosaic land cover within the production systems presented significant differences for SSP (F = 26.35, g.l. = 50, P < 0.05), SAF (F = 12.72, g.l. = 78, P < 0.05) and in SGT (F = 4.31, g.l. = 93, P < 0. 05).
Table 2: Analysis of the area of land cover in the production system landscapes in the Andean Amazon region in the Department of Caquetá
| Land Cover | SSP | SAF | SGT | |||||||||||
| Area | Area | Area | ||||||||||||
| Mean (ha)* | Max (ha) | Total (ha) | Mean (ha)* | Max (ha) | Total (ha) | Mean (ha)* | Max (ha) | Total (ha) | ||||||
| BDN | 5.38 | b | 26.84 | 43.00 | 9.83 | b | 21.04 | 78.63 | 13.90 | ab | 74.88 | 264.09 | ||
| BFG | 4.41 | b | 6.49 | 22.07 | 8.53 | b | 16.85 | 51.15 | 8.23 | abc | 12.85 | 32.94 | ||
| BRP | 1.54 | b | 5.66 | 20.05 | 1.93 | b | 13.20 | 36.59 | 1.74 | bc | 6.71 | 39.91 | ||
| CCH | - | - | - | 5.31 | b | 28.72 | 69.06 | - | - | - | ||||
| LAG | 0.42 | b | 2.51 | 13.35 | 0.74 | b | 2.76 | 4.46 | 0.33 | c | 1.27 | 3.29 | ||
| PAD | 10.38 | a | 62.04 | 436.11 | 7.14 | a | 53.07 | 342.87 | 13.99 | a | 72.77 | 419.73 | ||
| PEN | 7.04 | b | 23.23 | 28.15 | 2.72 | b | 5.94 | 13.61 | 4.11 | abc | 9.24 | 32.88 | ||
| PPL | 1.45 | b | 10.49 | 37.65 | 4.06 | b | 20.45 | 109.75 | 7.01 | abc | 61.95 | 434.90 | ||
| PHO | 1.03 | b | 4.15 | 15.42 | 1.64 | b | 6.54 | 24.53 | 3.24 | abc | 11.47 | 61.50 | ||
| RIO | - | - | - | - | - | - | 6.39 | abc | 15.02 | 44.72 | ||||
| RTV | 2.65 | b | 10.58 | 47.77 | 2.71 | b | 18.49 | 81.32 | 5.01 | abc | 31.92 | 125.28 | ||
| RTT | 2.38 | b | 10.94 | 47.68 | 1.74 | b | 9.10 | 57.41 | 1.45 | abc | 9.09 | 42.07 | ||
Bird census: Birds were surveyed by the point count method, in fixed stations within each mosaic. The points were established in a grid on the surface of the mosaic, which consisted of three parallel linear transects, each separated by 500 m. In each transect, three observation points were located 500 m apart from each other. Each census was carried out in a time of 15 min, on a radial surface of 50 m around each point.
All the species seen or heard were registered by a single observer (the first author of the article). The census was carried out in the years 2012 and 2013, with periodic interruptions due to the security conditions in the area. In total, 351 points (9 points in each of the 39 mosaics) were sampled in the whole area and 135 minutes of observation were made within each mosaic, for a total of 5 265 min of observation. The total species richness and the abundance of each species were estimated for each point and mosaic, and the association was carried out to the different vegetation coverings present in the mosaics.
Statistical analysis: In order to evaluate the effort and completeness of species richness based on the sampling effort, accumulation curves were plotted using Chao 2, Chao 1, Jackknife 1 and environmental Jackknife 2 estimators (Colwell & Coddington, 1994; Halffter & Moreno, 2005) calculated in the EstimateS program (Version 8.2). The rarefaction curve was used to compare the number of species when the samples differed in size (Gotelli & Colwell, 2001), which estimates the species richness as a function of the smallest sample size in the production systems.
In order to geographically represent the species richness observed in the sampling effort and the integrity of the inventory throughout territorial units (geospatial location of the mosaics), the KnobR (Biogeographical Knowledge in R) was performed, which calculates the integrity and a series of estimators of richness and species accumulation curves at a resolution of 15 geographical minutes, using the R-Wizard program.
The incidence of the production system and the area of the mosaic’s vegetation cover in the richness and abundance of species were tested through a Kruskal-Wallis analysis for each variable. The Jaccard index was used to calculate the similarity in species composition of bird communities between production systems. In order to describe the relationship of diversity indices of mosaic bird communities between production systems and mosaic landscape metrics, a principal component analysis (PCA) was performed, in which each index value was used as a descriptive variable.
Results
We registered 262 species of birds, 6 720 individuals, distributed across 23 orders and 54 families within the 39 production system mosaics. In total, 97 species of birds were observed only once during sampling. The average species richness and average abundance recorded per mosaic was 39 ± 19 species and 172.3 ± 101.2 individuals respectively.
The richness of species observed throughout territorial units (geospatial location of the mosaics) in the study area is between 71 and 87 % of completeness, in accordance with the estimated richness values determined through Jackknife and Chao estimators (Figure 1).
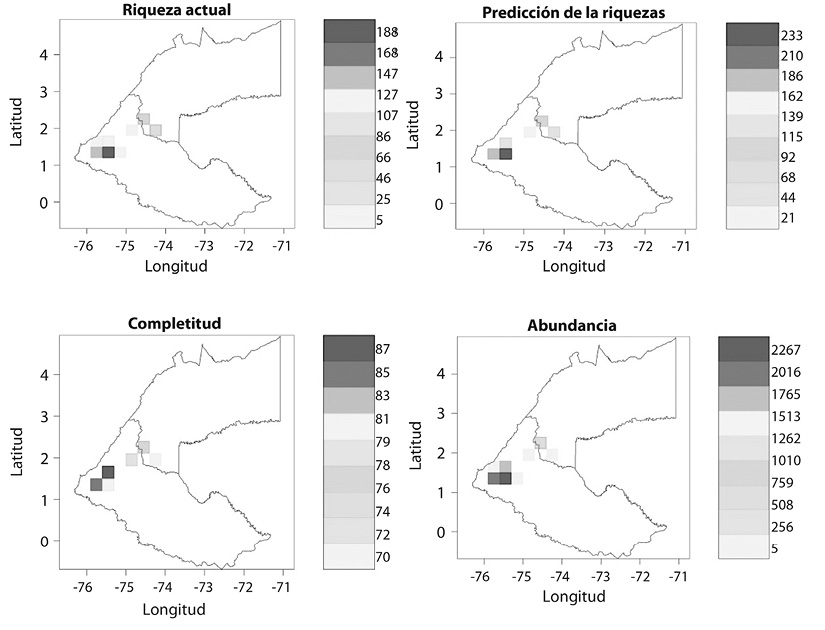
Figure 1 Analysis of the geographical distribution of the richness of birds (Biogeographical Knowledge) of the production systems in the Andean Amazon region in the Department of Caquetá in 15-minute spatial resolution, corresponding to cells of 650 km2.
The inventory of the region represented more than 80 % of the potential average species to be detected in the study area. Therefore, the integrity of the recorded species and the sampling effort allow us to understand the patterns and processes in the landscape mosaics of the production systems studied (Figure 2).
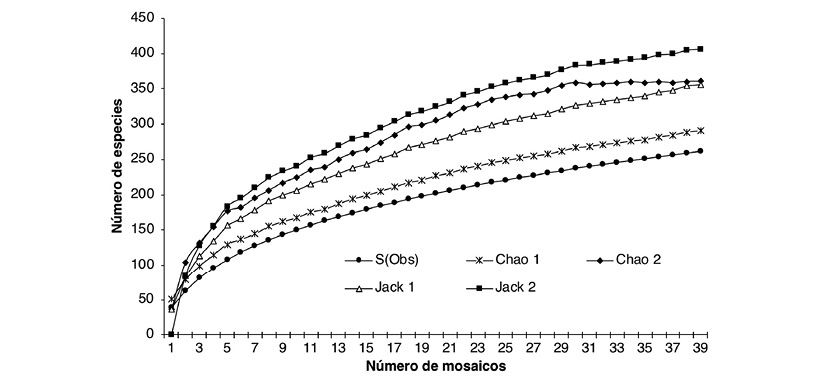
Figure 2 Species accumulation curve of the mosaics of the study area in the landscapes of the production systems in the Andean Amazon region in the Department of Caquetá.
Table 3 shows the estimated richness calculated for each production system. These estimators predicted a greater number of species than those observed. The system of traditional livestock production (SGT) was the system where the greatest difference between the values of observed and expected richness was presented. In total, the richness observed in all production systems represents between 64.3 and 90.3 % of the richness of the estimators. The SGT system presented the highest values of species richness (S = 193) and individuals (N = 3 238). However, the richness and average abundance of the mosaics between the systems did not present significant differences (K-W = 3.38, g.l. = 2, P > 0.05) and (K-W = 2.73, g.l. = 2, P > 0.05) respectively.
Table 3: Values of the richness and completeness of samples taken of birds in the mosaics of production systems of the Andean Amazon in the Department of Caquetá
| System | Number of Mosaics | Species observed | Estimator | % Completeness | |||
| Chao 1 | Chao 2 | Jackknife 1 | Jackknife 2 | ||||
| SGT | 19 | 193 | 223 | 283 | 267 | 310 | 62-86 |
| SAF | 11 | 186 | 207 | 279 | 259 | 302 | 62-90 |
| SSP | 9 | 101 | 113 | 133 | 136 | 152 | 66-89 |
| Total | 39 | 262 | 292 | 364 | 357 | 407 | 64-90 |
Two species, Bubulcus ibis and Ara severus, represent 20 % of the total number of individuals registered in the 39 mosaics; 62.9 % of the species identified had abundances of less than ten individuals, and more than half of these only registered one or two individuals. Even though we did not find significant differences in the average abundances in the mosaics between production systems, the distribution of species abundance among production systems varied considerably between the SSP system and the SAF and SGT systems.
The latter two systems had a more homogeneous distribution of abundance among their species. In general, the curve of abundance distribution in production systems of the Andean Amazon region presents few abundant species, some species with intermediate abundances and many rare species (Figure 3). In this sense, these curves follow the geometric distribution model (X2 = 5.76, P < 0.05).
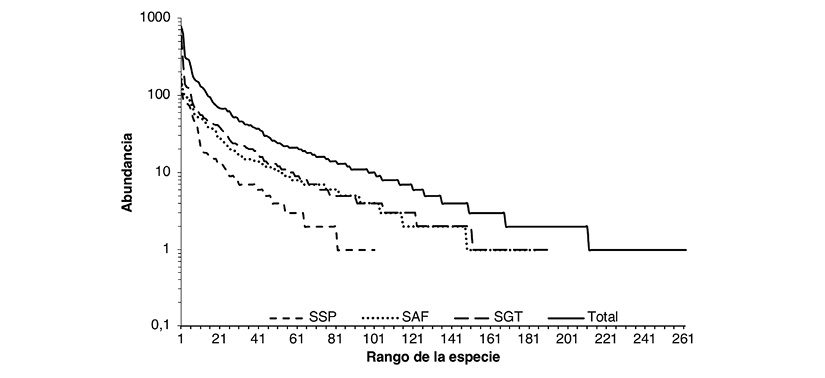
Figure 3 Distribution curve of the abundance of mosaics sampled in production systems in the Andean Amazon region in the Department of Caquetá.
The traditional livestock system registered 3 244 individuals, distributed in 22 orders, 51 families, 141 genera and 193 species; this system was dominated by the species: Bubulcus ibis (N = 592), Ara severus (N = 264), Thraupis palmarum (N = 140), Crotophaga ani (N = 126) and Tyrannus melancholicus (N = 124). It should be noted that 42 species presented only one individual. The families Ardeidae (N = 606, S = 6), Thraupidae (N = 424, S = 20) and Tyrannidae (N = 287, S = 28) were the families with the largest number of individuals, while families such as Anatidae, Caprimulgidae, Podicipedidae and Tytonidae only registered one species with one individual.
In the agroforestry system, 270 individuals corresponding to 21 orders, 45 families, 122 genera and 186 species were observed; only four species showed abundances greater than 90 individuals, A. severus (N = 186), T. melancholicus (N = 117), Thraupis episcopus (N = 97), and C. ani (N = 93). The Psitacidae Families (N = 365, S = 10), Tyrannidae (N = 326, S = 31) and Thraupidae (N = 308, S = 18) presented the greatest levels of richness and abundance; while the rest of the families obtained abundances between one and 15 individuals.
Finally, for the silvopastoral system, 1 206 individuals were found corresponding to 16 orders, 31 families, 80 genera and 101 species; only four species had the highest abundances, A. severus (N = 155), T. episcopus (N = 88), B. ibis (N = 86) and Amazona amazonica (N = 78); the remaining species showed very low abundances, among which 37 were represented by less than two individuals. The largest number of individuals were presented in the families: Psitacidae (N = 340, S = 10), Thraupidae (N = 199, S = 13) and Tyrannidae (N = 164, S = 18), unlike the Turdidae, Anatidae, Anhimidae, Alcedinidae and Therskiornithiadae families, which each had three or fewer individuals.
There is evidence of exclusivity and rarity of species among the systems. In SGT systems, 54 species (N = 150) presented exclusive registries and 134 were considered rare because they presented abundances of less than ten individuals. For the SAF systems, we found 52 exclusive species and 133 rare species, and in SSP systems we found 13 exclusive species. The SGT and SAF systems have 54 % similarity in the composition of their species, and these two compared with SSP have only 42 % similarity.
In general terms, the indices of richness, diversity and equity presented the highest values in the SAF system mosaics, while the indices of abundance and dominance were highest for the mosaics associated with the SSP and SGT systems.
The analysis of principal components (ACP) of the diversity indices, the first component (CP1) explains 63.89 % of the variance, and 24.90 % in the second component (CP2). The first axis groups the mosaics that presented the highest values of species dominance indices at the positive end, represented by Simpson and Berger-Parker, associated with SSP and SGT. At the negative end are the mosaics with the greatest Shannon diversity index, equitability (1-D) and richness (Margalef), associated with SAF systems. Similarly, the second component was associated with the mosaics that presented the highest number of individuals and species in the positive end; these mosaics belong to SGT and SAF systems. In the negative end were those with higher equity values (Equitability-J), which belong to SSP systems. In general, the two components are related to indices of richness, diversity, dominance and equity (Figure 4).
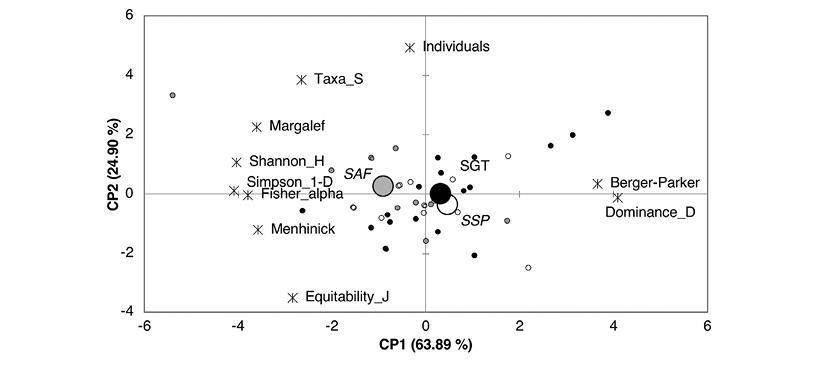
Figure 4 Analysis of the main components of diversity indices in production systems in the Andean Amazon region in the Department of Caquetá.
In the analysis of the main components of the diversity indices and the landscape metrics of the quadrants in the production systems, the first component contributes to the variance of 19.49 %. At its positive end, this component associates the indices of richness and diversity in landscapes with those within the bird community. At this extreme, biological diversity and landscape diversity present a positive association. The second component explains 17.90 % of the variance, and at its positive end it associates the variables of landscape metrics related to the shape and size of the patches with greater area, with the mosaics containing the highest species dominance values in the production system landscapes. At the negative end of this component, mosaics of greater richness and diversity of land patches are associated with the indices of richness and diversity of the bird assemblage (Figure 5).
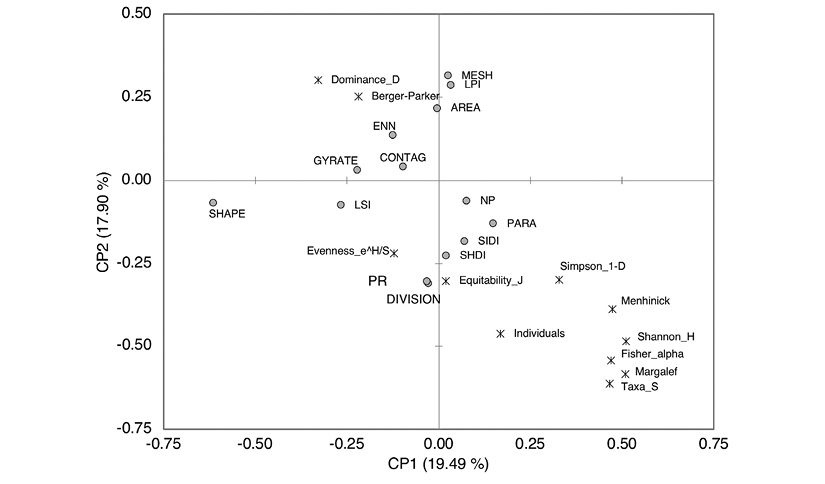
Figure 5 Analysis of the main components of the diversity indices and mosaic landscape metrics in the Agroforestry, Traditional Livestock and Silvopastoral production systems in the region of the Andean Amazon in the department of Caquetá.
In the analysis of canonical correlations, the first two components explain 45.98 % of the variability of the model. The first axis presents a positive correlation between the closed vegetation land uses and the dominance indices, and in the negative extreme, there is a correlation between open vegetation land uses and the indices of richness and diversity present in the mosaics. The second axis correlates the equity and richness indices with heterogeneous land uses in the positive end, and shows a decreasing complexity gradient for land uses with homogeneous coverage associated with dominance and abundance indices in the negative end (Figure 6).
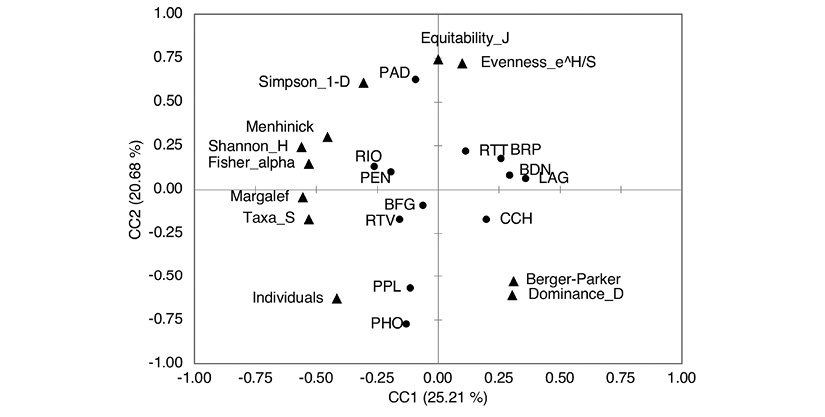
Discussion
In this research, 262 species of birds were observed across all landscape elements of the 39 mosaics of the agroforestry, silvopastoral and traditional livestock production systems present in the Andean Amazon region of Southeastern Colombia in the Department of Caquetá. The spatial distribution of the richness and composition of the species across the sampled mosaics allows us to gain better knowledge related to the species present in this region of the country. The species accumulation curves (effort of the collector), presented values close to the calculated estimates. In this sense, the methods used and the punctual efforts put forth have been efficient and effective in documenting the registries of bird species (Rosenstock & Van Riper, 2001; Stiles & Rosseli, 1998; Velasquez-Valencia, 2009).
The fundamental structure of the abundance of bird communities in the production systems studied pertain to disturbed environments (May, 1981). The distribution curves of abundance in these systems are adjusted to communities that have been subjected to forces that modify their homeostasis, or to stages of early succession. The dominance of some species is the response to the disturbance of habitats (Cháux & Velásquez, 2009; Pearson, Gray, & Johannessen, 1983), which modify the types of coverage in the elements of the landscape (Velasquez-Valencia, 2009). In this sense, the distribution of the abundance of the communities present in the SAF, SSP and SGT production systems indicate a detriment to the habitat, characteristic of productive systems with dominant anthropogenic matrices.
In the mosaics studied, fragments, patches of forest and stubble are present. Inside these mosaics, birds vary in a predictable way as a function of the heterogeneity of the vegetation covers which define the characteristic of the production, composition and basic configuration of the typological formations of vegetation or landscape (Velásquez-Valencia et al., 2005; Lindenmayer et al., 2002). This is evident in the differences found in the diversity and composition of bird species observed in the mosaics, where there is a decrease in diversity values from closed, complex and heterogeneous vegetation land use associated with the systems of agroforestry production to the open and homogeneous vegetation land use associated with traditional livestock production.
Although no differences were found in the average distribution of richness and abundance in the production systems, the composition of the species between the systems does not present a high level of similarity. Likewise, Cruz-Trujillo (2014), did not find differences in the richness of birds captured between the patches of livestock and agroforestry systems of the Amazonian foothills, nor in the vegetation coverings found within these systems.
The diversity of the assemblage of birds showed a strong relation to the conditions of heterogeneity of the mosaic in the context of the landscape. The presence of small remnants of habitats (riparian forests, dense forests, pastures with trees) assumes a complementarity or supplementary in the landscape, in the sense that birds access available resources in adjacent patches or areas in order to meet their requirements (Renjifo, 2001).
The richness of species was sensitive to changes in the composition and heterogeneity of the landscape. The variability and the distance between the types of habitat patches can affect the permeability of the landscape (Cerezo, Conde, & Poggio, 2011; Pearson, 1993; Saab, 1999; Sisk, Haddad, & Ehrlich, 1997; Trzcinski et al., 1999), which influences the dispersion and presence of bird species (McIntyre, 1995; Wiens, Stenseth, Horne, & Ims, 1993; Wilson et al., 2017). Mosaics with less contrast favored the dominance, abundance and distribution of some species, particularly those associated with open vegetation cover. In this sense, habitat patches are a limitation for the presence of species from open areas (Guadagnin & Maltchik, 2007; Herkert, 1994; Robbins, Dawson, & Dowell, 1989; Sampson, 1980).
The areas and the arrangement of vegetation cover types in the production system mosaics reflect the systematic management of these landscapes; this has repercussions on the implications for the practice of landscape conservation and restoration. The different types of vegetation cover serve as points of contact in the dispersion and distribution of the species in these areas.
Although the greatest value of richness, in terms of number of species, is found in the traditional livestock system, we must consider the ecological characteristics of the species that are concentrated here. Since the land uses associated with this system are made up of open vegetation in early stages of succession such as early stubble, clean pastures and pastures with abundant trees, a large range of food resources are generated, which are exploited by opportunistic species of birds that exploit more than one resource or specialists from grassland areas (Herkert, 1994; Johnson & Igl, 2001).
Nevertheless, the maximum value of species diversity (H) presented itself in agroforestry systems. These results suggest that it is possible to generalize the effect of improvement on the complexity of habitats in bird communities. It is probable that the response of the communities depends upon the characteristics of each system, such as management practices and land use, and related factors such as the size of the remaining patches of native vegetation and corridors of riparian vegetation (Velasquez-Valencia, 2009). Therefore, and considering that landscape mosaics in agroforestry production systems are mostly associated with closed vegetation (e.g., RTV, BFG), the presence and mobility of birds that frequent adjacent areas, such as Dendrocolaptidos and Picidos, are favored.
The management and protection that may be given to vegetation cover such as old stubble, BFG, or BRP should be the priority for conservation and the first starting point for the restoration and connectivity of landscapes in the Andean Amazon region of Colombia. Another approach is the design of a landscape for an entire community of species with respect to a certain quantity of habitat in the landscape and the isolation of patches (Westphal, Field, & Possingham, 2007). However, economic, environmental and social conditions undermine the perpetuity of these patches that give heterogeneity to the mosaics of these production landscapes. The presence of pastures with scattered trees is clearly an alternative management that largely guarantees the diversity and richness of bird species in these systems. However, the benefits will be greater upon obtaining a complete understanding of the functions and roles that landscape elements play in the conservation of bird diversity and in ecosystem services for the producer.
Mosaics with greater number and diversity of patch types of closed, symmetric vegetation coverage presented greater diversity and richness. Therefore, we prove the hypothesis of the dependence of the richness and abundance of bird species on the heterogeneity of the mosaic. These closed vegetation coverings were associated with the mosaics of agroforestry systems. The dominance of species in the production system mosaics presented a positive relationship to the size of the patches. Mosaics with patches of open vegetation cover of greater size presented a greater dominance of species. These types of coverage were associated with the traditional livestock system.
The size of the mosaics studied allows us to estimate the response of richness, diversity and composition of bird communities to landscape conditions at this scale. Birds respond to the variation of the vegetation coverage of the production system mosaics in this extension. However, it is likely that at larger scales, some other patterns may be estimated in relation to the class and landscape variables selected in this investigation. This is consistent with what was stated by Smith, Fahrig, and Francis (2011) which establishes the ability to predict the most important landscape variables for all response variables in landscapes of 500 m in radius.
Ethical statement: authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. A signed document has been filed in the journal archives

